
Proteome Analysis of Male Accessory Gland Secretions in the Diamondback Moth, Plutella xylostella (Lepidoptera: Plutellidae)
 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Tissue Collection
2.3. Protein Extraction and Digestion
2.4. Quantitative Proteomic Analysis of MAGs Based on Tandem Mass Tags (TMT)
2.5. Qualitative Proteomic Analysis of Mated-CB Based on Shotgun LC-MS/MS
2.6. Data Analysis
2.7. Identification of ACPs of P. xylostella
2.8. Annotation of ACPs and Comparison with Other Insects
2.9. Phylogenetic Analysis
2.10. Tissue-Specific Expression Analysis by qRT-PCR
3. Results
3.1. Identification of DAPs between Mated and Virgin MAG
3.2. Identification of Proteins in Mated-CB
3.3. Secreted ACPs Identification in the MAGs
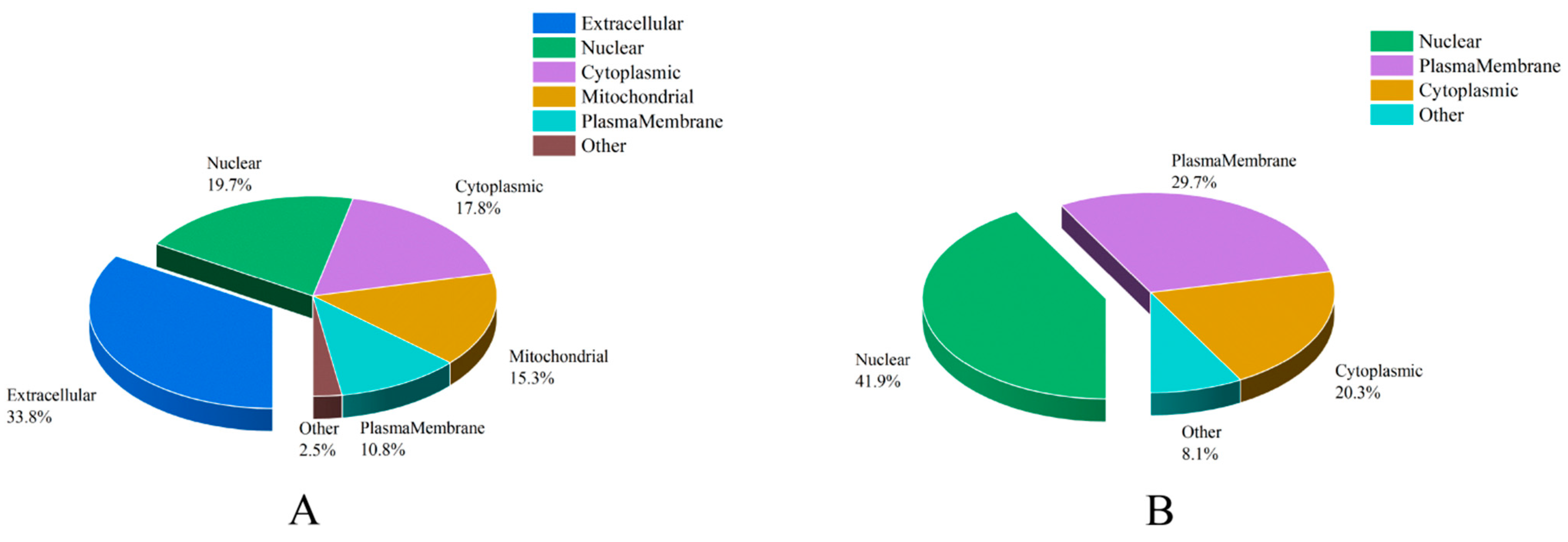
3.4. Subcellular Localization Analysis of ACPs
3.5. Functional Classification Analysis of ACPs
3.6. Comparison of the ACPs of P. xylostella with Other Insects
3.7. Tissue-Specific Expression Analysis by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SFPs | seminal fluid proteins |
| ACPs | accessory gland proteins |
| MAG | male accessory gland |
| TE | testis; |
| CB | copulatory bursa |
| VD | vas deferen |
| DAPs | differentially abundant proteins |
| qRT-PCR | reverse-transcription quantitative PCR |
| TMT | Tandem Mass Tag |
References
- Loher, W.; Ganjian, I.; Kubo, I.; Stanley-Samuelson, D.; Tobe, S.S. Prostaglandins: Their role in egg-laying of the cricket Teleogryllus commodus. Proc. Natl. Acad. Sci. USA 1981, 78, 7835–7838. [Google Scholar] [CrossRef]
- Pondeville, E.; Maria, A.; Jacques, J.C.; Bourgouin, C.; Dauphin-Villemant, C. Anopheles gambiae males produce and transfer the vitellogenic steroid hormone 20-hydroxyecdysone to females during mating. Proc. Natl. Acad. Sci. USA 2008, 105, 19631–19636. [Google Scholar] [CrossRef]
- Avila, F.W.; Sirot, L.K.; LaFlamme, B.A.; Rubinstein, C.D.; Wolfner, M.F. Insect seminal fluid proteins: Identification and function. Annu. Rev. Entomol. 2011, 56, 21–40. [Google Scholar] [CrossRef]
- Neubaum, D.M.; Wolfner, M.F. Mated Drosophila melanogaster females require a seminal fluid protein, Acp36DE, to store sperm efficiently. Genetics 1999, 153, 845–857. [Google Scholar] [CrossRef]
- Yu, J.F.; Li, C.; Xu, J.; Liu, J.H.; Ye, H. Male accessory gland secretions modulate female post-mating behavior in the moth Spodoptera litura. J. Insect Behav. 2014, 27, 105–116. [Google Scholar] [CrossRef]
- Lung, O.; Wolfner, M.F. Identification and characterization of the major Drosophila melanogaster mating plug protein. Insect Biochem. Mol. Biol. 2001, 31, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.S.; Stumm-Zollinger, E.; Aigaki, T.; Balmer, J.; Bienz, M.; Bohlen, P. A male accessory gland peptide that regulates reproductive behavior of female D. melanogaster. Cell 1988, 54, 291–298. [Google Scholar] [CrossRef]
- Moshitzky, P.; Fleischmann, I.; Chaimov, N.; Saudan, P.; Klauser, S.; Kubli, E.; Applebaum, S.W. Sex-peptide activates juvenile hormone biosynthesis in the Drosophila melanogaster corpus allatum. Arch. Insect Biochem. Physiol. 1996, 32, 363–374. [Google Scholar] [CrossRef]
- Chapman, T.; Bangham, J.; Vinti, G.; Seifried, B.; Lung, O.; Wolfner, M.F.; Smith, H.K.; Partridge, L. The sex peptide of Drosophila melanogaster: Female post-mating responses analyzed by using RNA interference. Proc. Natl. Acad. Sci. USA 2003, 100, 9923–9928. [Google Scholar] [CrossRef] [PubMed]
- Kingan, T.G.; Thomas-Laemont, P.A.; Raina, A.K. Male accessory gland factors rlicit change from ‘virgin’ to ‘mated’ behaviour in the female corn earworm Helicoverpa sea. J. Exp. Biol. 1993, 183, 61–76. [Google Scholar] [CrossRef]
- Ramm, S.A.; McDonald, L.; Hurst, J.L.; Beynon, R.J.; Stockley, P. Comparative proteomics reveals evidence for evolutionary diversification of rodent seminal fluid and its functional sig-nificance in sperm competition. Mol. Biol. Evol. 2009, 26, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Sepil, I.; Hopkins, B.R.; Dean, R.; Thezenas, M.L.; Charles, P.D.; Konietzny, R.; Fischer, R.; Kessler, B.M.; Wigby, S. Quantitative proteomics identification of seminal fluid proteins in male Drosophila melanogaster. Mol. Cell. Proteom. 2019, 18, S46–S58. [Google Scholar] [CrossRef] [PubMed]
- Baer, B.; Heazlewood, J.L.; Taylor, N.L.; Eubel, H.; Millar, A.H. The seminal fluid proteome of the honeybee Apis mellifera. Proteomics 2009, 9, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Sirot, L.K.; Hardstone, M.C.; Helinski, M.E.; Ribeiro, J.M.; Kimura, M.; Deewatthanawong, P.; Wolfner, M.F.; Harrington, L.C. Towards a semen proteome of the dengue vector mosquito: Protein identification and potential functions. PLoS Negl. Trop. Dis. 2011, 5, e989. [Google Scholar] [CrossRef]
- Simmons, L.W.; Tan, Y.F.; Millar, A.H. Sperm and seminal fluid proteomes of the field cricket Teleogryllus oceanicus: Identification of novel proteins transferred to females at mating. Insect Mol. Biol. 2013, 22, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Walters, J.R.; Harrison, R.G. Combined EST and proteomic analysis identifies rapidly evolving seminal fluid proteins in Heliconius butterflies. Mol. Biol. Evol. 2010, 27, 2000–2013. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Baulding, J.; Palli, S.R. Proteomics of Tribolium castaneum seminal fluid proteins: Identification of an angiotensin-converting enzyme as a key player in regulation of reproduction. J. Proteom. 2013, 78, 83–93. [Google Scholar] [CrossRef]
- Wei, D.; Li, H.M.; Tian, C.B.; Smagghe, G.; Jia, F.X.; Jiang, H.B.; Dou, W.; Wang, J.J. Proteome analysis of male accessory gland secretions in oriental fruit flies reveals juvenile hormone-binding protein, suggesting impact on female reproduction. Sci. Rep. 2015, 5, 16845. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Li, D.T.; Lu, J.B.; Zhang, W.X.; Zhang, C.X. Seminal fluid protein genes of the brown planthopper, Nilaparvata lugens. BMC Genom. 2016, 17, 654. [Google Scholar] [CrossRef] [PubMed]
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Feng, X.; Liu, S.S.; You, M.; Furlong, M.J. Biology, Ecology, and Management of the Diamondback Moth in China. Annu. Rev. Entomol. 2016, 61, 277–296. [Google Scholar] [CrossRef]
- You, M.; Ke, F.; You, S.; Wu, Z.; Liu, Q.; He, W.; Baxter, S.W.; Yuchi, Z.; Vasseur, L.; Gurr, G.M.; et al. Variation among 532 genomes unveils the origin and evolutionary history of a global insect herbivore. Nat. Commun. 2020, 11, 2321. [Google Scholar] [CrossRef]
- Myron, P.Z.; Asad, S.; Rehan, S.; David, A.; Liu, S.; Michael, J.F. Estimating the economic cost of one of the world’s major insect pests, Plutella xylostella (Lepidoptera: Plutellidae): Just How Long is a Piece of String? J. Econ. Entomol. 2012, 105, 1115–1129. [Google Scholar]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Oda, Y.; Uryu, M.; Hayakawa, Y. Insect cytokine growth-blocking peptide triggers a termination system of cellular immunity by inducing its binding protein. J. Biol. Chem. 2003, 278, 38579–38585. [Google Scholar] [CrossRef]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Liu, M.; Pi, H.; Xi, Y.; Wang, L.; Tian, L.; Chen, M.; Xie, J.; Deng, P.; Zhang, T.; Zhou, C.; et al. KIF5A-dependent axonal transport deficiency disrupts autophagic flux in trimethyltin chlo-ride-induced neurotoxicity. Autophagy 2021, 17, 903–924. [Google Scholar] [CrossRef]
- Wu, C.X.; Liu, Z.F. Proteomic profiling of sweat exosome suggests its involvement in skin immunity. J. Investig. Dermatol. 2018, 138, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, Y.; Xu, Q.; Zhu, J.; Lin, Y. TMT-based quantitative proteomics analysis reveals the key proteins related with the differentiation process of goat intramuscular adipocytes. BMC Genom. 2021, 22, 417. [Google Scholar] [CrossRef]
- Huang, W.; Huang, P.; Yü, D.; Li, C.; Huang, S.; Qi, P.; Huang, S.; Keyhani, N.O.; Huang, Z. Proteomic analysis of a hypervirulent mutant of the insect-pathogenic fungus Metarhizium anisopliae reveals changes in pathogenicity and terpenoid pathways. Microbiol. Spectr. 2022, 10, e0076022. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, Q.; Barbero, M.; Liu, L.; Nguyen, T.; Xu, A.; Ji, L. Proteins and signaling pathways response to dry needling combined with static stretching treatment for chronic myofascial pain in a RAT model: An explorative proteomic study. Int. J. Mol. Sci. 2019, 20, 564. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Chen, H.; Liu, Y.; Liu, Y. Proteomic analysis of kiwifruit in response to the postharvest pathogen, Botrytis cinerea. Front. Plant Sci. 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Thailayil, J.; Magnusson, K.; Godfray, H.C.; Crisanti, A.; Catteruccia, F. Spermless males elicit large-scale female responses to mating in the malaria mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2011, 108, 13677–13681. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.P.; Fang, Y.L.; Zhang, Z.N. Effect of male and female multiple mating on the fecundity, fertility, and longevity of diamondback moth, Plutella xylostella (L.). J. Appl. Entomol. 2005, 129, 39–42. [Google Scholar] [CrossRef]
- Pilch, B.; Mann, M. Large-scale and high-confidence proteomic analysis of human seminal plasma. Genome Biol. 2006, 7, R40. [Google Scholar] [CrossRef]
- Degner, E.C.; Ahmed-Braimah, Y.H.; Borziak, K.; Wolfner, M.F.; Harrington, L.C.; Dorus, S. Proteins, transcripts, and genetic architecture of seminal fluid and sperm in the mosquito Aedes aegypti. Mol. Cell. Proteom. 2019, 18, S6–S22. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Heifetz, Y.; Vandenberg, L.N.; Cohn, H.I.; Wolfner, M.F. Two cleavage products of the Drosophila accessory gland protein ovulin can independently induce ovulation. Proc. Natl. Acad. Sci. USA 2005, 102, 743–748. [Google Scholar] [CrossRef]
- Findlay, G.D.; Yi, X.; Maccoss, M.J.; Swanson, W.J. Proteomics reveals novel Drosophila seminal fluid proteins transferred at mating. PLoS Biol. 2008, 6, e178. [Google Scholar] [CrossRef]
- Braswell, W.E.; Andres, J.A.; Maroja, L.S.; Harrison, R.G.; Howard, D.J.; Swanson, W.J. Identification and comparative analysis of accessory gland proteins in Orthoptera. Genome 2006, 49, 1069–1080. [Google Scholar] [CrossRef]
- Mueller, J.L.; Ripoll, D.R.; Aquadro, C.F.; Wolfner, M.F. Comparative structural modeling and inference of conserved protein classes in Drosophila seminal fluid. Proc. Natl. Acad. Sci. USA 2004, 101, 13542–13547. [Google Scholar] [CrossRef] [PubMed]
- Laflamme, B.A.; Wolfner, M.F. Identification and function of proteolysis regulators in seminal fluid. Mol. Reprod. Dev. 2013, 80, 80–101. [Google Scholar] [CrossRef] [PubMed]
- Sirot, L.K.; Poulson, R.L.; McKenna, M.C.; Girnary, H.; Wolfner, M.F.; Harrington, L.C. Identity and transfer of male reproductive gland proteins of the dengue vector mosquito, Aedes aegypti: Potential tools for control of female feeding and reproduction. Insect Biochem. Mol. Biol. 2008, 38, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Dorus, S.; Wasbrough, E.R.; Busby, J.; Wilkin, E.C.; Karr, T.L. Sperm proteomics reveals intensified selection on mouse sperm membrane and acrosome genes. Mol. Biol. Evol. 2010, 27, 1235–1246. [Google Scholar] [CrossRef]
- Ravi, R.K.; Sirot, L.K.; Wolfner, M.F. Predicted seminal astacin-like protease is required for processing of reproductive proteins in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2006, 103, 18674–18679. [Google Scholar] [CrossRef]
- Marshall, J.L.; Huestis, D.L.; Hiromasa, Y.; Wheeler, S.; Oppert, C.; Marshall, S.A.; Tomich, J.M.; Oppert, B. Identification, RNAi knockdown, and functional analysis of an ejaculate protein that mediates a postmating, prezygotic phenotype in a cricket. PLoS ONE 2009, 4, e7537. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Y.; Bi, H.; Xu, J.; Liu, Z.; Niu, C.; He, L.; James, A.A.; Li, K.; Huang, Y. Mutation of the seminal protease gene, serine protease 2, results in male sterility in diverse lepidopterans. Insect Biochem. Mol. Biol. 2020, 116, 103243. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Y.; Li, X.; Li, R.; Xu, Y.; Shi, L.; Wang, H. Apolipoprotein D in Lepidoptera: Evolution and functional divergence. Biochem. Biophys. Res. Commun. 2020, 526, 472–478. [Google Scholar] [CrossRef]
- Izadi, H.; Subrahmanyam, B. Effect of JH III and substituted oxime ethers on the in vitro protein and RNA synthesis in male accessory reproductive gland (MARG) of Spodoptera litura (F.). Commun. Agric. Appl. Biol. Sci. 2005, 70, 849–856. [Google Scholar]
- Tian, L.; Ji, B.Z.; Liu, S.W.; Jin, F.; Gao, J.; Li, S. Juvenile hormone III produced in male accessory glands of the longhorned beetle, Apriona germari, is transferred to female ovaries during copulation. Arch. Insect Biochem. Physiol. 2010, 75, 57–67. [Google Scholar] [CrossRef]
- Huang, M.; Lin, H.; Xu, C.; Yu, Q.; Wang, X.; Qin, J.G.; Chen, L.; Han, F.; Li, E. Growth, metabolite, antioxidative capacity, transcriptome, and the metabolome response to dietary choline chloride in pacific white shrimp Litopenaeus vannamei. Animals 2020, 10, 2246. [Google Scholar] [CrossRef] [PubMed]
- Morais, C.J.; Atkins, G.L.; Sanchez, L.M.; Lopez-Boado, Y.S.; Lopez-Otin, C.; Sawyer, L. Arachidonic acid binds to apolipoprotein D: Implications for the protein’s function. FEBS Lett. 1995, 366, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Wang, X.; Shang, L.; Huang, Y.; Li, T.; Wu, C.; Zhang, R.; Zhang, J. Involvement of a versatile pattern recognition receptor, apolipophorin-III in prophenoloxidase activation and antibacterial defense of the Chinese oak silkworm, Antheraea pernyi. Dev. Comp. Immunol. 2016, 65, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef]
- Zeng, Y.; Yang, Y.T.; Wu, Q.J.; Wang, S.L.; Xie, W.; Zhang, Y.J. Genome-wide analysis of odorant-binding proteins and chemosensory proteins in the sweet potato whitefly, Bemisia tabaci. Insect Sci. 2019, 26, 620–634. [Google Scholar] [CrossRef]
- Chintapalli, V.R.; Wang, J.; Dow, J.A. Using FlyAtlas to identify better Drosophila melanogaster models of human disease. Nat. Genet. 2007, 39, 715–720. [Google Scholar] [CrossRef]
- Dong, S.; Ye, Z.; Tikhe, C.V.; Tu, Z.J.; Zwiebel, L.J.; Dimopoulos, G. Pleiotropic odorant-binding proteins promote Aedes aegypti reproduction and flavivirus transmission. mBio 2021, 12, e253121. [Google Scholar] [CrossRef]
- McGraw, L.A.; Gibson, G.; Clark, A.G.; Wolfner, M.F. Genes regulated by mating, sperm, or seminal proteins in mated female Drosophila melanogaster. Curr. Biol. 2004, 14, 1509–1514. [Google Scholar] [CrossRef]
- Lung, O.; Kuo, L.; Wolfner, M.F. Drosophila males transfer antibacterial proteins from their accessory gland and ejaculatory duct to their mates. J. Insect Physiol. 2001, 47, 617–622. [Google Scholar] [CrossRef]
- Pace, K.E.; Baum, L.G. Insect galectins: Roles in immunity and development. Glycoconj. J. 2002, 19, 607–614. [Google Scholar] [CrossRef]
- Tanji, T.; Ohashi-Kobayashi, A.; Natori, S. Participation of a galactose-specific C-type lectin in Drosophila immunity. Biochem. J. 2006, 396, 127–138. [Google Scholar] [CrossRef]
- Findlay, G.D.; Sitnik, J.L.; Wang, W.; Aquadro, C.F.; Clark, N.L.; Wolfner, M.F. Evolutionary rate covariation identifies new members of a protein network required for Drosophila melanogaster female post-mating responses. PLoS Genet. 2014, 10, e1004108. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Tian, C.B.; Liu, S.H.; Wang, T.; Smagghe, G.; Jia, F.X.; Dou, W.; Wang, J.J. Transcriptome analysis to identify genes for peptides and proteins involved in immunity and reproduction from male accessory glands and ejaculatory duct of Bactrocera dorsalis. Peptides 2016, 80, 48–60. [Google Scholar] [CrossRef] [PubMed]
- King, T.P.; Lu, G. Hornet venom allergen antigen 5, Dol m 5: Its T-cell epitopes in mice and its antigenic cross-reactivity with a mammalian testis protein. J. Allergy Clin. Immunol. 1997, 99, 630–639. [Google Scholar] [CrossRef] [PubMed]
- Simske, J.S. Claudins reign: The claudin/EMP/PMP22/gamma channel protein family in C. elegans. Tissue Barriers 2013, 1, e25502. [Google Scholar] [CrossRef]
- Koziel, M.J.; Kowalska, K.; Piastowska-Ciesielska, A.W. Claudins: New players in human fertility and reproductive system cancers. Cancers 2020, 12, 711. [Google Scholar] [CrossRef]
- Chiba, K.; Yamaguchi, K.; Ando, M.; Miyake, H.; Fujisawa, M. Expression pattern of testicular claudin-11 in infertile men. Urology 2012, 80, 1161.e13–1161.e17. [Google Scholar] [CrossRef]
- Newquist, G.; Hogan, J.; Walker, K.; Lamanuzzi, M.; Bowser, M.; Kidd, T. Control of male and female fertility by the netrin axon guidance genes. PLoS ONE 2013, 8, e72524. [Google Scholar] [CrossRef]
- Nie, X.; Dai, X.; Zhao, Y.; Xu, H.; Han, Z.; Jia, R.; Ren, Q.; Huang, X. Identification of three novel Spätzle genes in Eriocheir sinensis and their roles during white spot syndrome virus infection. Fish Shellfish Immunol. 2022, 128, 168–180. [Google Scholar] [CrossRef]
- Jiang, H.; Vilcinskas, A.; Kanost, M.R. Immunity in lepidopteran insects. Adv. Exp. Med. Biol. 2010, 708, 181–204. [Google Scholar]
- Kramer, S.J.; Toschi, A.; Miller, C.A.; Kataoka, H.; Quistad, G.B.; Li, J.P.; Carney, R.L.; Schooley, D.A. Identification of an allatostatin from the tobacco hornworm Manduca sexta. Proc. Natl. Acad. Sci. USA 1991, 88, 9458–9462. [Google Scholar] [CrossRef] [PubMed]
- Jan, A.V. Allatostatin C and its paralog allatostatin double C: The arthropod somatostatins. Insect Biochem. Mol. Biol. 2008, 39, 161–170. [Google Scholar]
- Jan, A.V. Allatostatins C, double C and triple C, the result of a local gene triplication in an ancestral arthropod. Gen. Comp. Endocrinol. 2016, 230–231, 153–157. [Google Scholar]
- Cizmar, D.; Roller, L.; Pillerova, M.; Slama, K.; Zitnan, D. Multiple neuropeptides produced by sex-specific neurons control activity of the male accessory glands and gonoducts in the silkworm Bombyx mori. Sci. Rep. 2019, 9, 2253. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.P. The regulation of expression of insect cuticle protein genes. Insect Biochem. Mol. Biol. 2010, 40, 205–213. [Google Scholar] [CrossRef]
- Lu, J.B.; Guo, J.S.; Chen, X.; Cheng, C.; Luo, X.M.; Zhang, X.Y.; Moussian, B.; Chen, J.P.; Li, J.M.; Zhang, C.X. Chitin synthase 1 and five cuticle protein genes are involved in serosal cuticle formation during early embryogenesis to enhance eggshells in Nilaparvata lugens. Insect Sci. 2022, 29, 363–378. [Google Scholar] [CrossRef]
- Hellsten, E.; Evans, J.P.; Bernard, D.J.; Janne, P.A.; Nussbaum, R.L. Disrupted sperm function and fertilin β processing in mice deficient in the inositol polyphosphate 5-phosphatase Inpp5b. Dev. Biol. 2001, 240, 641–653. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Functional Category | D. melanogaster | A. aegypti | A. mellifera | Heliconius Butterflies |
|---|---|---|---|---|---|
| Cell and extracellular structure | |||||
| Collagen (2) | Cell and extracellular structure | T | |||
| Tetraspanin | Cell and extracellular structure | T | |||
| Metabolism | |||||
| Gamma-glutamylcyclotransferase | Metabolism: amino acid | ||||
| Aromatic-L-amino-acid decarboxylase | Metabolism: amino acid | T | |||
| Vanin-like protein | Metabolism: amino acid | ||||
| Beta-galactosidase | Metabolism: carbohydrate | T | T | ||
| Beta-mannosidase | Metabolism: carbohydrate | T | T | ||
| C-type lysozyme | Metabolism: carbohydrate | ||||
| Chitinase | Metabolism: carbohydrate | T | T | T | |
| Trehalase (2) | Metabolism: carbohydrate | T | |||
| Hyaluronidase | Metabolism: carbohydrate | ||||
| Lipase (2) | Metabolism: lipid | T | T | ||
| Phospholipase | Metabolism: lipid | T | T | ||
| Glycosylphosphatidylinositol-specific phospholipase C | Metabolism: lipid | T | |||
| Palmitoyl-protein thioesterase | Metabolism: lipid | ||||
| Lysophospholipid acyltransferase | Metabolism: lipid | T | |||
| Phospholipid phosphatase | Metabolism: lipid | ||||
| Venom acid phosphatase | Metabolism: lipid | T | T | ||
| Adenosine deaminase (2) | Metabolism: nucleotide | T | T | ||
| Other | |||||
| Protein with chitin binding Peritrophin-A domain | Other: chitin binding | ||||
| Cysteine-rich secretory protein (2) | Other: immune | T | T | T | |
| C-type lectin | Other: immune | T | T | ||
| Galectin | Other: immune | ||||
| Von Willebrand factor | Other: immune | T | |||
| Immune-related protein | Other: immune | ||||
| Gag-pol polyprotein precursor | Other: reverse transcript | ||||
| Reverse transcriptase | Other: reverse transcript | ||||
| Aldo-keto reductase (2) | Other: oxidoreductase | T | |||
| Peptidylglycine alpha-hydroxylating monooxygenase (2) | Other: oxidoreductase | T | |||
| Gamma interferon inducible Lysosomal thiol reductase | Other: oxidoreductase | T | T | ||
| Senecionine N-oxygenase | Other: oxidoreductase | T | |||
| Sulfhydryl oxidase | Other: oxidoreductase | T | T | ||
| GMC oxidoreductase | Other: oxidoreductase | T | T | T | |
| DnaJ-class molecular chaperone | Other: chaperone | T | |||
| Cuticle protein | Other: protection | ||||
| Protein modification machinery | |||||
| Glutaminyl-peptide cyclotransferase | Protein modification machinery | T | |||
| Proteolysis regulators | |||||
| Lysosomal aspartic protease | Proteolysis regulators: protease | T | |||
| Furin-like protease | Proteolysis regulators: protease | T | |||
| Serine carboxypeptidase (3) | Proteolysis regulators: protease | T | |||
| Zinc carboxypeptidase (4) | Proteolysis regulators: protease | T | T | ||
| Trypsin (18) | Proteolysis regulators: protease | T | T | T | T |
| Serpin (7) | Proteolysis regulators: protease inhibitor | T | T | T | |
| Cysteine proteinase inhibitor (4) | Proteolysis regulators: protease inhibitor | T | T | ||
| RNA and protein synthesis | |||||
| Endonuclease | RNA and protein synthesis | T | |||
| Heterogeneous nuclear ribonucleoprotein | RNA and protein synthesis | ||||
| Endoribonuclease (2) | RNA and protein synthesis | ||||
| RNA polymerase II transcription elongation factor | RNA and protein synthesis | ||||
| Polypyrimidine tract-binding protein | RNA and protein synthesis | ||||
| Transcription activator | RNA and protein synthesis | ||||
| Translation initiation factor 4E-binding protein | RNA and protein synthesis | ||||
| Signal transduction | |||||
| Ecdysteroid-regulated 16 kDa protein (2) | Signal transduction | T | T | T | |
| Spaetzle (2) | Signal transduction | ||||
| Nucleobindin | Signal transduction | T | T | ||
| Phosphatidylethanolamine-binding protein | Signal transduction | T | T | T | |
| Chemosensory protein | Signal transduction | T | |||
| PMP-22/EMP/MP20/Claudin tight junction domain-containing protein | Signal transduction | ||||
| Netrin-1 | Signal transduction | ||||
| Allatostatin-CC | Signal transduction | ||||
| Transporters and protein export machinery | |||||
| Type II inositol 1,4,5-trisphosphate 5-phosphatase | Transporters and protein export machinery | ||||
| ADP-ribosylation factor-like protein | Transporters and protein export machinery | T | |||
| Apolipophorin-III | Transporters and protein export machinery | ||||
| Apolipoprotein D | Transporters and protein export machinery | T | T | ||
| Unknown | |||||
| Hypothetical protein (blast) (3) | Unknown | T | T | ||
| Unknown (17) | Unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, L.-J.; Li, F.; Song, Y.; Zhang, Z.-F.; Fan, Y.-L.; Liu, T.-X. Proteome Analysis of Male Accessory Gland Secretions in the Diamondback Moth, Plutella xylostella (Lepidoptera: Plutellidae). Insects 2023, 14, 132. https://doi.org/10.3390/insects14020132
Wu L-J, Li F, Song Y, Zhang Z-F, Fan Y-L, Liu T-X. Proteome Analysis of Male Accessory Gland Secretions in the Diamondback Moth, Plutella xylostella (Lepidoptera: Plutellidae). Insects. 2023; 14(2):132. https://doi.org/10.3390/insects14020132
Chicago/Turabian StyleWu, Li-Juan, Fan Li, Yue Song, Zhan-Feng Zhang, Yong-Liang Fan, and Tong-Xian Liu. 2023. "Proteome Analysis of Male Accessory Gland Secretions in the Diamondback Moth, Plutella xylostella (Lepidoptera: Plutellidae)" Insects 14, no. 2: 132. https://doi.org/10.3390/insects14020132
APA StyleWu, L.-J., Li, F., Song, Y., Zhang, Z.-F., Fan, Y.-L., & Liu, T.-X. (2023). Proteome Analysis of Male Accessory Gland Secretions in the Diamondback Moth, Plutella xylostella (Lepidoptera: Plutellidae). Insects, 14(2), 132. https://doi.org/10.3390/insects14020132