
Identification and Functions of JHE 6 Specifically Expressed in Bombyx mori Silk Gland

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Method
2.1. Experimental Insects
2.2. Bioinformatic Analysis
2.3. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and Quantitative Real Time-PCR (qRT-PCR)
2.4. Polyclonal Antibody Preparation and Western Blotting (WB)
2.5. Immunohistochemistry
2.6. JHA and 20E Treatment
2.7. dsRNA Treatment
2.8. Statistical Analysis
3. Results
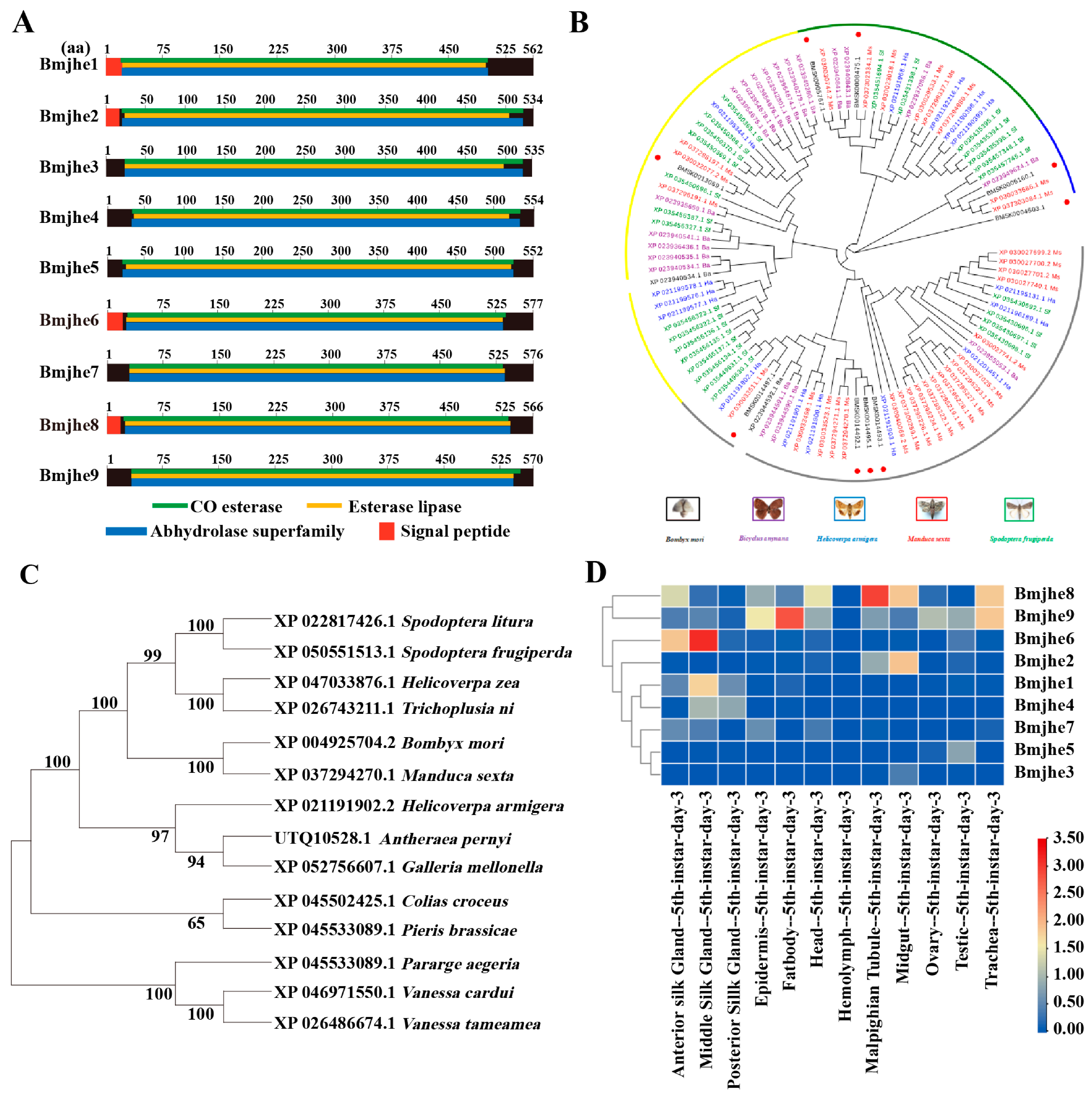
3.1. Bioinformatic Analysis of the JHE Family Members in the Bombyx mori Genome
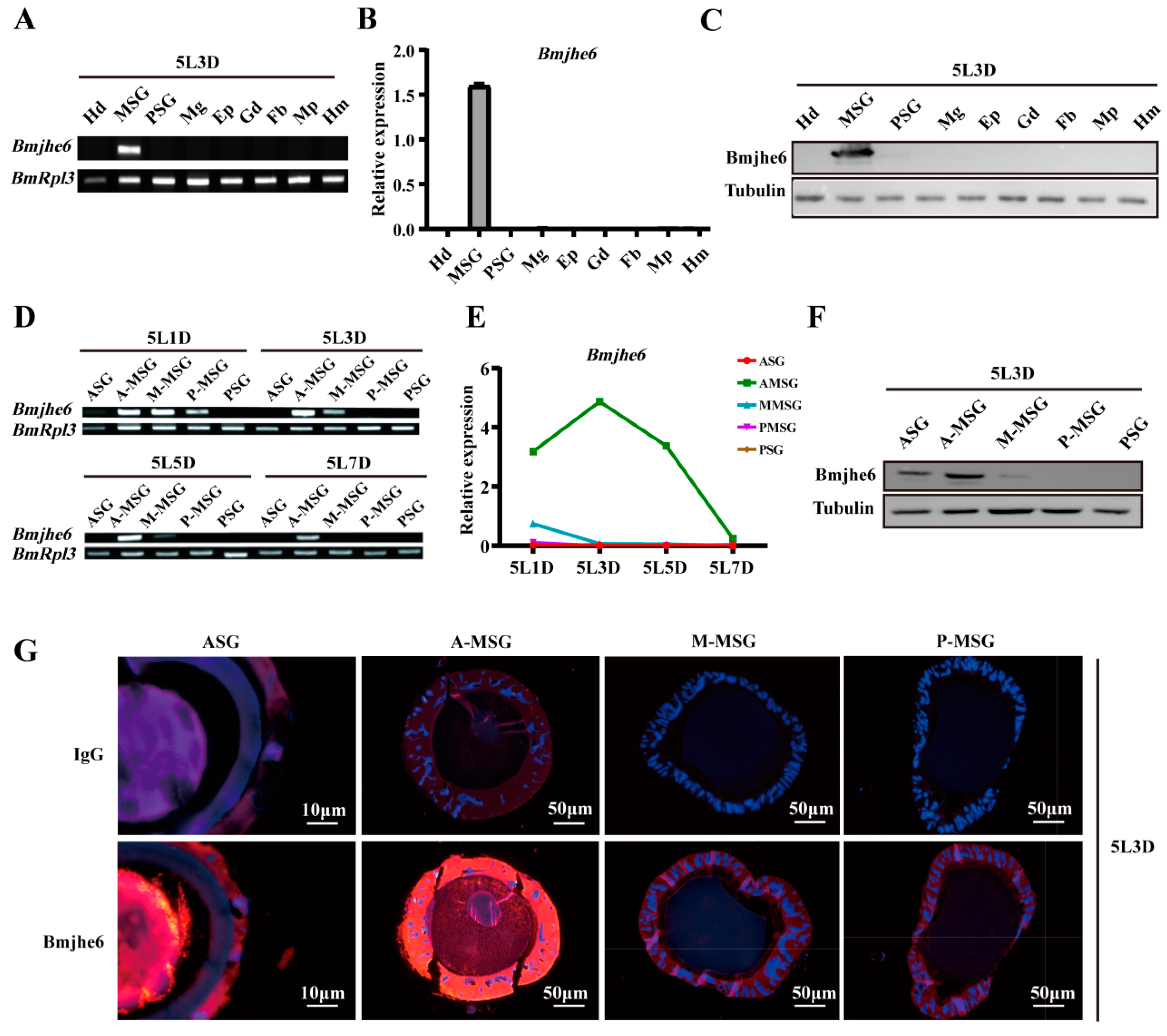
3.2. Bmjhe6 Expression Patterns and Localization
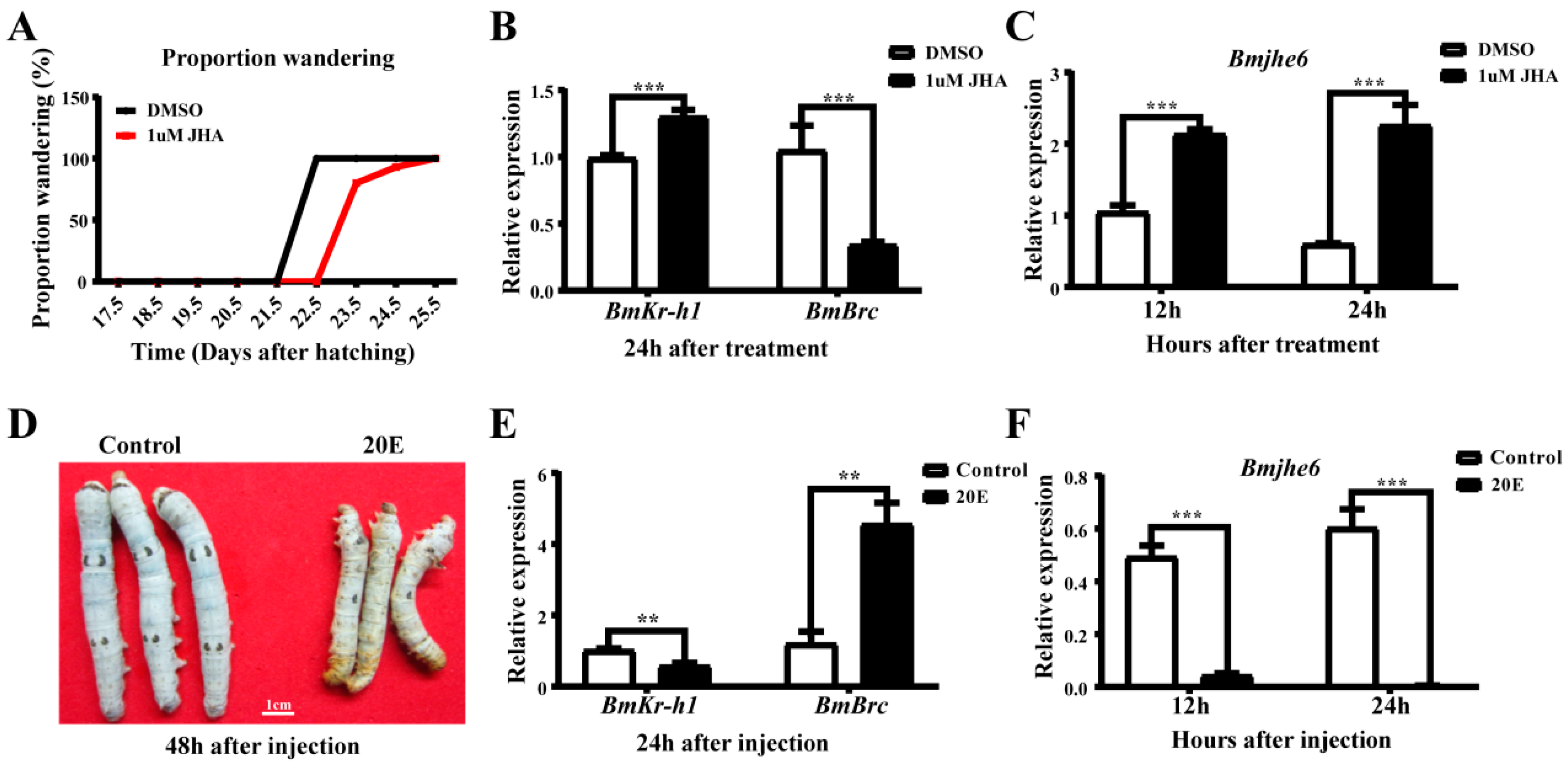
3.3. The Response of Bmjhe6 to JH and 20E
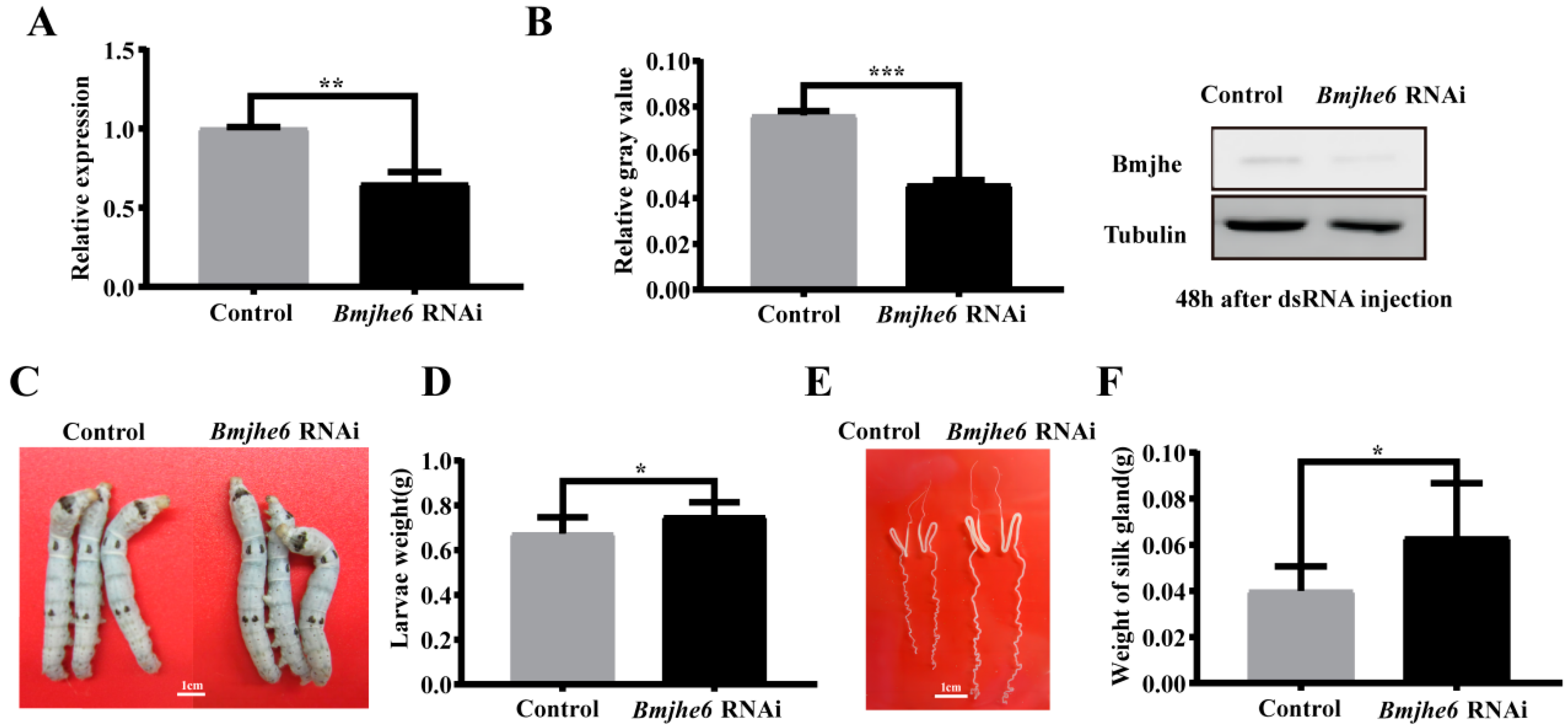
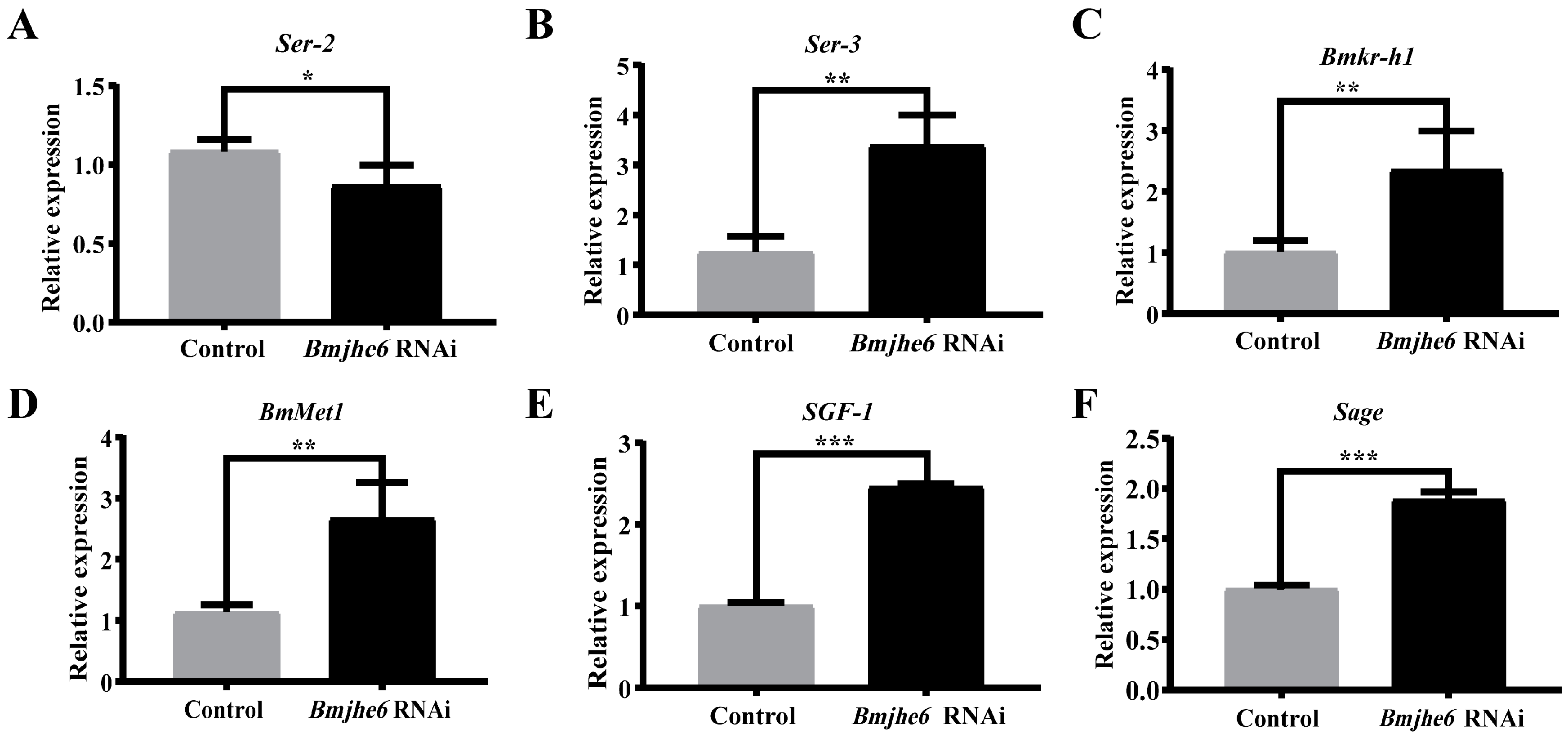
3.4. The Effects on Silk Gland and the Expression of Silk Protein Genes after JHE Knockdown
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, Z.; Tong, C.; Qiu, B.; Yang, H.; Xu, J.; Zheng, S.; Song, Q.; Feng, Q.; Deng, H. 20E-mediated regulation of BmKr-h1 by BmKRP promotes oocyte maturation. BMC Biol. 2021, 19, 39. [Google Scholar] [CrossRef]
- Luo, M.; Li, D.; Wang, Z.; Guo, W.; Kang, L.; Zhou, S. Juvenile hormone differentially regulates two Grp78 genes encoding protein chaperones required for insect fat body cell homeostasis and vitellogenesis. J. Biol. Chem. 2017, 292, 8823–8834. [Google Scholar] [CrossRef] [PubMed]
- Tusun, A.; Li, M.; Liang, X.; Yang, T.; Yang, B.; Wang, G. Juvenile hormone epoxide hydrolase: A promising target for hemipteran pest management. Sci. Rep. 2017, 7, 789. [Google Scholar] [CrossRef]
- Munyiri, F.N.; Ishikawa, Y. Molecular cloning and developmental expression of the gene encoding juvenile hormone esterase in the yellow-spotted longicorn beetle, Psacothea hilaris. Insect Biochem. Mol. Biol. 2007, 37, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Touhara, K.; Bonning, B.C.; Hammock, B.D.; Prestwich, G.D. Action of juvenile hormone (JH) esterase on the JH-JH binding protein complex. An in vitro model of JH metabolism in a caterpillar. Insect Biochem. Mol. Biol. 1995, 25, 727–734. [Google Scholar] [CrossRef]
- Xu, Z.; Yan, R.; Qian, J.; Chen, D.; Guo, Y.; Zhu, G.; Wu, H.; Chen, M. RNAi-mediated knockdown of juvenile hormone esterase causes mortality and malformation in Tribolium castaneum. Entomol. Res. 2022, 52, 476–482. [Google Scholar] [CrossRef]
- Abdel-Aal, Y.A.; Hammock, B.D. Transition state analogs as ligands for affinity purification of juvenile hormone esterase. Science 1986, 233, 1073–1076. [Google Scholar] [CrossRef]
- Venkataraman, V.; O’Mahony, P.J.; Manzcak, M.; Jones, G. Regulation of juvenile hormone esterase gene transcription by juvenile hormone. Dev. Genet. 1994, 15, 391–400. [Google Scholar] [CrossRef]
- Hanzlik, T.N.; Abdel-Aal, Y.A.; Harshman, L.G.; Hammock, B.D. Isolation and sequencing of cDNA clones coding for juvenile hormone esterase from Heliothis virescens: Evidence for a catalytic mechanism for the serine carboxylesterases different from that of the serine proteases. J. Biol. Chem. 1989, 264, 12419–12425. [Google Scholar] [CrossRef]
- Dai, H.; Liu, B.; Yang, L.; Yao, Y.; Liu, M.; Xiao, W.; Li, S.; Ji, R.; Sun, Y. Investigating the Regulatory Mechanism of the Sesquiterpenol Nerolidol from a Plant on Juvenile Hormone-Related Genes in the Insect Spodoptera exigua. Int. J. Mol. Sci. 2023, 24, 13330. [Google Scholar] [CrossRef]
- Yang, J.; Chen, S.; Xu, X.; Lin, S.; Wu, J.; Lin, G.; Bai, J.; Song, Q.; You, M.; Xie, M. Novel miR-108 and miR-234 target juvenile hormone esterase to regulate the response of Plutella xylostella to Cry1Ac protoxin. Ecotoxicol. Environ. Saf. 2023, 254, 114761. [Google Scholar] [CrossRef]
- Shiotsuki, T.; Bonning, B.C.; Hirai, M.; Kikuchi, K.; Hammock, B.D. Characterization and affinity purification of juvenile hormone esterase from Bombyx mori. Biosci. Biotechnol. Biochem. 2000, 64, 1681–1687. [Google Scholar] [CrossRef]
- Hammock, B.D.; Bonning, B.C.; Possee, R.D.; Hanzlik, T.N.; Maeda, S. Expression and effects of the juvenile hormone esterase in a baculovirus vector. Nature 1990, 344, 458–461. [Google Scholar] [CrossRef]
- Hirai, M.; Kamimura, M.; Kikuchi, K.; Yasukochi, Y.; Kiuchi, M.; Shinoda, T.; Shiotsuki, T. cDNA cloning and characterization of Bombyx mori juvenile hormone esterase: An inducible gene by the imidazole insect growth regulator KK-42. Insect Biochem. Mol. Biol. 2002, 32, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, X.; Shiotsuki, T.; Wang, Z.; Xu, X.; Huang, Y.; Li, M.; Li, K.; Tan, A. Depletion of juvenile hormone esterase extends larval growth in Bombyx mori. Insect Biochem. Mol. Biol. 2017, 81, 72–79. [Google Scholar] [CrossRef]
- Tan, A.; Tanaka, H.; Tamura, T.; Shiotsuki, T. Precocious metamorphosis in transgenic silkworms overexpressing juvenile hormone esterase. Proc. Natl. Acad. Sci. USA 2005, 102, 11751–11756. [Google Scholar] [CrossRef]
- Cheng, T.; Zhang, X.; Peng, Z.; Fan, Y.; Zhang, L.; Liu, C. Effects of Osiris9a on Silk Properties in Bombyx mori Determined by Transgenic Overexpression. Int. J. Mol. Sci. 2020, 21, 1888. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Li, S.; Feng, Q. Advances in silkworm studies accelerated by the genome sequencing of Bombyx mori. Annu. Rev. Entomol. 2014, 59, 513–536. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Zheng, H.; Fan, Y.; Cheng, T.; Liu, C. Identification and location of sericin in silkworm with anti-sericin antibodies. Int. J. Biol. Macromol. 2021, 184, 522–529. [Google Scholar] [CrossRef]
- Peng, Z.; Hu, W.; Li, X.; Zhao, P.; Xia, Q. Bending–Spinning Produces Silkworm and Spider Silk with Enhanced Mechanical Properties. Macromolecules 2023, 56, 1199–1212. [Google Scholar] [CrossRef]
- Peng, Z.; Yang, X.; Liu, C.; Dong, Z.; Wang, F.; Wang, X.; Hu, W.; Zhang, X.; Zhao, P.; Xia, Q. Structural and mechanical properties of silk from different instars of Bombyx mori. Biomacromolecules 2019, 20, 1203–1216. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, F.; Akai, H. Insect silk glands: Their types, development and function, and effects of environmental factors and morphogenetic hormones on them. Int. J. Insect Morphol. 1990, 19, 79–132. [Google Scholar] [CrossRef]
- Shigematsu, H.; Moriyama, H. Effect of ecdysterone on fibroin synthesis in the posterior division of the silk gland of the silkworm, Bombyx mori. J. Insect Physiol. 1970, 16, 2015–2022. [Google Scholar] [CrossRef]
- Hamada, Y.; Yamashita, O.; Suzuki, Y. Haemolymph control of sericin gene expression studied by organ transplantation. Cell Differ. 1987, 20, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Liu, Y.L.; Shen, W.D.; Xu, Y.X.; Wei, Z.G. Effect of juvenile hormone and ecdysone treatment on silk protein gene expression in cultured silkglands. Sci. Ser. 2017, 43, 8. [Google Scholar]
- Tripoulas, N.A.; Samols, D. Developmental and hormonal regulation of sericin RNA in the silkworm, Bombyx mori. Dev. Biol. 1986, 116, 328–336. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhao, X.M.; Liu, C.; Jiang, L.J.; Li, Q.Y.; Zhou, M.T.; Cheng, T.C.; Mita, K.; Xia, Q.Y. A juvenile hormone transcription factor Bmdimm-fibroin H chain pathway is involved in the synthesis of silk protein in silkworm, Bombyx mori. J. Biol. Chem. 2015, 290, 972–986. [Google Scholar] [CrossRef]
- Hammock, B. Regulation of juvenile hormone titer: Degradation. In Comprehensive Insect Physiology, Biochemistry, and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: New York, NY, USA, 1985; Volume 7. [Google Scholar]
- Tsubota, T.; Shimomura, M.; Ogura, T.; Seino, A.; Nakakura, T.; Mita, K.; Shinoda, T.; Shiotsuki, T. Molecular characterization and functional analysis of novel carboxyl/cholinesterases with GQSAG motif in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 100–112. [Google Scholar] [CrossRef]
- Cai, R.; Tao, G.; Zhao, P.; Xia, Q.; He, H.; Wang, Y. POU-M2 promotes juvenile hormone biosynthesis by directly activating the transcription of juvenile hormone synthetic enzyme genes in Bombyx mori. Open Biol. 2022, 12, 220031. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| dsRNA | Number of RNAi Silkworm | Survival Rate (%) | Phenotypic Penetrance (%) |
|---|---|---|---|
| Bmjhe6 dsRNA | 30 | 87 | 100 |
| EGFP dsRNA | 30 | 93 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhang, J.; Wu, K.; Yang, H.; Cheng, T.; Liu, C. Identification and Functions of JHE 6 Specifically Expressed in Bombyx mori Silk Gland. Insects 2023, 14, 908. https://doi.org/10.3390/insects14120908
Zhang X, Zhang J, Wu K, Yang H, Cheng T, Liu C. Identification and Functions of JHE 6 Specifically Expressed in Bombyx mori Silk Gland. Insects. 2023; 14(12):908. https://doi.org/10.3390/insects14120908
Chicago/Turabian StyleZhang, Xia, Jikailang Zhang, Keli Wu, Hongguo Yang, Tingcai Cheng, and Chun Liu. 2023. "Identification and Functions of JHE 6 Specifically Expressed in Bombyx mori Silk Gland" Insects 14, no. 12: 908. https://doi.org/10.3390/insects14120908
APA StyleZhang, X., Zhang, J., Wu, K., Yang, H., Cheng, T., & Liu, C. (2023). Identification and Functions of JHE 6 Specifically Expressed in Bombyx mori Silk Gland. Insects, 14(12), 908. https://doi.org/10.3390/insects14120908