
Male and Female Tortricid Moth Response to Non-Pheromonal Semiochemicals


Abstract
:Simple Summary
Abstract
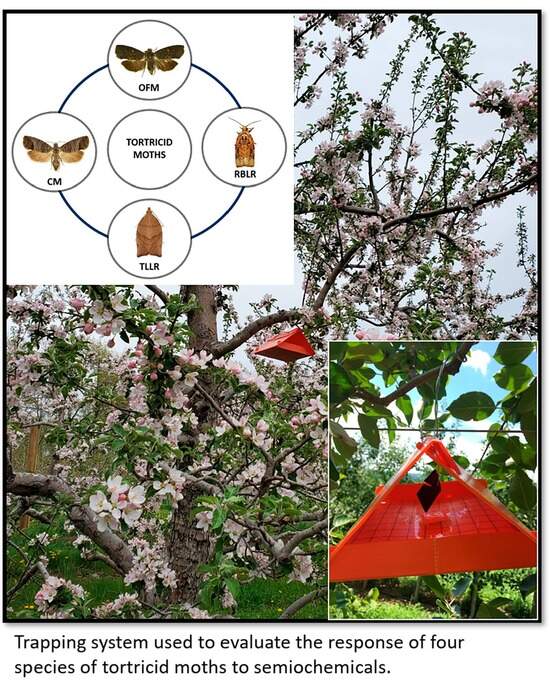
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Odor Treatments and Trap Types
2.3. Data Collection
2.4. Weather Information
2.5. Statistical Analysis
3. Results
3.1. 2020 Study
3.1.1. Early Season Captures (13 May to 19 June)
3.1.2. Mid-Season Captures (20 June to 4 August)
3.1.3. Late Season Captures (5 August to 18 September)
3.2. 2021 Study
3.2.1. Mid-Season Captures (10 June to 21 July)
3.2.2. Late-Season Captures (22 July to 9 September)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapman, P.J.; Lienk, S.E. Tortricid Fauna of Apple in New York (Lepidoptera: Tortricidae); Including an account of apple’s occurrence in the state, especially as a naturalized plant; New York State Agricultural Experiment Station: Geneva, NY, USA, 1971; p. 122. [Google Scholar]
- Lacey, L.A.; Arthurs, S.P.; Knight, A.L.; Huber, J. Microbial control of lepidopteran pests of apple orchards. In Field Manual of Techniques in Invertebrate Pathology: Application and Evaluation of Pathogens for Control of Insects and Other Invertebrate Pests; Springer: Dordrecht, Germany, 2007; pp. 528–546. [Google Scholar]
- Knight, A.L.; Judd, G.J.; Gilligan, T.; Fuentes-Contreras, E.; Walker, W.B. Integrated management of tortricid pests of tree fruit. In Integrated Management of Diseases and Insect Pests of Tree Fruit; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 377–424. [Google Scholar]
- Williamson, E.R.; Folwell, R.J.; Knight, A.L.; Howell, J.F. Economics of employing pheromones for mating disruption of the codling moth, Carpocapsa pomonella. Crop Prot. 1996, 15, 473–477. [Google Scholar] [CrossRef]
- Knight, A.L. Codling moth areawide integrated pest management. In Areawide Pest Management: Theory and Implementation; CABI: Wallingford, UK, 2008; pp. 159–190. [Google Scholar] [CrossRef]
- Knight, A.L.; Mujica, V.; Herrera, S.L.; Tasin, M. Addition of terpenoids to pear ester plus acetic acid increases catches of codling moth (Lepidoptera: Tortricidae). J. Appl. Entomol. 2019, 143, 813–821. [Google Scholar] [CrossRef]
- Knight, A.L.; Brunner, J.F.; Alston, D. Survey of azinphosmethyl resistance in codling moth (Lepidoptera: Tortricidae) in Washington and Utah. J. Econ. Entomol. 1994, 87, 285–292. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A. Semiochemical strategies for tortricid moth control in apple orchards and vineyards in Italy. J. Chem. Ecol. 2016, 42, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Yin, C.; Tang, R.; Chen, M.; Wu, Q.; Huang, C.; Qian, W.; Rota-Stabelli, O.; Yang, N.; Wang, S.; et al. A chromosome-level genome assembly of Cydia pomonella provides insights into chemical ecology and insecticide resistance. Nat. Commun. 2019, 10, 4237. [Google Scholar] [CrossRef]
- Gut, L.J.; Brunner, J.F. Implementing codling moth mating disruption in Washington pome fruit orchards. In Tree Fruit Research Extension Center Information Series; Washington State University: Wenatchee, WA, USA, 1996; Volume 1. [Google Scholar]
- Akotsen-Mensah, C.; Blaauw, B.; Short, B.; Leskey, T.C.; Bergh, J.C.; Polk, D.; Nielsen, A.L. Using IPM-CPR as a management program for apple orchards. J. Econ. Entomol. 2020, 113, 1894–1902. [Google Scholar] [CrossRef]
- Orpet, R.J.; Jones, V.P.; Beers, E.H.; Reganold, J.P.; Goldberger, J.R.; Crowder, D.W. Perceptions and outcomes of conventional vs. organic apple orchard management. Agric. Ecosyst. Environ. 2020, 289, 106723. [Google Scholar] [CrossRef]
- Knight, A.L.; Light, D.M. Developing action thresholds for codling moth (Lepidoptera: Tortricidae) with pear ester-and codlemone-baited traps in apple orchards treated with sex pheromone mating disruption. Can. Entomol. 2005, 137, 739–747. [Google Scholar] [CrossRef]
- Landolt, P.J.; Ohler, B.; Lo, P.; Cha, D.; Davis, T.S.; Suckling, D.M.; Brunner, J. N-Butyl Sulfide as an attractant and coattractant for male and female codling moth (Lepidoptera: Tortricidae). Environ. Entomol. 2014, 43, 291–297. [Google Scholar] [CrossRef]
- Jaffe, B.D.; Guédot, C.; Landolt, P.J. Mass-trapping codling moth, Cydia pomonella (Lepidopteran: Torticidae), using a kairomone lure reduces fruit damage in commercial apple orchards. J. Econ. Entomol. 2018, 111, 1983–1986. [Google Scholar] [CrossRef]
- Light, D.M.; Knight, A.L.; Henrick, C.A.; Rajapaska, D.; Lingren, B.; Dickens, J.C.; Reynolds, K.M.; Buttery, R.G.; Merrill, G.; Roitman, J.; et al. A pear-derived kairomone with pheromonal potency that attracts male and female codling moth, Cydia pomonella (L.). Sci. Nat. 2001, 88, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Stelinski, L.; Gut, L.; Thomson, D. Codling moth management and chemical ecology. Annu. Rev. Entomol. 2008, 53, 503–522. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Brumley, J.A.; Smithhisler, C.L.; Biddick, L.L.; Hofstetter, R.W. Apple fruit infested with codling moth are more attractive to neonate codling moth larvae and possess increased amounts of (E,E)-α-Farnesene. J. Chem. Ecol. 2000, 26, 1685–1699. [Google Scholar] [CrossRef]
- Hern, A.; Dorn, S. Induction of volatile emissions from ripening apple fruits infested with Cydia pomonella and the attraction of adult females. Entomol. Exp. Appl. 2002, 102, 145–151. [Google Scholar] [CrossRef]
- Coracini, M.; Bengtsson, M.; Liblikas, I.; Witzgall, P. Attraction of codling moth males to apple volatiles. Entomol. Exp. Appl. 2004, 110, 1–10. [Google Scholar] [CrossRef]
- Natale, D.; Mattiacci, L.; Hern, A.; Pasqualini, E.; Dorn, S. Response of female Cydia molesta (Lepidoptera: Tortricidae) to plant derived volatiles. Bull. Entomol. Res. 2003, 93, 335–342. [Google Scholar] [CrossRef]
- Natale, D.; Mattiacci, L.; Pasqualini, E.; Dorn, S. Apple and peach fruit volatiles and the apple constituent butyl hexanoate attract female oriental fruit moth, Cydia molesta, in the laboratory. J. Appl. Entomol. 2004, 128, 22–27. [Google Scholar] [CrossRef]
- Piñero, J.C.; Dorn, S. Synergism between aromatic compounds and green leaf volatiles derived from the host plant underlies female attraction in the oriental fruit moth. Entomol. Exp. Appl. 2007, 125, 185–194. [Google Scholar] [CrossRef]
- Il’ichev, A.L.; Kugimiya, S.; Williams, D.G.; Takabayashi, J. Volatile compounds from young peach shoots attract males of oriental fruit moth in the field. J. Plant Interact. 2009, 4, 289–294. [Google Scholar] [CrossRef]
- Varela, N.; Avilla, J.; Gemeno, C.; Anton, S. Ordinary glomeruli in the antennal lobe of male and female tortricid moth Grapholita molesta (Busck) (Lepidoptera: Tortricidae) process sex pheromone and host-plant volatiles. J. Exp. Biol. 2011, 214, 637–645. [Google Scholar] [CrossRef]
- Najar-Rodriguez, A.; Orschel, B.; Dorn, S. Season-long volatile emissions from peach and pear trees in situ, overlapping profiles, and olfactory attraction of an oligophagous fruit moth in the laboratory. J. Chem. Ecol. 2013, 39, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.K.; Hull, L.A.; Rajotte, E.G.; Krawczyk, G.; Bohnenblust, E. Evaluating sex-pheromone-and kairomone-based lures for attracting codling moth adults in mating disruption versus conventionally managed apple orchards in Pennsylvania. Pest Manag. Sci. 2011, 67, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Landolt, P.J.; Suckling, D.M.; Judd, G.J.R. Positive interaction of a feeding attractant and a host kairomone for trapping the codling moth, Cydia pomonella (L.). J. Chem. Ecol. 2007, 33, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L.; Light, D.M.; Trimble, R.M. Identifying (E)-4, 8-Dimethyl-1, 3, 7-Nonatriene plus acetic acid as a new lure for male and female codling moth (Lepidoptera: Tortricidae). Environ. Entomol. 2011, 40, 420–430. [Google Scholar] [CrossRef]
- Lu, P.F.; Huang, L.Q.; Wang, C.Z. Identification and field evaluation of pear fruit volatiles attractive to the oriental fruit moth, Cydia molesta. J. Chem. Ecol. 2012, 38, 1003–1016. [Google Scholar] [CrossRef]
- Barros-Parada, W.; Ammagarahalli, B.; Basoalto, E.; Fuentes-Contreras, E.; Gemeno, C. Captures of oriental fruit moth, Grapholita molesta (Lepidoptera: Tortricidae), in traps baited with host-plant volatiles in Chile. Appl. Entomol. Zool. 2018, 53, 193–204. [Google Scholar] [CrossRef]
- Knight, A.L.; Basoalto, E.; Hilton, R.; Molinari, F.; Zoller, B.; Hansen, R.; Krawczyk, G.; Hull, L. Monitoring oriental fruit moth (Lepidoptera: Tortricidae) with the Ajar bait trap in orchards under mating disruption. J. Appl. Entomol. 2013, 137, 650–660. [Google Scholar] [CrossRef]
- Cichon, L.; Fuentes-Contreras, E.; Garrido, S.; Lago, J.; Barros-Parada, W.; Basoalto, E.; Hilton, H.; Knight, A.L. Monitoring oriental fruit moth (Lepidoptera: Tortricidae) with sticky traps baited with terpinyl acetate and sex pheromone. J. Appl. Entomol. 2013, 137, 275–281. [Google Scholar] [CrossRef]
- Knight, A.L.; Hilton, R.; Basoalto, E.; Stelinski, L.L. Use of glacial acetic acid to enhance bisexual monitoring of tortricid pests with kairomone lures in pome fruits. Environ. Entomol. 2014, 43, 1628–1640. [Google Scholar] [CrossRef]
- Mujica, V.; Preti, M.; Basoalto, E.; Cichon, L.; Fuentes-Contreras, E.; Barros-Parada, W.; Krawczyk, G.; Nunes, M.Z.; Walgenbach, J.F.; Hansen, R.; et al. Improved monitoring of oriental fruit moth (Lepidoptera: Tortricidae) with terpinyl acetate plus acetic acid membrane lures. J. Appl. Entomol. 2018, 142, 731–744. [Google Scholar] [CrossRef]
- Padilha, A.C.; Arioli, C.J.; Boff, M.I.C.; Rosa, J.M.; Botton, M. Traps and baits for luring Grapholita molesta (Busck) adults in mating disruption-treated apple orchards. Neotrop. Entomol. 2018, 47, 152–159. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Guo, Y.F.; Chai, X.H.; Li, J.; Ma, R.Y. Behavioral effects of different attractants on adult male and female oriental fruit moths, Grapholita molesta. Pest Manag. Sci. 2020, 76, 3225–3235. [Google Scholar] [CrossRef]
- Knight, A.L.; El-Sayed, A.M.; Judd, G.J.R.; Basoalto, E. Development of 2-phenylethanol plus acetic acid lures to monitor obliquebanded leafroller (Lepidoptera: Tortricidae) under mating disruption. J. Appl. Entomol. 2017, 141, 729–739. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Knight, A.L.; Basoalto, E.; Suckling, D.M. Caterpillar-induced plant volatiles attract conspecific herbivores and a generalist predator. J. Appl. Entomol. 2018, 142, 495–503. [Google Scholar] [CrossRef]
- Judd, G.J.; Knight, A.L.; El-Sayed, A.M. Trapping Pandemis limitata (Lepidoptera: Tortricidae) moths with mixtures of acetic acid, caterpillar-induced apple-leaf volatiles, and sex pheromone. Can. Entomol. 2017, 149, 813–822. [Google Scholar] [CrossRef]
- Preti, M.; Knight, A.L.; Favaro, R.; Basoalto, E.; Tasin, M.; Angeli, S. Comparison of new kairomone-based lures for Cydia pomonella (Lepidoptera: Tortricidae) in Italy and USA. Insects 2021, 12, 72. [Google Scholar] [CrossRef]
- Löfstedt, C. Population variation and genetic control of pheromone communication systems in moths. Entomol. Exp. Appl. 1990, 54, 199–218. [Google Scholar] [CrossRef]
- Ioriatti, C.; Molinari, F.; Pasqualini, E.; De Cristofaro, A.; Schmidt, S.; Espinha, I. The plant volatile attractant (E, Z)-2, 4-Ethyl-Decadienoate (DA2313) for codling moth monitoring. Bull. Zool. Agric. 2003, 35, 127–137. [Google Scholar]
- Trimble, R.M.; El-Sayed, A.M. Potential of ethyl (2E, 4Z)-2, 4-decadienoate for monitoring activity of codling moth (Lepidoptera: Tortricidae) in eastern North American apple orchards. Can. Entomol. 2005, 137, 110–116. [Google Scholar] [CrossRef]
- Meraner, A.; Brandstätter, A.; Thaler, R.; Aray, B.; Unterlechner, M.; Niederstätter, H.; Parson, W.; Zelger, R.; Dalla Via, J.; Dallinger, R. Molecular phylogeny and population structure of the codling moth (Cydia pomonella) in Central Europe: I. Ancient clade splitting revealed by mitochondrial haplotype markers. Mol. Phylogenetics Evol. 2008, 48, 825–837. [Google Scholar] [CrossRef]
- Mitchell, V.J.; Manning, L.-A.; Cole, L.; Suckling, D.M.; El-Sayed, A.M. Efficacy of the pear ester as a monitoring tool for codling moth Cydia pomonella (Lepidoptera: Tortricidae) in New Zealand Apple Orchards. Pest Manag. Sci. 2008, 64, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Dekker, T.; Kárpáti, Z. Coding and evolution of pheromone preference in moths. In Insect Sex Pheromone Research and Beyond. Entomology Monographs; Ishikawa, Y., Ed.; Springer: Singapore, 2020; pp. 265–286. [Google Scholar] [CrossRef]
- Piñero, J.C.; Giovanni Galizia, C.; Dorn, S. Synergistic behavioral responses of female oriental fruit moths (Lepidoptera: Tortricidae) to synthetic host plant-derived mixtures are mirrored by odor-evoked calcium activity in their antennal lobes. J. Insect Physiol. 2008, 54, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Myers, C.T.; Krawczyk, G.; Agnello, A.M. Response of tortricid moths and non-target insects to pheromone trap color in commercial apple orchards. J. Entomol. Sci. 2009, 44, 69–77. [Google Scholar] [CrossRef]
- Fuková, I.; Neven, L.G.; Bárcenas, N.M.; Gund, N.A.; Dalíková, M.; Marec, F. Rapid assessment of the sex of codling moth Cydia pomonella (Linnaeus) (Lepidoptera: Tortricidae) Eggs and Larvae. J. Appl. Entomol. 2009, 133, 249–261. [Google Scholar] [CrossRef]
- Shang, S. Morphological characteristics of reproductive system of the codling moth Cydia pomonella. ARTOAJ 2021, 25, 001–006. [Google Scholar] [CrossRef]
- Schumacher, P.; WEyENETH, A.; Weber, D.C.; Dorn, S. Long flights in Cydia pomonella L. (Lepidoptera: Tortricidae) measured by a flight mill: Influence of sex, mated status and age. Physiol. Entomol. 1997, 22, 149–160. [Google Scholar] [CrossRef]
- Boivin, T.; Chadœuf, J.; Bouvier, J.; Beslay, D.; Sauphanor, B. Modelling the interactions between phenology and insecticide resistance genes in the codling moth Cydia pomonella. Pest Manag. Sci. 2005, 61, 53–67. [Google Scholar] [CrossRef]
- Gu, H.; Hughes, J.; Dorn, S. Trade-off between mobility and fitness in Cydia pomonella L.(Lepidoptera: Tortricidae). Ecol. Entomol. 2006, 31, 68–74. [Google Scholar] [CrossRef]
- Timm, A.E.; Geertsema, H.; Warnich, L. Gene flow among Cydia pomonella (Lepidoptera: Tortricidae) geographic and host populations in South Africa. J. Econ. Entomol. 2006, 99, 341–348. [Google Scholar] [CrossRef]
- Fieser, L.F.; Fieser, M. Organic Chemistry; D. C. Heath and Company: Boston, MA, USA, 1944; pp. 368–369. [Google Scholar]
- Opgrande, J.L.; Dobratz, C.J.; Brown, E.; Liang, J.; Conn, G.S.; Shelton, F.J.; With, J. Benzaldehyde. In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000. [Google Scholar]
- Haynes, K.F.; Zhao, J.Z.; Latif, A. Identification of floral compounds from Abelia grandiflora that stimulate upwind flight in cabbage looper moths. J. Chem. Ecol. 1991, 17, 637–646. [Google Scholar] [CrossRef]
- Honda, K.; Ômura, H.; Hayashi, N. Identification of floral volatiles from Ligustrum Japonicum that stimulate flower-visiting by cabbage butterfly, Pieris Rapae. J. Chem. Ecol. 1998, 24, 2167–2180. [Google Scholar] [CrossRef]
- Bruce, T.J.; Cork, A. Electrophysiological and behavioral responses of female Helicoverpa armigera to compounds identified in flowers of African marigold, Tagetes erecta. J. Chem. Ecol. 2001, 27, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.C.; Prokopy, R.J. Field Evaluation of plant odor and pheromonal combinations for attracting plum curculios. J. Chem. Ecol. 2003, 29, 2735–2748. [Google Scholar] [CrossRef] [PubMed]
- Lohonyai, Z.; Vuts, J.; Kárpáti, Z.; Koczor, S.; Domingue, M.J.; Fail, J.; Birkett, M.A.; Tóth, M.; Imrei, Z. Benzaldehyde: An alfalfa-related compound for the spring attraction of the pest weevil Sitona humeralis (Coleoptera: Curculionidae). Pest Manag. Sci. 2019, 75, 3153–3159. [Google Scholar] [CrossRef]
- Ethington, M.W.; Hughes, G.P.; VanDerLaan, N.R.; Ginzel, M.D. Chemically-mediated colonization of black cherry by the peach bark beetle, Phloeotribus liminaris. J. Chem. Ecol. 2021, 47, 303–312. [Google Scholar] [CrossRef]
- Xiang, H.-M.; Ma, R.-Y.; Diao, H.-L.; Li, X.-W.; He, X.-J.; Guo, Y.-F. Peach-specific aldehyde nonanal attracts female oriental fruit moths, Grapholita molesta (Lepidoptera: Tortricidae). J. Asia Pac. Entomol. 2017, 20, 1419–1424. [Google Scholar] [CrossRef]
- Vallat, A.; Dorn, S. Changes in volatile emissions from apple trees and associated response of adult female codling moths over the fruit-growing season. J. Agric. Food Chem. 2005, 53, 4083–4090. [Google Scholar] [CrossRef]
- Buchbauer, G.; Jirovetz, L.; Wasicky, M.; Nikiforov, A. Headspace and essential oil analysis of apple flowers. J. Agric. Food Chem. 1993, 41, 116–118. [Google Scholar] [CrossRef]
- Knight, A.L.; Cichon, L.; Lago, J.; Fuentes-Contreras, E.; Barros-Parada, W.; Hull, L.; Krawczyk, G.; Zoller, B.; Hansen, R.; Basoalto, E. Monitoring oriental fruit moth and codling moth (Lepidoptera: Tortricidae) with combinations of pheromones and kairomones. J. Appl. Entomol. 2014, 138, 783–794. [Google Scholar] [CrossRef]
- Allred, D.B. Responses of Males to a Pheromone Blend of Female Oriental Fruit Moth with and without E8, E10-Dodecadien-1-ol, a Pheromone Component of Codling Moth (Lepidoptera: Tortricidae). Doctoral Dissertation, Oregon State University, Corvallis, OR, USA, 1995. [Google Scholar]
- Knight, A.L.; Basoalto, E.; Stelinski, L.L. Variability in the efficacy of sex pheromone lures for monitoring oriental fruit moth (Lepidoptera: Tortricidae). J. Appl. Entomol. 2016, 140, 261–267. [Google Scholar] [CrossRef]
- Zhang, G.; Chen, J.; Yu, H.; Tian, X.; Wu, J. Molecular and functional characterization of pheromone binding protein 1 from the oriental fruit moth, Grapholita molesta (Busck). Sci. Rep. 2018, 8, 2276. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lure | Description | |
|---|---|---|
| 1 | Pherocon® CM L2-P | Codling moth pheromone lure (positive control) |
| 2 | Pherocon® Megalure CM 4K Dual® | Pear ester, acetic acid, (E)-4,8-dimethyl-1,3,7-nonatriene (DMNT) and PyrLOX [3] |
| 3 | Pherocon® Megalure CM 4K Dual® + benzaldehyde | Blend of four kairomones + benzaldehyde |
| 4 | TRE 2266 | Linalool oxide + DMNT |
| 5 | TRE 2267 | TRE 2266 + benzaldehyde |
| 6 | Unbaited control | Negative control |
| Lures | Average Number of Female Moths Captured per Season per Trap | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2020 (n = 8) | 2021 (n = 10) | |||||||||
| OFM | CM | OFM | CM | |||||||
| Early | Mid | Late | Early | Mid | Late | Mid | Late | Mid | Late | |
| CM L2-P | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Megalure | 1.13 | 0 | 0.13 | 0 | 1 | 0.13 | 0.2 | 0.7 | 1.4 | 1.6 |
| Megalure + benzaldehyde | 0.63 | 0.13 | 0.38 | 0 | 0.25 | 0 | 0.2 | 1.2 | 2.2 | 0.8 |
| TRE 2266 | 0.13 | 0 | 0.13 | 0 | 0.13 | 0.13 | 0 | 0.1 | 0 | 0.4 |
| TRE 2267 | 0.38 | 0 | 0 | 0 | 0 | 0 | 0 | 0.4 | 0.3 | 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giri, A.P.; Short, B.D.; Piñero, J.C. Male and Female Tortricid Moth Response to Non-Pheromonal Semiochemicals. Insects 2023, 14, 884. https://doi.org/10.3390/insects14110884
Giri AP, Short BD, Piñero JC. Male and Female Tortricid Moth Response to Non-Pheromonal Semiochemicals. Insects. 2023; 14(11):884. https://doi.org/10.3390/insects14110884
Chicago/Turabian StyleGiri, Ajay P., Brent D. Short, and Jaime C. Piñero. 2023. "Male and Female Tortricid Moth Response to Non-Pheromonal Semiochemicals" Insects 14, no. 11: 884. https://doi.org/10.3390/insects14110884
APA StyleGiri, A. P., Short, B. D., & Piñero, J. C. (2023). Male and Female Tortricid Moth Response to Non-Pheromonal Semiochemicals. Insects, 14(11), 884. https://doi.org/10.3390/insects14110884