

First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae)
 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Yellow Mealworms Colony
2.2. Substrate Composition/Preparation
2.3. Experimental Set-up
2.4. Mealworm Growth Perfomance
2.5. Carotenoids Analysis
- 0.55 = the final hexane layer volume ratio to the volume of mixed solvents added for hexane:acetone: ethanol (2:1:1);
- W (mg) = the weight of sample analyzed;
- V (mL) = the volume of mixed solvents added;
- 537 = the molecular weights of lycopene and β-carotene (g/mol);
- 6.95, 1.59, 9.38, and 6.70 = numerical coefficients obtained by solving the Lambert–Beer equations, which link the value of the concentrations of lycopene and β-Carotene to the absorbance and the respective ε (molar extinction coefficients) to the two established wavelengths (444 nm and 503 nm) [57].
2.6. Lipid Analysis in Mealworms
2.7. Nutritional Quality Indices
2.8. Protein Analysis
2.9. Statistical Analysis
3. Results
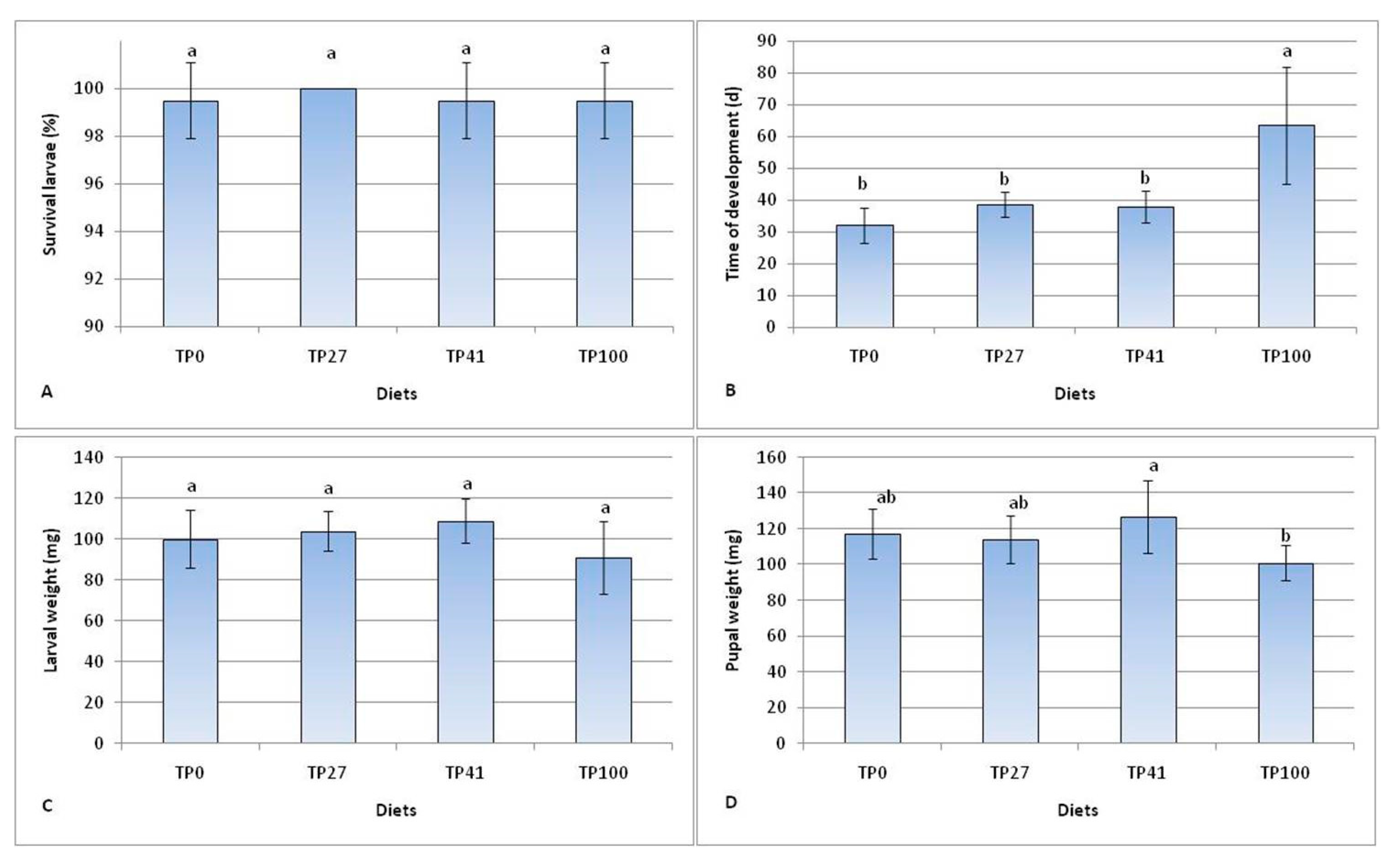
3.1. Larval Performances
3.2. Efficiency Indicators
3.3. Lycopene and β-Carotene Quantification
3.4. Larval Nutritional Value
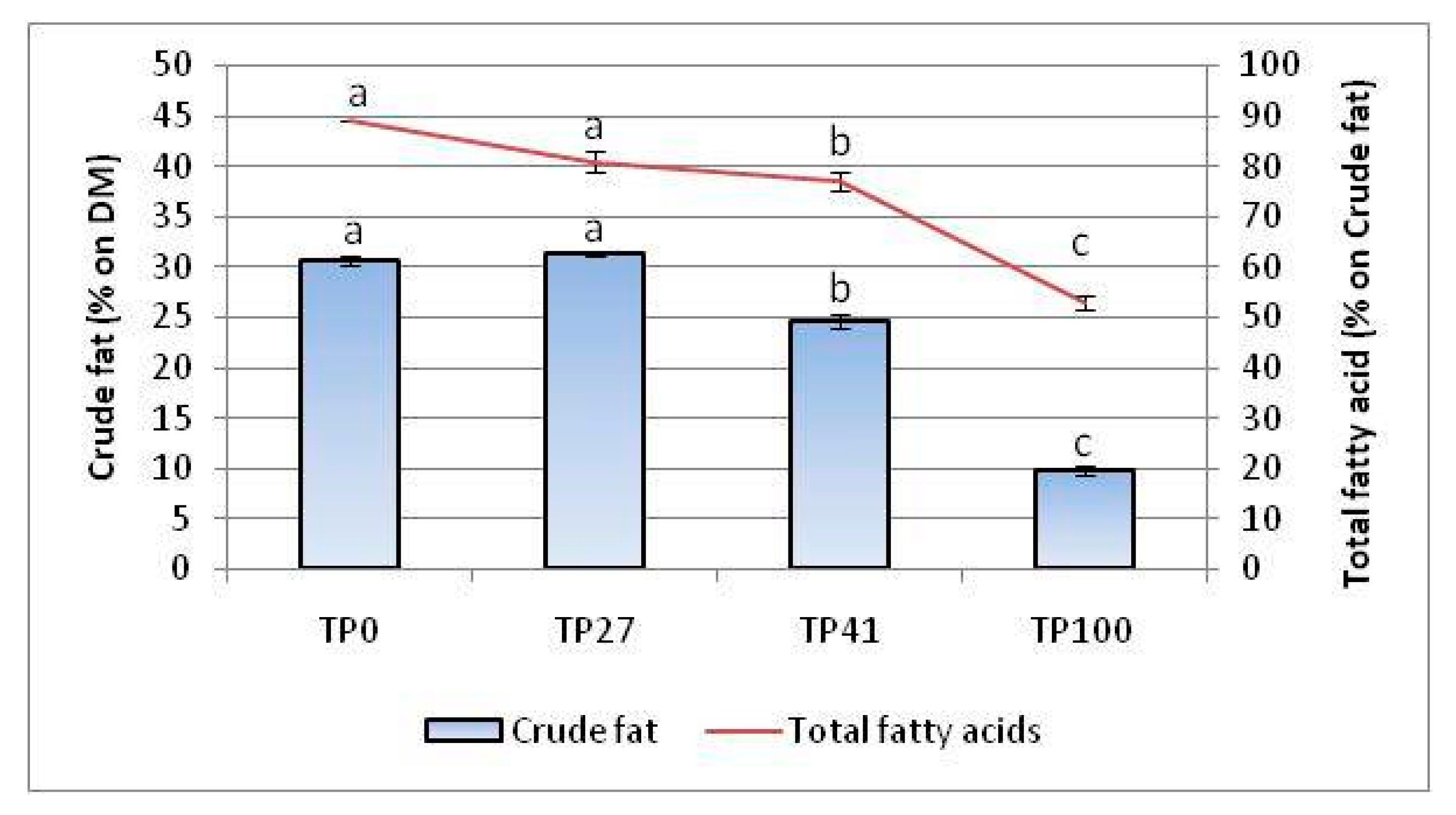
3.5. Lipid Quality Indices
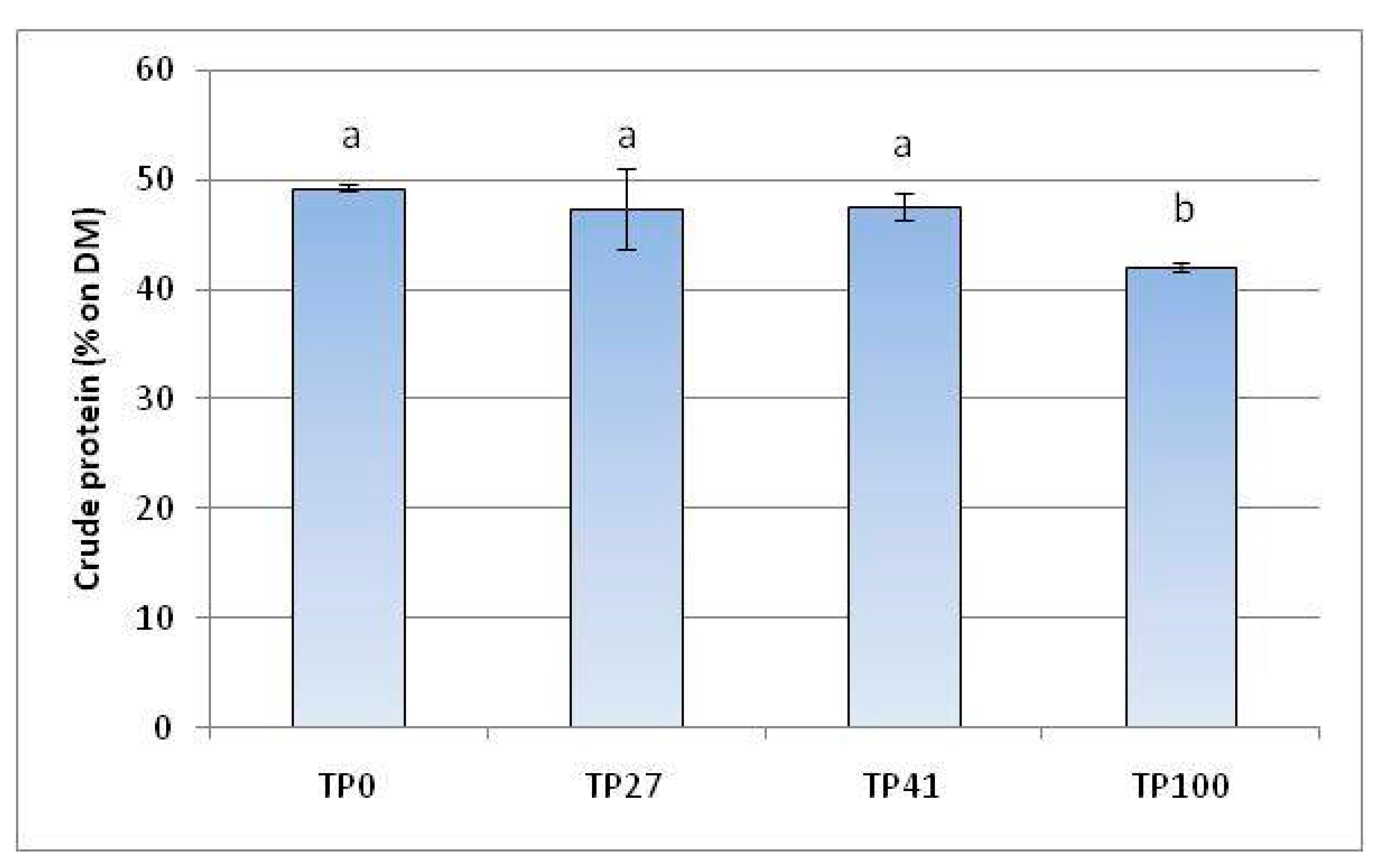
3.6. Crude Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect Rearing: Potential, Challenges, and Circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Jensen, H.; Elleby, C.; Domínguez, I.P.; Chatzopoulos, T.; Charlebois, P. Insect-Based Protein Feed: From Fork to Farm. J. Insects Food Feed 2021, 7, 1219–1233. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An Exploration on Greenhouse Gas and Ammonia Production by Insect Species Suitable for Animal or Human Consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef]
- Amrul, N.F.; Kabir Ahmad, I.; Ahmad Basri, N.E.; Suja, F.; Abdul Jalil, N.A.; Azman, N.A. A Review of Organic Waste Treatment Using Black Soldier Fly (Hermetia illucens). Sustainability 2022, 14, 4565. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; Camenzuli, L.; van der Lee, M.K.; Oonincx, D.G.A.B. Uptake of Cadmium, Lead and Arsenic by Tenebrio molitor and Hermetia illucens from Contaminated Substrates. PLoS ONE 2016, 11, e0166186. [Google Scholar] [CrossRef]
- Ribeiro, N.T.G.M. Tenebrio molitor for Food or Feed: Rearing Conditions and the Effects of Pesticides on Its Performance. Ph.D. Thesis, Escola Superior Agrária de Coimbra, Coimbra, Portugal, 2017. [Google Scholar]
- Houbraken, M.; Spranghers, T.; De Clercq, P.; Cooreman-Algoed, M.; Couchement, T.; De Clercq, G.; Verbeke, S.; Spanoghe, P. Pesticide Contamination of Tenebrio molitor (Coleoptera: Tenebrionidae) for Human Consumption. Food Chem. 2016, 201, 264–269. [Google Scholar] [CrossRef]
- Lee, M.; Simakova, A.V.; Babkina Irina, B.; Bagirov, R.T.; Frank, Y.A. Interaction of Microplastics and Terrestrial and Aquatic Insects (Bioaccumulation, Degradation, Ecotoxicological Effects). Acta Biol. Sib. 2023, 6, 549–564. [Google Scholar]
- Bordiean, A.; Krzyżaniak, M.; Aljewicz, M.; Stolarski, M.J. Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods 2022, 11, 3075. [Google Scholar] [CrossRef] [PubMed]
- El Deen, S.N.; Lamaj, F.; Verrastro, V.; Al Bitar, L.; Baldacchino, F. Effects of Two Diets on Adults’ Survival and Productivity in Mass-Rearing of Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Insects Food Feed 2021, 7, 1149–1157. [Google Scholar] [CrossRef]
- Noyens, I.; Schoeters, F.; Van Peer, M.; Berrens, S.; Goossens, S.; Van Miert, S. The Nutritional Profile, Mineral Content and Heavy Metal Uptake of Yellow Mealworm Reared with Supplementation of Agricultural Sidestreams. Sci. Rep. 2023, 13, 11604. [Google Scholar] [CrossRef]
- Kröncke, N.; Benning, R. Influence of Dietary Protein Content on the Nutritional Composition of Mealworm Larvae (Tenebrio molitor L.). Insects 2023, 14, 261. [Google Scholar] [CrossRef]
- Riekkinen, K.; Väkeväinen, K.; Korhonen, J. The Effect of Substrate on the Nutrient Content and Fatty Acid Composition of Edible Insects. Insects 2022, 13, 590. [Google Scholar] [CrossRef] [PubMed]
- Dreassi, E.; Cito, A.; Zanfini, A.; Materozzi, L.; Botta, M.; Francardi, V. Dietary Fatty Acids Influence the Growth and Fatty Acid Composition of the Yellow Mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). Lipids 2017, 52, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Francardi, V.; Cito, A.; Fusi, S.; Botta, M.; Dreassi, E. Linseed To Increase N-3 Fatty Acids in Tenebrio molitor (Coleoptera Tenebrionidae). Redia 2017, 100, 73–76. [Google Scholar] [CrossRef]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Adamaki-Sotiraki, C.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Waste Orange Peels as a Feed Additive for the Enhancement of the Nutritional Value of Tenebrio molitor. Foods 2023, 12, 783. [Google Scholar] [CrossRef]
- Rossi, G.; Mattioli, S.; Rondoni, G.; Bosco, A.D.; Servili, M.; Castellini, C.; Conti, E. Characterisation of Fatty Acid Profiles of Tenebrio Molitor Larvae Reared on Diets Enriched with Edible Oils. J. Insects Food Feed 2022, 8, 901–912. [Google Scholar] [CrossRef]
- Anderson, S.J. Increasing Calcium Levels in Cultured Insects. Zoo Biol. 2000, 19, 1–9. [Google Scholar] [CrossRef]
- Latney, L.V.; Toddes, B.D.; Wyre, N.R.; Brown, D.C.; Michel, K.E.; Briscoe, J.A. Effects of Various Diets on the Calcium and Phosphorus Composition of Mealworms (Tenebrio molitor Larvae) and Superworms (Zophobas Morio Larvae). Am. J. Vet. Res. 2017, 78, 178–185. [Google Scholar] [CrossRef]
- Brai, A.; Vagaggini, C.; Pasqualini, C.; Poggialini, F.; Tarchi, F.; Francardi, V.; Dreassi, E. Use of Distillery By-Products as Tenebrio Molitor Mealworm Feed Supplement. J. Insects Food Feed 2023, 9, 611–623. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 15 October 2023).
- World Processing Tomato Council. Available online: www.wptc.to/production/ (accessed on 20 August 2023).
- Tamošiūnas, A.; Khiari, B.; Jeguirim, M. Biofuels Production: Biogas, Biodiesel and Bioethanol from Tomato Wastes. In Tomato Processing by-Products; Elsevier: Amsterdam, The Netherlands, 2022; pp. 333–370. [Google Scholar]
- Lu, Z.; Wang, J.; Gao, R.; Ye, F.; Zhao, G. Sustainable Valorisation of Tomato Pomace: A Comprehensive Review. Trends Food Sci. Technol. 2019, 86, 172–187. [Google Scholar] [CrossRef]
- Lamidi, R.O.; Jiang, L.; Pathare, P.B.; Wang, Y.D.; Roskilly, A.P. Recent Advances in Sustainable Drying of Agricultural Produce: A Review. Appl. Energy 2019, 233–234, 367–385. [Google Scholar] [CrossRef]
- Fondevila, M.; Guada, J.A.; Gasa, J.; Castrillo, C. Tomato Pomace as a Protein Supplement for Growing Lambs. Small Rumin. Res. 1994, 13, 117–126. [Google Scholar] [CrossRef]
- Heuzé, V.; Tran, G.; Hassoun, P.; Bastianelli, D.; Lebas, F. Tomato Pomace, Tomato Skins and Tomato Seeds; Food and Agriculture Organization: Rome, Italy, 2021. [Google Scholar]
- Del Valle, M.; Cámara, M.; Torija, M.-E. Chemical Characterization of Tomato Pomace. J. Sci. Food Agric. 2006, 86, 1232–1236. [Google Scholar] [CrossRef]
- Knoblich, M.; Anderson, B.; Latshaw, D. Analyses of Tomato Peel and Seed Byproducts and Their Use as a Source of Carotenoids. J. Sci. Food Agric. 2005, 85, 1166–1170. [Google Scholar] [CrossRef]
- Nesci, S.; Spagnoletta, A.; Oppedisano, F. Inflammation, Mitochondria and Natural Compounds Together in the Circle of Trust. Int. J. Mol. Sci. 2023, 24, 6106. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic Effects of Inflammation on Vitamin A and Carotenoids in Humans and Animal Models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- Grune, T.; Lietz, G.; Palou, A.; Ross, A.C.; Stahl, W.; Tang, G.; Thurnham, D.; Yin, S.; Biesalski, H.K. β-Carotene Is an Important Vitamin A Source for Humans. J. Nutr. 2010, 140, 2268S–2285S. [Google Scholar] [CrossRef]
- de Almeida Torres, R.J.; dos Anjos Ferreira, A.L.; Luchini, A.; de Almeida Torres, R.J.; Correa, C.R. The Role of Non-Enzymatic Antioxidants on Age-Related Macular Degeneration. Front. Drug Chem. Clin. Res 2022, 5, 1–19. [Google Scholar]
- Heinrich, U.; Wiebusch, M.; Tronnier, H.; Gärtner, C.; Eichler, O.; Sies, H.; Stahl, W. Supplementation with β-Carotene or a Similar Amount of Mixed Carotenoids Protects Humans from UV-Induced Erythema. J. Nutr. 2003, 133, 98–101. [Google Scholar] [CrossRef]
- Palozza, P.; Simone, R.E.; Catalano, A.; Mele, M.C. Tomato Lycopene and Lung Cancer Prevention: From Experimental to Human Studies. Cancers 2011, 3, 2333–2357. [Google Scholar] [CrossRef] [PubMed]
- Jacques, P.F.; Lyass, A.; Massaro, J.M.; Vasan, R.S.; D’Agostino Sr, R.B. Relationship of Lycopene Intake and Consumption of Tomato Products to Incident CVD. Br. J. Nutr. 2013, 110, 545–551. [Google Scholar] [CrossRef]
- Lu, S.; Chen, S.; Li, H.; Paengkoum, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; et al. Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review. Animals 2022, 12, 3294. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Modirsanei, M.; Radfar, M.; Kiaei, M.M.; Farkhoy, M.; Honarzad, J. Digestibility and Metabolisable Energy Values of Dried Tomato Pomace for Laying and Meat Type Cockerels. Anim. Feed Sci. Technol. 2008, 141, 384–390. [Google Scholar] [CrossRef]
- Reda, F.M.; Madkour, M.; El-Azeem, N.A.; Aboelazab, O.; Ahmed, S.Y.A.; Alagawany, M. Tomato Pomace as a Nontraditional Feedstuff: Productive and Reproductive Performance, Digestive Enzymes, Blood Metabolites, and the Deposition of Carotenoids into Egg Yolk in Quail Breeders. Poult. Sci. 2022, 101, 101730. [Google Scholar] [CrossRef]
- Marcos, C.N.; de Evan, T.; Molina-Alcaide, E.; Carro, M.D. Nutritive Value of Tomato Pomace for Ruminants and Its Influence on In Vitro Methane Production. Animals 2019, 9, 343. [Google Scholar] [CrossRef]
- Weiss, W.P.; Frobose, D.L.; Koch, M.E. Wet Tomato Pomace Ensiled with Corn Plants for Dairy Cows. J. Dairy Sci. 1997, 80, 2896–2900. [Google Scholar] [CrossRef]
- Leni, G.; Maistrello, L.; Pinotti, G.; Sforza, S.; Caligiani, A. Production of Carotenoid-Rich Hermetia Illucens Larvae Using Specific Agri-Food by-Products. J. Insects Food Feed 2023, 9, 171–181. [Google Scholar] [CrossRef]
- Brai, A.; Poggialini, F.; Trivisani, C.I.; Vagaggini, C.; Tarchi, F.; Francardi, V.; Dreassi, E. Efficient Use of Agricultural Waste to Naturally Fortify Tenebrio Molitor Mealworms and Evaluation of Their Nutraceutical Properties. J. Insects Food Feed 2023, 9, 599–610. [Google Scholar] [CrossRef]
- Naser El Deen, S.; Spranghers, T.; Baldacchino, F.; Deruytter, D. The Effects of the Particle Size of Four Different Feeds on the Larval Growth of Tenebrio molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2022, 119, 242–249. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 18th ed.; AOAC International: Gaitherburg, MD, USA, 2005. [Google Scholar]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio molitor and Alphitobius Diaperinus Larvae on Seed Cleaning Process Byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef] [PubMed]
- Riaz, K.; Iqbal, T.; Khan, S.; Usman, A.; Al-Ghamdi, M.S.; Shami, A.; El Hadi Mohamed, R.A.; Almadiy, A.A.; Al Galil, F.M.A.; Alfuhaid, N.A.; et al. Growth Optimization and Rearing of Mealworm (Tenebrio molitor L.) as a Sustainable Food Source. Foods 2023, 12, 1891. [Google Scholar] [CrossRef] [PubMed]
- Rho, M.S.; Pum Lee, K. Nutrient-Specific Food Selection Buffers the Effect of Nutritional Imbalance in the Mealworm Beetle, Tenebrio molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2015, 112, 251–258. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-llan, D.I.; Tedders, W.L. Use of Nutrient Self-Selection as a Diet Refining Tool in Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- Purschke, B.; Brüggen, H.; Scheibelberger, R.; Jäger, H. Effect of Pre-Treatment and Drying Method on Physico-Chemical Properties and Dry Fractionation Behaviour of Mealworm Larvae (Tenebrio molitor L.). Eur. Food Res. Technol. 2018, 244, 269–280. [Google Scholar] [CrossRef]
- Azzollini, D.; Derossi, A.; Severini, C. Understanding the Drying Kinetic and Hygroscopic Behaviour of Larvae of Yellow Mealworm (Tenebrio Molitor) and the Effects on Their Quality. J. Insects Food Feed 2016, 2, 233–243. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.; Benning, R.; Haase, H. Effect of Different Drying Methods on Nutrient Quality of the Yellow Mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef]
- Verhaeghe, T.; Vlaemynck, G.; De Block, J.; Van Weyenberg, S.; Hendrickx, M. Thermal Inactivation Kinetics of Proteases and Polyphenoloxidase in Brown Shrimp (Crangon Crangon). Food Chem. 2016, 197, 641–647. [Google Scholar] [CrossRef]
- Cacchiarelli, C.; Fratini, F.; Puccini, M.; Vitolo, S.; Paci, G.; Mancini, S. Effects of Different Blanching Treatments on Colour and Microbiological Profile of Tenebrio molitor and Zophobas Morio Larvae. LWT 2022, 157, 113112. [Google Scholar] [CrossRef]
- Waldbauer, G.B. The Consumption and Utilization of Food by Insects. Adv. Insect Physiol. 1968, 5, 229–288. [Google Scholar]
- Anthon, G.; Barrett, D.M. Standardization Of A Rapid Spectrophotometric Method For Lycopene Analysis. Acta Hortic. 2007, 758, 111–128. [Google Scholar] [CrossRef]
- Gkinali, A.-A.; Matsakidou, A.; Paraskevopoulou, A. Characterization of Tenebrio molitor Larvae Protein Preparations Obtained by Different Extraction Approaches. Foods 2022, 11, 3852. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.-H.; Son, Y.-J.; Kim, S.-H.; Yun, E.-Y.; Kang, H.-J.; Hwang, I.-K. Physicochemical Properties and Oxidative Stabilities of Mealworm (Tenebrio molitor) Oils under Different Roasting Conditions. Food Sci. Biotechnol. 2016, 25, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Laroche, M.; Perreault, V.; Marciniak, A.; Gravel, A.; Chamberland, J.; Doyen, A. Comparison of Conventional and Sustainable Lipid Extraction Methods for the Production of Oil and Protein Isolate from Edible Insect Meal. Foods 2019, 8, 572. [Google Scholar] [CrossRef] [PubMed]
- Tasselli, G.; Filippucci, S.; Borsella, E.; D’Antonio, S.; Gelosia, M.; Cavalaglio, G.; Turchetti, B.; Sannino, C.; Onofri, A.; Mastrolitti, S.; et al. Yeast Lipids from Cardoon Stalks, Stranded Driftwood and Olive Tree Pruning Residues as Possible Extra Sources of Oils for Producing Biofuels and Biochemicals. Biotechnol. Biofuels 2018, 11, 147. [Google Scholar] [CrossRef]
- Di Fidio, N.; Liuzzi, F.; Mastrolitti, S.; Albergo, R.; De Bari, I. Single Cell Oil Production from Undetoxified Arundo donax L. Hydrolysate by Cutaneotrichosporon Curvatus. J. Microbiol. Biotechnol. 2019, 29, 256–267. [Google Scholar] [CrossRef]
- Lawrence, G.D. Perspective: The Saturated Fat–Unsaturated Oil Dilemma: Relations of Dietary Fatty Acids and Serum Cholesterol, Atherosclerosis, Inflammation, Cancer, and All-Cause Mortality. Adv. Nutr. 2021, 12, 647–656. [Google Scholar] [CrossRef]
- Massaro, M.; Scoditti, E.; Carluccio, M.A.; Montinari, M.R.; De Caterina, R. Omega–3 Fatty Acids, Inflammation and Angiogenesis: Nutrigenomic Effects as an Explanation for Anti-Atherogenic and Anti-Inflammatory Effects of Fish and Fish Oils. LifestyleGenom. 2008, 1, 4–23. [Google Scholar] [CrossRef]
- Khalili Tilami, S.; Kouřimská, L. Assessment of the Nutritional Quality of Plant Lipids Using Atherogenicity and Thrombogenicity Indices. Nutrients 2022, 14, 3795. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional Indices for Assessing Fatty Acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, J.; Bessa, R.J.; Santos-Silva, F. Effect of Genotype, Feeding System and Slaughter Weight on the Quality of Light Lambs. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Cazzola, R.; Rondanelli, M.; Russo-Volpe, S.; Ferrari, E.; Cestaro, B. Decreased Membrane Fluidity and Altered Susceptibility to Peroxidation and Lipid Composition in Overweight and Obese Female Erythrocytes. J. Lipid Res. 2004, 45, 1846–1851. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, M.S.; Yamasaki, H.; Yazawa, K.; Masumura, S. Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Plasma Membrane Fluidity of Aortic Endothelial Cells. Lipids 1999, 34, 1297–1304. [Google Scholar] [CrossRef]
- Abril, D.; Mirabal-Gallardo, Y.; González, A.; Marican, A.; Durán-Lara, E.F.; Silva Santos, L.; Valdés, O. Comparison of the Oxidative Stability and Antioxidant Activity of Extra-Virgin Olive Oil and Oils Extracted from Seeds of Colliguaya Integerrima and Cynara Cardunculus Under Normal Conditions and After Thermal Treatment. Antioxidants 2019, 8, 470. [Google Scholar] [CrossRef]
- Fatemi, S.H.; Hammond, E.G. Analysis of Oleate, Linoleate and Linolenate Hydroperoxides in Oxidized Ester Mixtures. Lipids 1980, 15, 379–385. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow Mealworm Protein for Food Purposes-Extraction and Functional Properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef]
- Janssen, R.H.; Lakemond, C.M.M.; Fogliano, V.; Renzone, G.; Scaloni, A.; Vincken, J.-P. Involvement of Phenoloxidase in Browning during Grinding of Tenebrio molitor Larvae. PLoS ONE 2017, 12, e0189685. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, K.P. Geometric Analysis of Nutrient Balancing in the Mealworm Beetle, Tenebrio molitor L. (Coleoptera: Tenebrionidae). J. Insect Physiol. 2014, 71, 37–45. [Google Scholar] [CrossRef]
- Rho, M.S.; Lee, K.P. Balanced Intake of Protein and Carbohydrate Maximizes Lifetime Reproductive Success in the Mealworm Beetle, Tenebrio molitor (Coleoptera: Tenebrionidae). J. Insect Physiol. 2016, 91–92, 93–99. [Google Scholar] [CrossRef]
- Ruschioni, S.; Loreto, N.; Foligni, R.; Mannozzi, C.; Raffaelli, N.; Zamporlini, F.; Pasquini, M.; Roncolini, A.; Cardinali, F.; Osimani, A.; et al. Addition of Olive Pomace to Feeding Substrate Affects Growth Performance and Nutritional Value of Mealworm (Tenebrio molitor L.) Larvae. Foods 2020, 9, 317. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stasiak, M.; Li, L.; Xie, B.; Fu, Y.; Gidzinski, D.; Dixon, M.; Liu, H. Rearing Tenebrio molitor in BLSS: Dietary Fiber Affects Larval Growth, Development, and Respiration Characteristics. Acta Astronaut. 2016, 118, 130–136. [Google Scholar] [CrossRef]
- Morales Ramos, J.; Rojas, M.-G.; Shapiro Ilan, D.; Tedders, W. Developmental Plasticity in Tenebrio molitor (Coleoptera: Tenebrionidae): Analysis of Instar Variation in Number and Development Time under Different Diets. J. Entomol. Sci. 2010, 45, 75–90. [Google Scholar] [CrossRef]
- van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth Performance and Feed Conversion Efficiency of Three Edible Mealworm Species (Coleoptera: Tenebrionidae) on Diets Composed of Organic by-Products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Kelstrup, H.C.; Emery, V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects 2020, 11, 827. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G. Effect of Larval Density on Food Utilization Efficiency of Tenebrio Molitor (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2015, 108, 2259–2267. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, S.; Paci, G.; Fratini, F.; Dal Bosco, A.; Tuccinardi, T.; Mancini, S. Former Foodstuff in Mealworm Farming: Effects on Fatty Acids Profile, Lipid Metabolism and Antioxidant Molecules. LWT 2021, 147, 111644. [Google Scholar] [CrossRef]
- Finke, M.D. Complete Nutrient Content of Four Species of Commercially Available Feeder Insects Fed Enhanced Diets during Growth. Zoo Biol. 2015, 34, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Enhancing the Nutritional Profile of Tenebrio molitor Using the Leaves of Moringa Oleifera. Foods 2023, 12, 2612. [Google Scholar] [CrossRef]
- Rovai, D.; Ortgies, M.; Amin, S.; Kuwahara, S.; Schwartz, G.; Lesniauskas, R.; Garza, J.; Lammert, A. Utilization of Carrot Pomace to Grow Mealworm Larvae (Tenebrio molitor). Sustainability 2021, 13, 9341. [Google Scholar] [CrossRef]
- Cobbs, C.; Heath, J.; Stireman, J.O.; Abbot, P. Carotenoids in Unexpected Places: Gall Midges, Lateral Gene Transfer, and Carotenoid Biosynthesis in Animals. Mol. Phylogenet. Evol. 2013, 68, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Mein, J.R.; Dolnikowski, G.G.; Ernst, H.; Russell, R.M.; Wang, X.-D. Enzymatic Formation of Apo-Carotenoids from the Xanthophyll Carotenoids Lutein, Zeaxanthin and β-Cryptoxanthin by Ferret Carotene-9′,10′-Monooxygenase. Arch. Biochem. Biophys. 2011, 506, 109–121. [Google Scholar] [CrossRef]
- Poveda, J. Insect Frass in the Development of Sustainable Agriculture. A Review. Agron. Sustain. Dev. 2021, 41, 5. [Google Scholar] [CrossRef]
- Errico, S.; Spagnoletta, A.; Verardi, A.; Moliterni, S.; Dimatteo, S.; Sangiorgio, P. Tenebrio Molitor as a Source of Interesting Natural Compounds, Their Recovery Processes, Biological Effects, and Safety Aspects. Compr. Rev. Food Sci. Food Saf. 2022, 21, 148–197. [Google Scholar] [CrossRef] [PubMed]
- Ravzanaadii, N.; Kim, S.-H.; Choi, W.-H.; Hong, S.-J.; Kim, N.-J. Nutritional Value of Mealworm, Tenebrio molitor as Food Source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect Meal as Renewable Source of Food for Animal Feeding: A Review. J. Clean. Prod. 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Zheng, T.; Li, H.; Han, N.; Wang, S.; Hackney Price, J.; Wang, M.; Zhang, D. Functional Characterization of Two Elongases of Very Long-Chain Fatty Acid from Tenebrio molitor L. (Coleoptera: Tenebrionidae). Sci. Rep. 2017, 7, 10990. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; Finke, M.D. Nutritional Value of Insects and Ways to Manipulate Their Composition. J. Insects Food Feed 2021, 7, 639–659. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schlüter, O.K. Nutritional Composition and Safety Aspects of Edible Insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO: Rome, Italy, 2013; No. 171. [Google Scholar]
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.-H. Role of Temperature on Growth and Metabolic Rate in the Tenebrionid Beetles Alphitobius Diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.-L.; Lognay, G.; Francis, F.; Caparros Megido, R. About Lipid Metabolism in Hermetia illucens (L. 1758): On the Origin of Fatty Acids in Prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef]
- van Dooremalen, C.; Ellers, J. A Moderate Change in Temperature Induces Changes in Fatty Acid Composition of Storage and Membrane Lipids in a Soil Arthropod. J. Insect Physiol. 2010, 56, 178–184. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, B.; Struți, D.; Păpuc, T.; Cighi, V.; Boaru, A. Effect of the Energy Content of Diets on the Development and Quality of the Fat Reserves of Larvae and Reproduction of Adults of the Black Soldier Fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2021, 118, 297–306. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, Z.; Ma, Y.; Qu, Y.; Lu, X.; Guo, H.; Luo, H. Effect of Dietary Lycopene Supplementation on Growth Performance, Meat Quality, Fatty Acid Profile and Meat Lipid Oxidation in Lambs in Summer Conditions. Small Rumin. Res. 2015, 131, 99–106. [Google Scholar] [CrossRef]
- Hernández-López, S.H.; Rodríguez-Carpena, J.G.; Lemus-Flores, C.; Grageola-Nuñez, F.; Estévez, M. Avocado Waste for Finishing Pigs: Impact on Muscle Composition and Oxidative Stability during Chilled Storage. Meat Sci. 2016, 116, 186–192. [Google Scholar] [CrossRef]
- Biondi, L.; Luciano, G.; Cutello, D.; Natalello, A.; Mattioli, S.; Priolo, A.; Lanza, M.; Morbidini, L.; Gallo, A.; Valenti, B. Meat Quality from Pigs Fed Tomato Processing Waste. Meat Sci. 2020, 159, 107940. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio molitor Larvae as an Alternative Food Source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Mercola, J.; D’Adamo, C.R. Linoleic Acid: A Narrative Review of the Effects of Increased Intake in the Standard American Diet and Associations with Chronic Disease. Nutrients 2023, 15, 3129. [Google Scholar] [CrossRef]
- Fasel, N.J.; Mène-Saffrané, L.; Ruczyński, I.; Komar, E.; Christe, P. Diet Induced Modifications of Fatty-Acid Composition in Mealworm Larvae (Tenebrio molitor). J. Food Res. 2017, 6, 22–31. [Google Scholar] [CrossRef]
- Brandstetter, B.; Ruther, J. An Insect with a Delta-12 Desaturase, the Jewel Wasp Nasonia Vitripennis, Benefits from Nutritional Supply with Linoleic Acid. Sci. Nat. 2016, 103, 40. [Google Scholar] [CrossRef]
- Amoedo, N.D.; Punzi, G.; Obre, E.; Lacombe, D.; De Grassi, A.; Pierri, C.L.; Rossignol, R. AGC1/2, the Mitochondrial Aspartate-Glutamate Carriers. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2394–2412. [Google Scholar] [CrossRef] [PubMed]
- Romero-Lorente, M.Á.; Fabrikov, D.; Montes, J.; Morote, E.; Barroso, F.G.; Vargas-García, M.D.C.; Varga, Á.T.; Sánchez-Muros, M.J. Pre-Treatment of Fish By-Products to Optimize Feeding of Tenebrio molitor L. Larvae. Insects 2022, 13, 125. [Google Scholar] [CrossRef] [PubMed]
- Lawal, K.G.; Kavle, R.R.; Akanbi, T.O.; Mirosa, M.; Agyei, D. Enrichment in Specific Fatty Acids Profile of Tenebrio molitor and Hermetia illucens Larvae through Feeding. Futur. Foods 2021, 3, 100016. [Google Scholar] [CrossRef]
- Otero, P.; Gutierrez-Docio, A.; Navarro del Hierro, J.; Reglero, G.; Martin, D. Extracts from the Edible Insects Acheta Domesticus and Tenebrio molitor with Improved Fatty Acid Profile Due to Ultrasound Assisted or Pressurized Liquid Extraction. Food Chem. 2020, 314, 126200. [Google Scholar] [CrossRef] [PubMed]
- Oppedisano, F.; Mollace, R.; Tavernese, A.; Gliozzi, M.; Musolino, V.; Macrì, R.; Carresi, C.; Maiuolo, J.; Serra, M.; Cardamone, A.; et al. PUFA Supplementation and Heart Failure: Effects on Fibrosis and Cardiac Remodeling. Nutrients 2021, 13, 2965. [Google Scholar] [CrossRef] [PubMed]
- Fedor, D.; Kelley, D.S. Prevention of Insulin Resistance by N-3 Polyunsaturated Fatty Acids. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Oppedisano, F.; Bulotta, R.M.; Maiuolo, J.; Gliozzi, M.; Musolino, V.; Carresi, C.; Ilari, S.; Serra, M.; Muscoli, C.; Gratteri, S.; et al. The Role of Nutraceuticals in Osteoarthritis Prevention and Treatment: Focus on n-3 PUFAs. Oxid. Med. Cell. Longev. 2021, 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Role of Soluble Epoxide Hydrolase in Metabolism of PUFAs in Psychiatric and Neurological Disorders. Front. Pharmacol. 2019, 10, 36. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Health-Promoting Ingredients from Four Selected Azorean Macroalgae. Food Res. Int. 2016, 89, 432–438. [Google Scholar] [CrossRef]
- Ahmad, N.; Shabbir, U.; Sameen, A.; Manzoor, M.F.; Ahmad, M.H.; Ismail, T.; Ahmed, S.; Siddique, R. Hypocholesterolemic Effect of Designer Yogurts Fortified with Omega Fatty Acids and Dietary Fibers in Hypercholesterolemic Subjects. Food Sci. Technol. 2021, 41, 1000–1008. [Google Scholar] [CrossRef]
- Pekkoh, J.; Phinyo, K.; Thurakit, T.; Lomakool, S.; Duangjan, K.; Ruangrit, K.; Pumas, C.; Jiranusornkul, S.; Yooin, W.; Cheirsilp, B.; et al. Lipid Profile, Antioxidant and Antihypertensive Activity, and Computational Molecular Docking of Diatom Fatty Acids as ACE Inhibitors. Antioxidants 2022, 11, 186. [Google Scholar] [CrossRef]
- Moussavi Javardi, M.S.; Madani, Z.; Movahedi, A.; Karandish, M.; Abbasi, B. The Correlation between Dietary Fat Quality Indices and Lipid Profile with Atherogenic Index of Plasma in Obese and Non-Obese Volunteers: A Cross-Sectional Descriptive-Analytic Case-Control Study. Lipids Health Dis. 2020, 19, 213. [Google Scholar] [CrossRef]
- Gantner, M.; Król, K.; Piotrowska, A.; Sionek, B.; Sadowska, A.; Kulik, K.; Wiącek, M. Adding Mealworm (Tenebrio molitor L.) Powder to Wheat Bread: Effects on Physicochemical, Sensory and Microbiological Qualities of the End-Product. Molecules 2022, 27, 6155. [Google Scholar] [CrossRef]
- Zielińska, E.; Pankiewicz, U. Nutritional, Physiochemical, and Antioxidative Characteristics of Shortcake Biscuits Enriched with Tenebrio molitor Flour. Molecules 2020, 25, 5629. [Google Scholar] [CrossRef]
- Djouadi, A.; Sales, J.R.; Carvalho, M.O.; Raymundo, A. Development of Healthy Protein-Rich Crackers Using Tenebrio molitor Flour. Foods 2022, 11, 702. [Google Scholar] [CrossRef]
- Sharifinia, M.; Bahmanbeigloo, Z.A.; Keshavarzifard, M.; Khanjani, M.H.; Daliri, M.; Koochaknejad, E.; Jasour, M.S. Fishmeal Replacement by Mealworm (Tenebrio Molitor) in Diet of Farmed Pacific White Shrimp (Litopenaeus Vannamei): Effects on Growth Performance, Serum Biochemistry, and Immune Response. Aquat. Living Resour. 2023, 36, 19. [Google Scholar] [CrossRef]
- Biasato, I.; Gasco, L.; De Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Sterpone, L.; et al. Yellow Mealworm Larvae (Tenebrio molitor) Inclusion in Diets for Male Broiler Chickens: Effects on Growth Performance, Gut Morphology, and Histological Findings. Poult. Sci. 2018, 97, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Ao, X.; Kim, I.H. Effects of Dietary Dried Mealworm (Ptecticus Tenebrifer) Larvae on Growth Performance and Nutrient Digestibility in Weaning Pigs. Livest. Sci. 2019, 230, 103815. [Google Scholar] [CrossRef]
- Son, Y.-J.; Choi, S.Y.; Hwang, I.-K.; Nho, C.W.; Kim, S.H. Could Defatted Mealworm (Tenebrio molitor) and Mealworm Oil Be Used as Food Ingredients? Foods 2020, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Naccarato, A.; Vommaro, M.L.; Amico, D.; Sprovieri, F.; Pirrone, N.; Tagarelli, A.; Giglio, A. Triazine Herbicide and NPK Fertilizer Exposure: Accumulation of Heavy Metals and Rare Earth Elements, Effects on Cuticle Melanization, and Immunocompetence in the Model Species Tenebrio molitor. Toxics 2023, 11, 499. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| By-Product | Dry Matter (%) | Crude Protein (%) | Crude Fat (%) | Crude Fiber (%) | Ash (%) | Carbohydrate (%) |
|---|---|---|---|---|---|---|
| Bran | 91.2 | 16.7 | 6.5 | 36.1 | 4.2 | 30.2 |
| Tomato pomace | 92.1 | 9.5 | 3.2 | 67.1 | 3.9 | 8.9 |
| Brewer’s spent grain | 93.4 | 24.7 | 4.8 | 42.0 | 2.6 | 24.0 |
| Yeast | 93.0 | 47.6 | 2.4 | 6.8 | 8.0 | 13.8 |
| Diet | Bran (%) | Tomato Pomace (%) | Brewer’s Spent Grain (%) | Yeast (%) | Protein Value (%) | Carbohydrate (%) | p:c 4 | Crude Fiber (%) | Fat (%) | Energy (kcal (100 g) |
|---|---|---|---|---|---|---|---|---|---|---|
| TP0 2 | 100.0 | - | - | - | 16.7 | 30.2 | 1:1.8 | 36.1 | 6.5 | 318.3 |
| TP27 3 | 50.0 | 27.0 | 23.0 | - | 16.6 | 23.0 | 1:1.4 | 45.8 | 5.2 | 296.9 |
| TP41 3 | 50.0 | 41.0 | - | 9.0 | 16.5 | 20.0 | 1:1.2 | 46.2 | 4.8 | 281.4 |
| TP100 | - | 100.0 | - | - | 9.5 | 8.9 | 1:0.9 | 67.1 | 3.2 | 236.6 |
| Diet | FC (mg Larvae 1) | FCR | SRG (% day−1) | ECI (%) | ECD (%) |
|---|---|---|---|---|---|
| TP0 2 | 181.1 ± 44.7 c | 2.7 ± 0.2 d | 4.9 ± 0.7 a | 15.4 ± 1.1 a | 34.6 ± 3.3 c |
| TP27 | 222.5 ± 29.6 bc | 3.2 ± 0.1 c | 4.1 ± 0.4 b | 13.1 ± 0.5 b | 42.8 ± 4.5 b |
| TP41 | 291.1 ± 38.5 a | 3.8 ± 0.3 b | 4.3 ± 0.4 b | 10.8 ± 0.7 c | 30.0 ± 6.3 c |
| TP100 | 245.7 ± 80.3 ab | 4.3 ± 0.4 a | 2.5 ± 0.7 c | 9.8 ± 1.0 d | 65.9 ± 12.7 a |
| Diet | Feed | Mealworm | Frass | |||
|---|---|---|---|---|---|---|
| Lycopene (ug/g) | β-Carotene (ug/g) | Lycopene (ug/g) | β-Carotene (ug/g) | Lycopene (ug/g) | β-Carotene (ug/g) | |
| TP0 2 | 2.7 ± 0.2 d | 0.3 ± 0.1 d | 0.1 ± 0.1 c | 1.4 ± 1.0 bc | 0.7 ± 0.0 d | 12.1 ± 0.3 d |
| TP27 | 22.7 ± 0.8 c | 45.3 ± 2.0 c | 0.1 ± 0.0 c | 1.1 ± 0.2 c | 12.4 ± 0.4 c | 51.0 ± 0.6 c |
| TP41 | 52.4 ± 1.7 b | 95.1 ± 0.7 b | 0.6 ± 0.3 b | 2.6 ± 0.8 b | 24.1 ± 0.3 b | 76.3 ± 1.2 b |
| TP100 | 179.8 ± 2.7 a | 241.5 ± 2.5 a | 1.2 ± 0.3 a | 7.3 ± 0.1 a | 39.7 ± 1.6 a | 147.5 ± 4.6 a |
| Fatty Acid (%) | Diets | ||||
|---|---|---|---|---|---|
| Common Name | Lipid Number | TP0 2 | TP27 | TP41 | TP100 |
| Caprilic acid | C8:0 | n.d. | n.d. | n.d. | n.d. |
| Capric acid | C10:0 | n.d. | n.d. | n.d. | n.d. |
| Lauric acid | C12:0 | n.d. | n.d. | n.d. | n.d. |
| Myristic acid | C14:0 | 3.7 ± 0.0 a | 3.8 ± 0.0 a | 3.4 ± 0.1 b | 2.7 ± 0.1 c |
| Palmitic acid | C16:0 | 15.6 ± 0.0 a | 14.8 ± 0.1 ab | 15.3 ± 0.8 a | 13.8 ± 0.1 c |
| Palmitoleic acid | C16:1 | 1.6 ± 0.2 b | 4.2 ± 0.0 a | 1.4 ± 0.2 b | 1.0 ± 0.0 c |
| Stearic acid | C18:0 | 2.9 ± 0.1 c | 3.2 ± 0.1 bc | 3.4 ± 0.2 b | 5.6 ± 0.1 a |
| Oleic acid | C18:1 | 50.2 ± 0.2 a | 44.7 ± 0.3 b | 42.9 ± 0.2 b | 26.2 ± 1.5 c |
| α-Linoleic acid | C18:2n-6 | 25.7 ± 0.3 d | 28.9 ± 0.3 c | 32.6 ± 1.0 b | 48.1 ± 1.0 a |
| α-Linolenic acid | C18:3n3 | 0.4 ± 0.0 c | 0.5 ± 0.0 c | 1.0 ± 0.1 b | 2.7 ± 0.1 a |
| Arachidic acid | C20:0 | n.d. | n.d. | n.d. | n.d. |
| Behenic acid | C22:0 | n.d. | n.d. | n.d. | n.d. |
| Erucic acid | C22:1 | n.d. | n.d. | n.d. | n.d. |
| Lignoceric acid | C24:0 | n.d. | n.d. | n.d. | n.d. |
| Σ SFA | 22.1 ± 0.2 a | 21.8 ± 0.0 a | 22.1 ± 0.6 a | 22.0 ± 0.1 a | |
| Σ MUFA | 51.7 ± 0.1 a | 48.8 ± 0.3 b | 44.3 ± 0.5 c | 27.2 ± 1.0 d | |
| Σ PUFA | 26.1 ± 0.2 d | 29.4 ± 0.2 c | 33.6 ± 1.0 b | 50.8 ± 1.0 a | |
| Σ UFA | 77.9 ± 0.2 a | 78.2 ± 0.1 a | 77.9 ± 0.6 a | 78.0 ± 0.1 a | |
| PUFA:SFA ratio | 1.2 ± 0.0 d | 1.4 ± 0.0 c | 1.5 ± 0.1 b | 2.3 ± 0.0 a | |
| MUFA:PUFA ratio | 2.0 ± 0.0 a | 1.7 ± 0.0 b | 1.3 ± 0.0 c | 0.5 ± 0.0 d | |
| ω6:ω3 ratio | 64.3 ± 3.9 a | 57.8 ± 7.3 a | 32.6 ± 1.7 b | 17.8 ± 0.1 c | |
| Index 1 | Diets | |||
|---|---|---|---|---|
| TP0 2 | TP27 | TP41 | TP100 | |
| Cox Index | 3.2 ± 0.1 d | 3.5 ± 0.0 c | 4.0 ± 0.1 b | 5.8 ± 0.1 a |
| IT | 0.6 ± 0.0 a | 0.5 ± 0.0 a | 0.5 ± 0.0 a | 0.5 ± 0.0 a |
| IA | 0.4 ± 0.0 a | 0.4 ± 0.0 a | 0.4 ± 0.0 a | 0.3 ± 0.0 b |
| HH | 4.0 ± 0.0 b | 4.0 ± 0.0 b | 4.1 ± 0.2 b | 4.7 ± 0.0 a |
| UI | 104.4 ± 0.5 d | 108.2 ± 0.1 c | 112.5 ± 1.2 b | 131.4 ± 1.0 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldacchino, F.; Spagnoletta, A.; Lamaj, F.; Vitale, M.L.; Verrastro, V. First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects 2023, 14, 854. https://doi.org/10.3390/insects14110854
Baldacchino F, Spagnoletta A, Lamaj F, Vitale ML, Verrastro V. First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects. 2023; 14(11):854. https://doi.org/10.3390/insects14110854
Chicago/Turabian StyleBaldacchino, Ferdinando, Anna Spagnoletta, Flutura Lamaj, Maria Luisa Vitale, and Vincenzo Verrastro. 2023. "First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae)" Insects 14, no. 11: 854. https://doi.org/10.3390/insects14110854
APA StyleBaldacchino, F., Spagnoletta, A., Lamaj, F., Vitale, M. L., & Verrastro, V. (2023). First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects, 14(11), 854. https://doi.org/10.3390/insects14110854