
Review of the Narrow-Banded Hawkmoth, Neogurelca montana (Rothschild & Jordan, 1915) (Lepidoptera: Sphingidae) in China, with Morphological and Phylogenetic Analysis



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
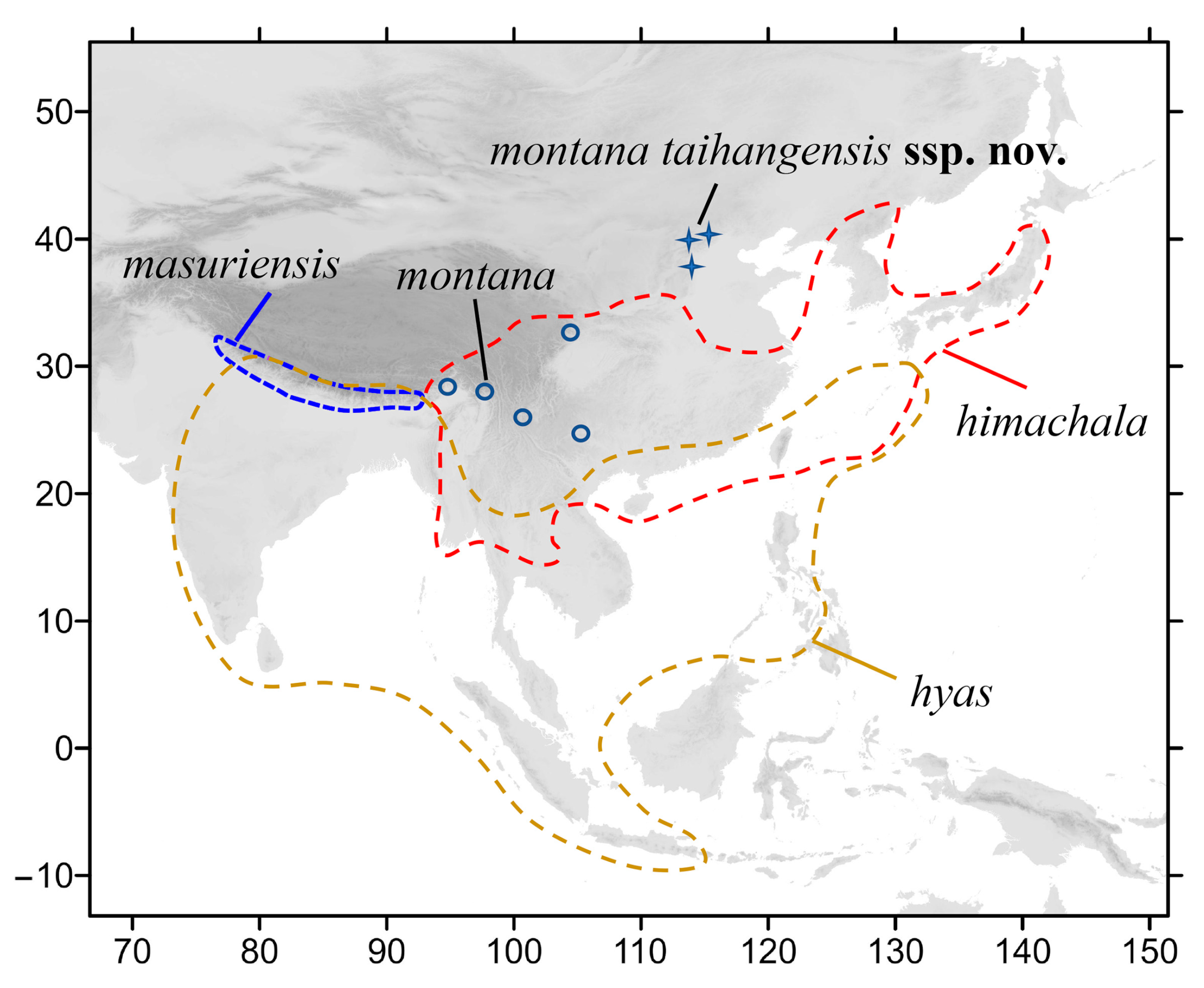
2.1. Taxon Sampling
2.2. DNA Extraction and Amplification
2.3. Phylogenetic Analysis and Species Delimitation
2.4. Morphological Comparison
2.5. Larvae Collection and Rearing
3. Results
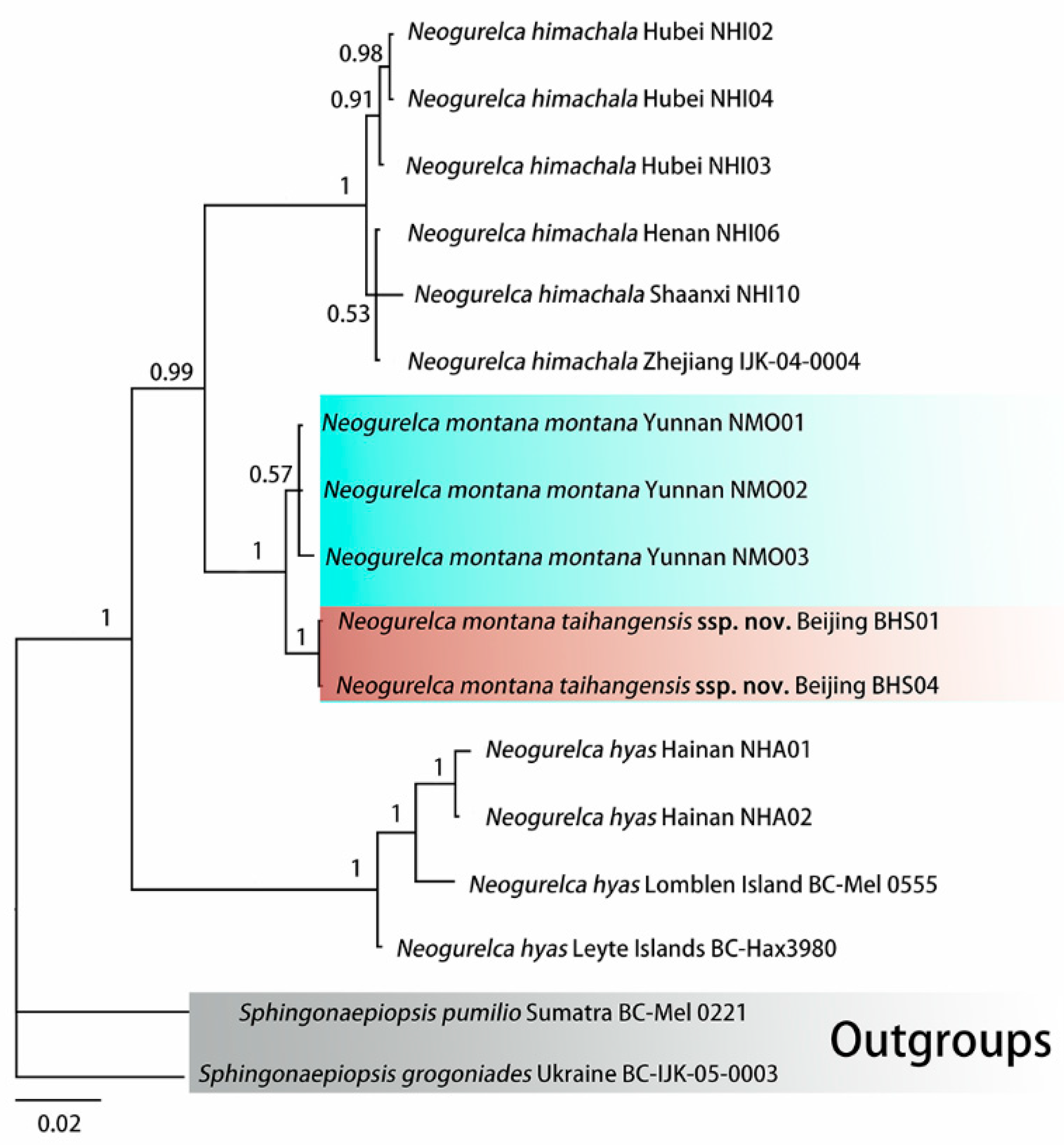
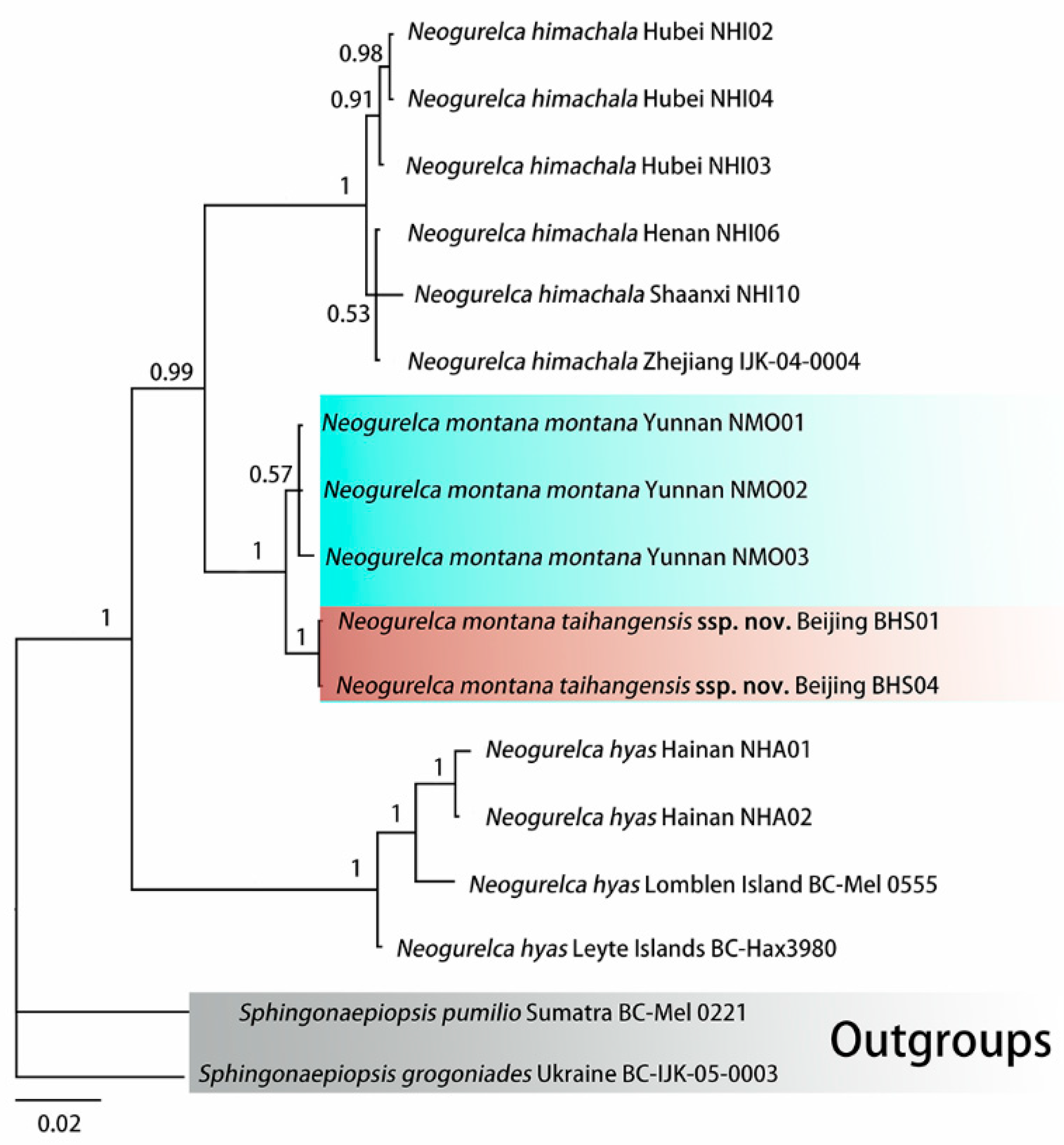
3.1. Molecular Phylogenetic Relationships and Delimitation
3.1.1. Molecular Phylogenetic Analysis
3.1.2. Molecular Species Delimitation
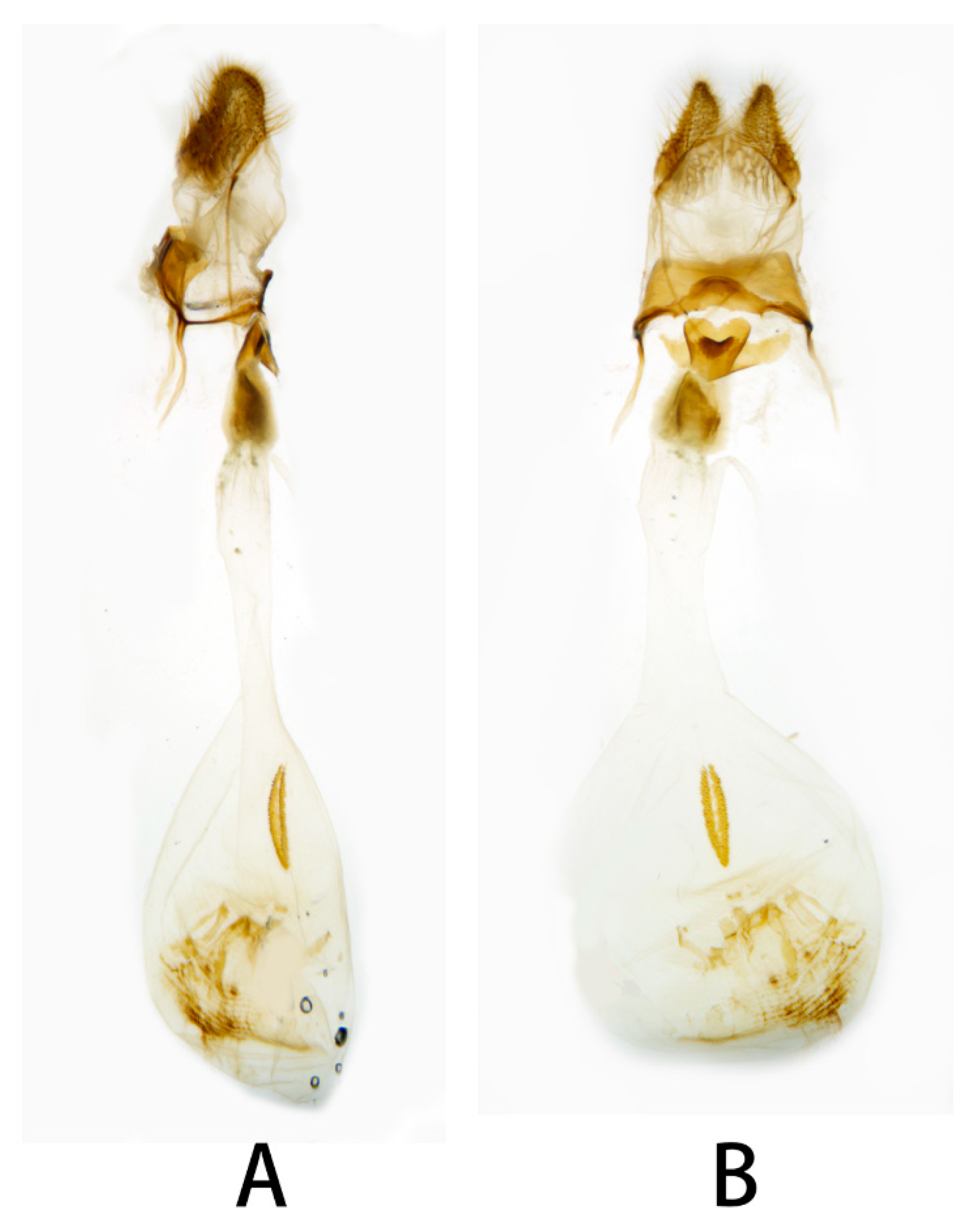
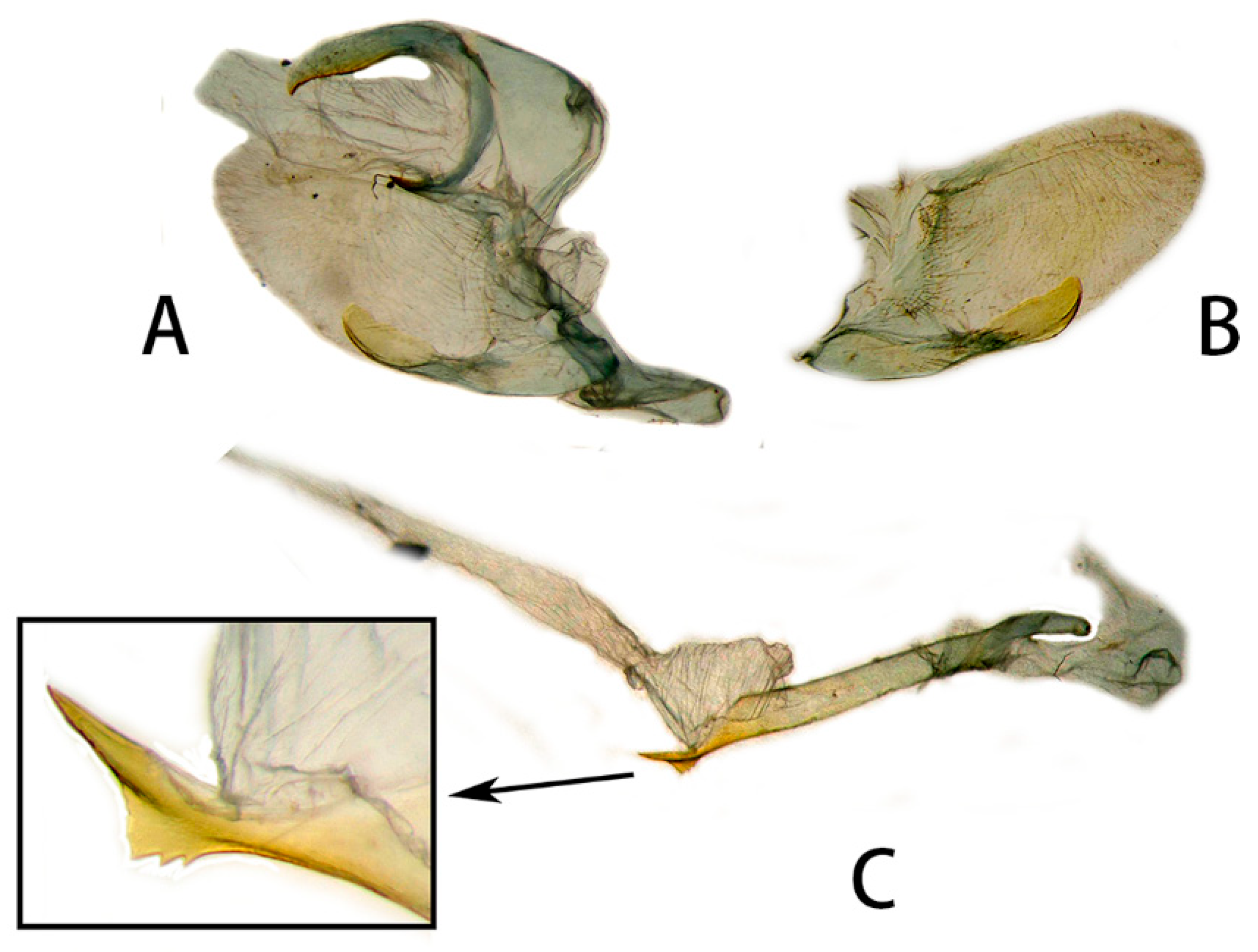
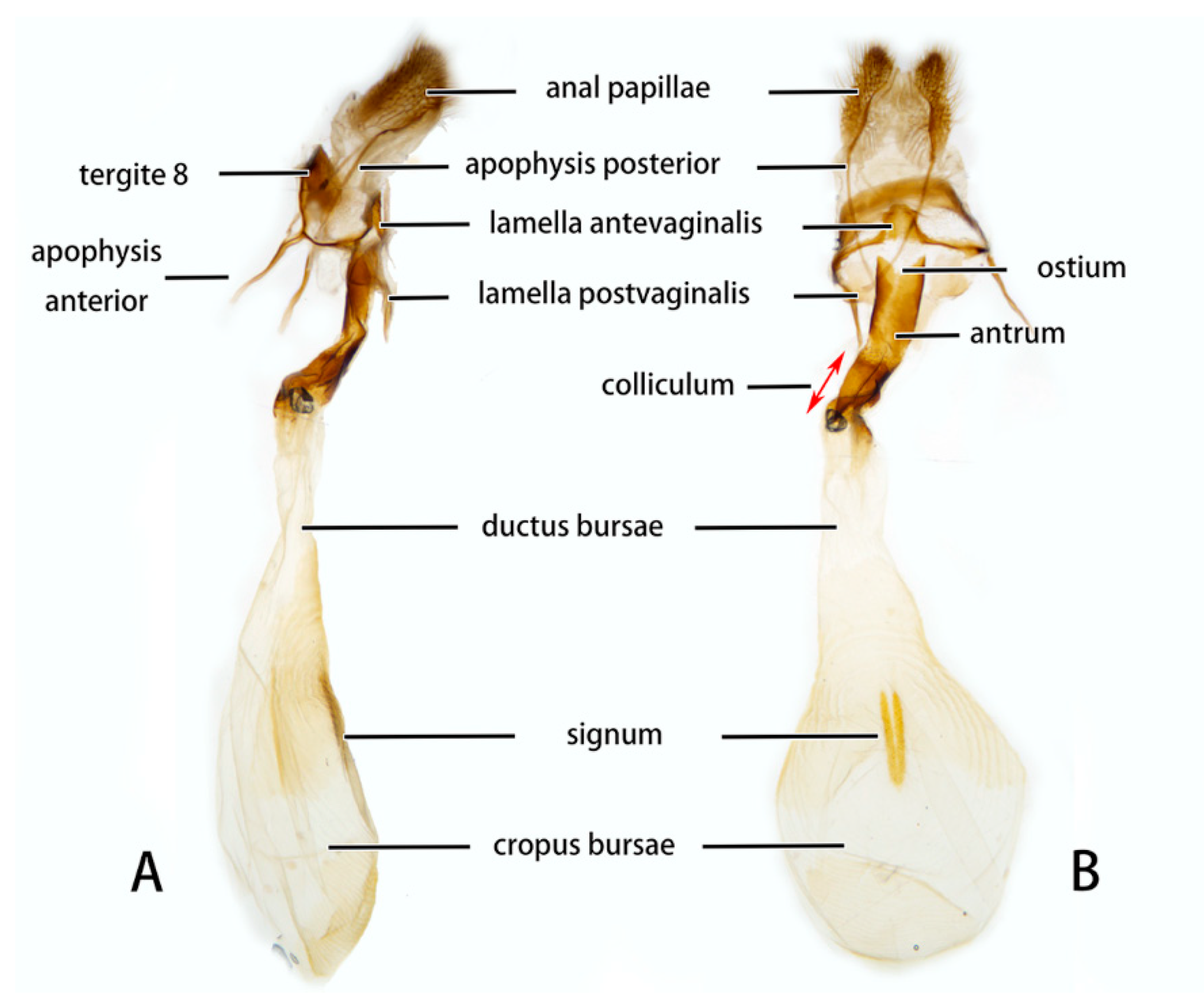
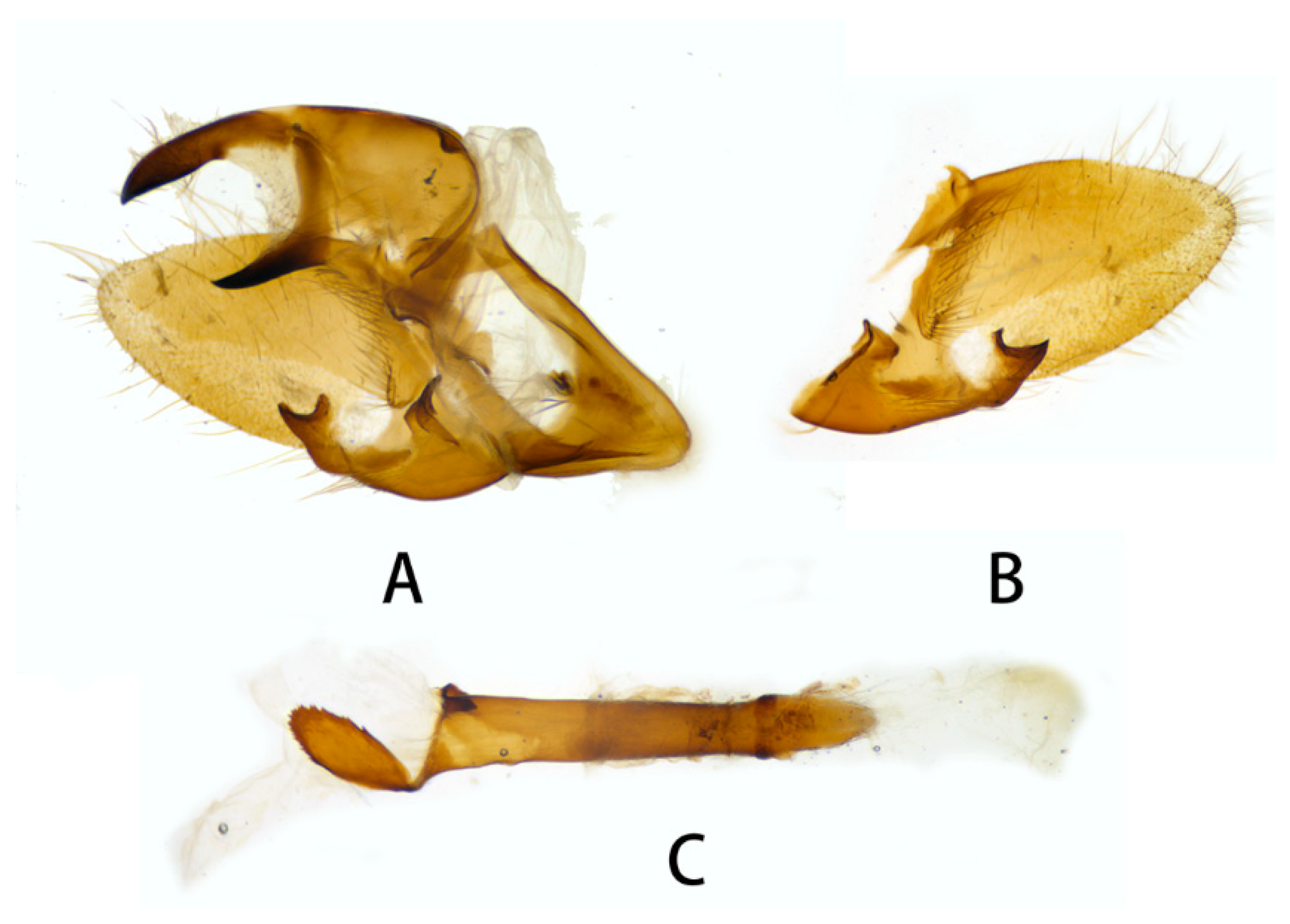
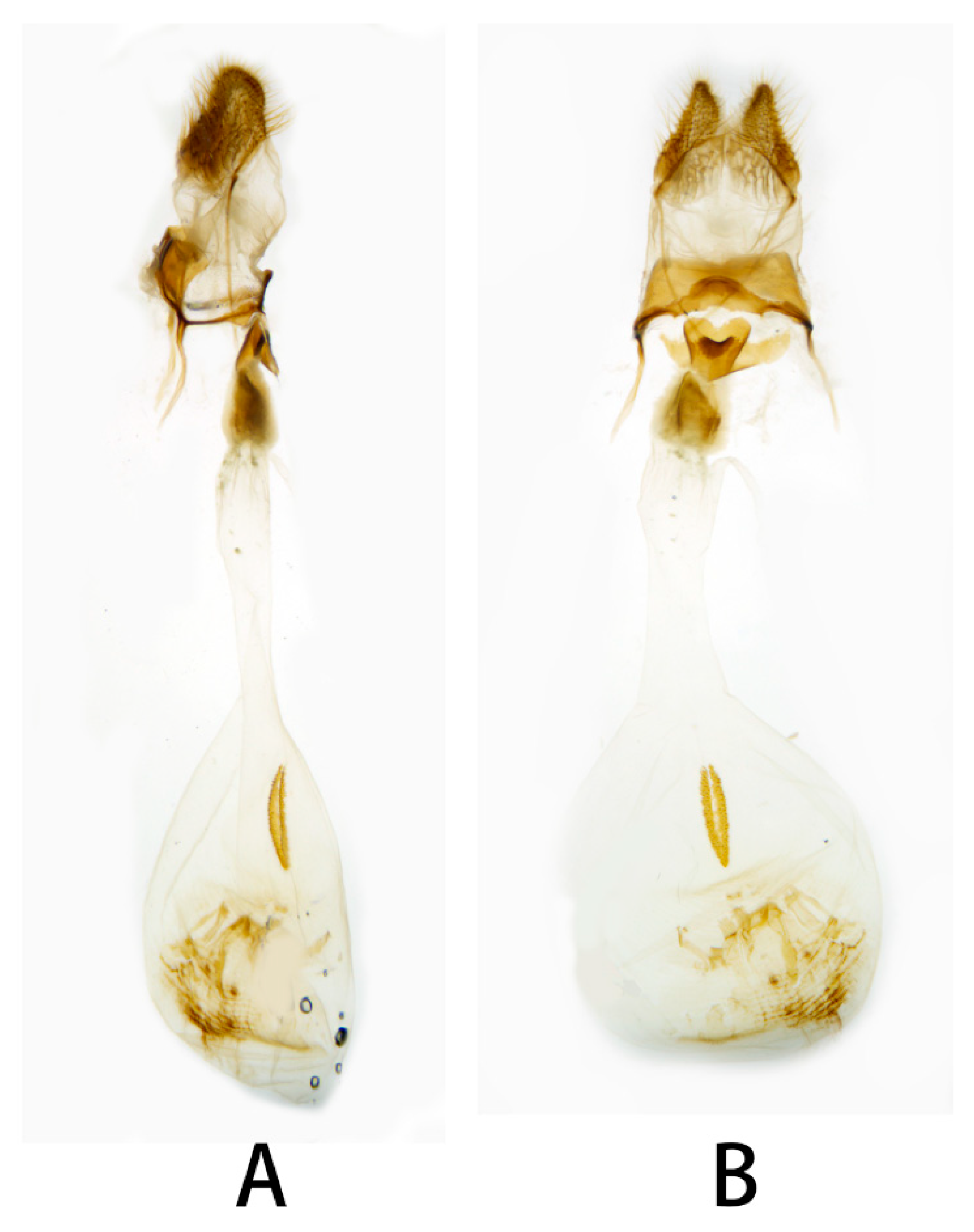
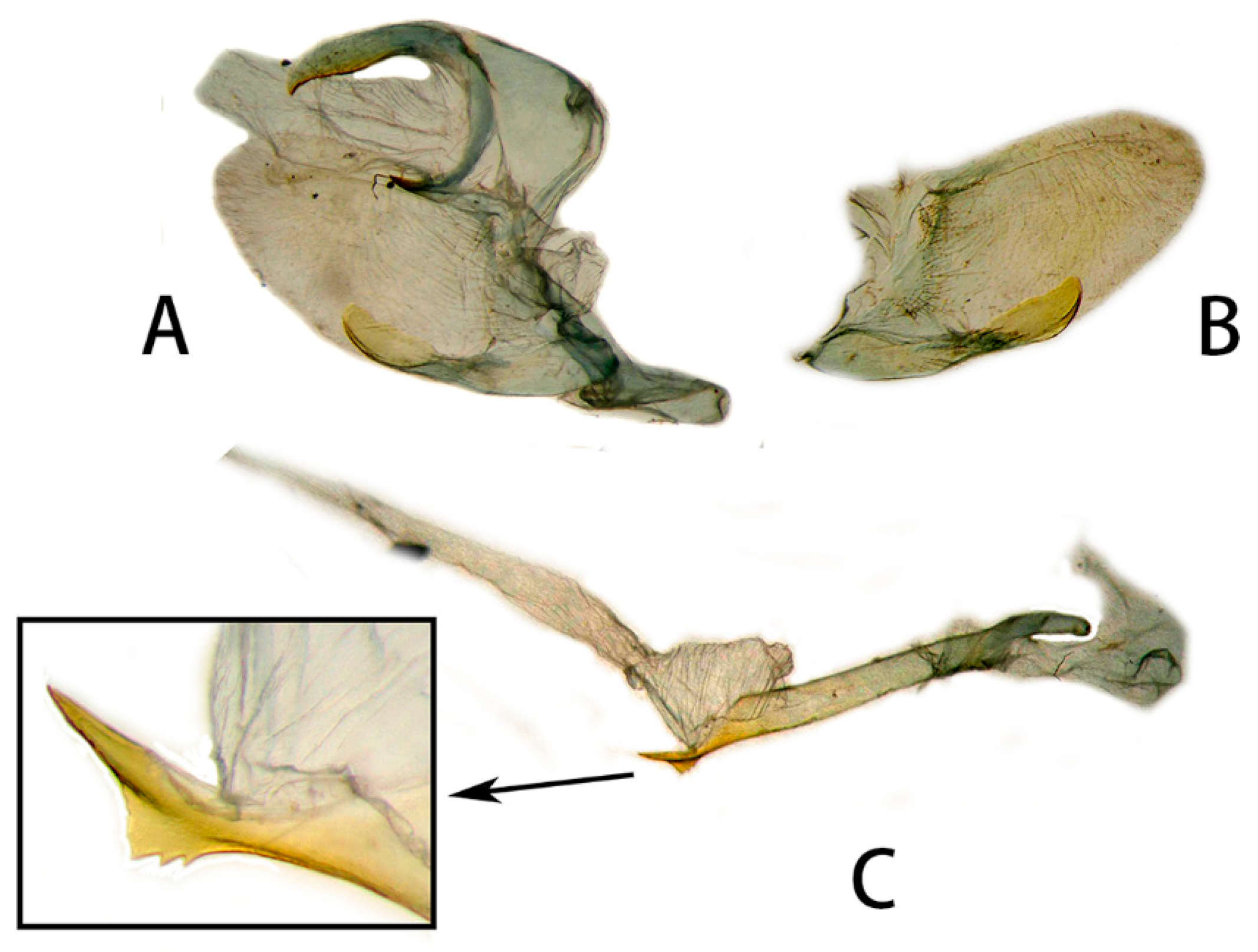
3.2. Morphological Comparison
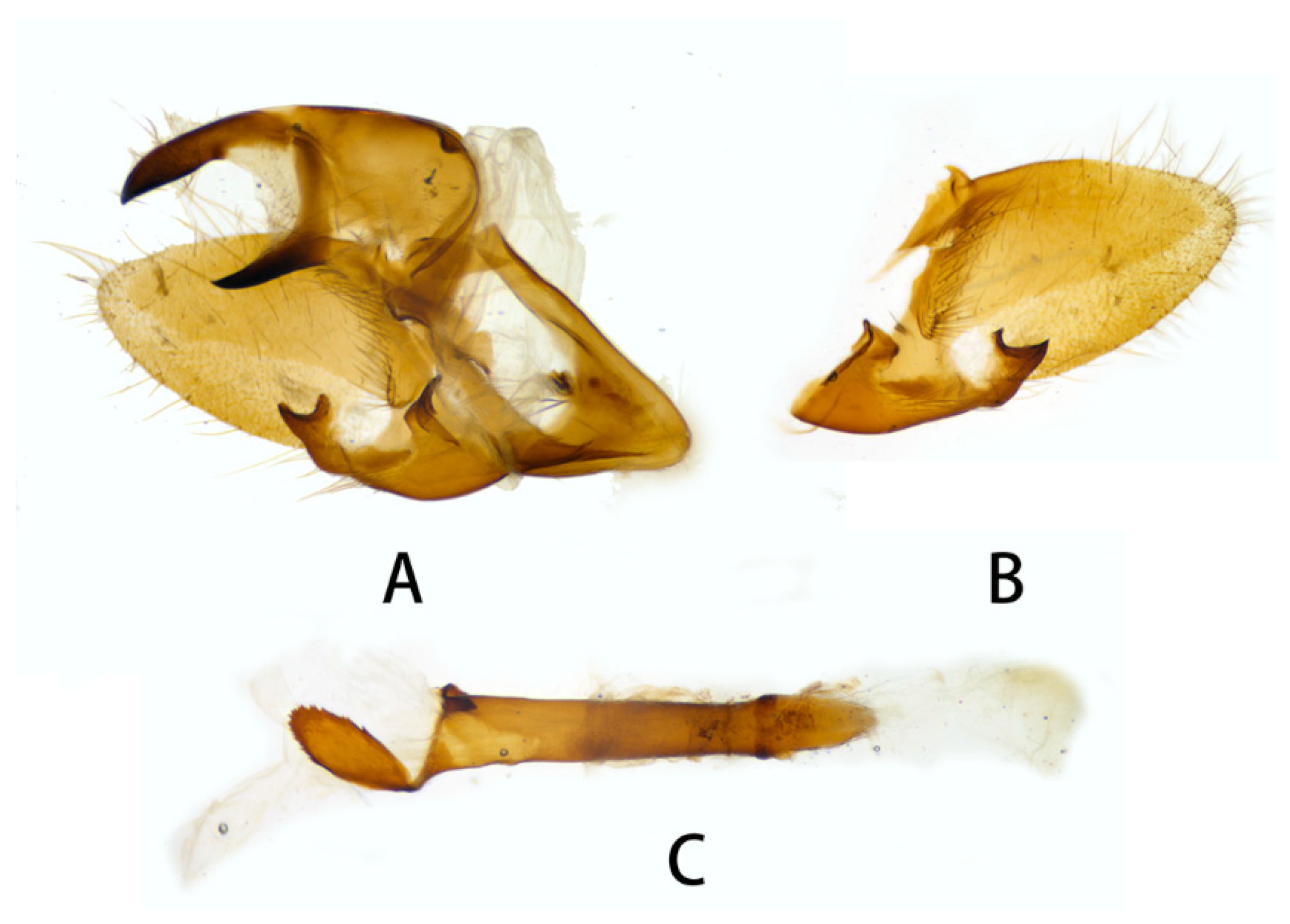
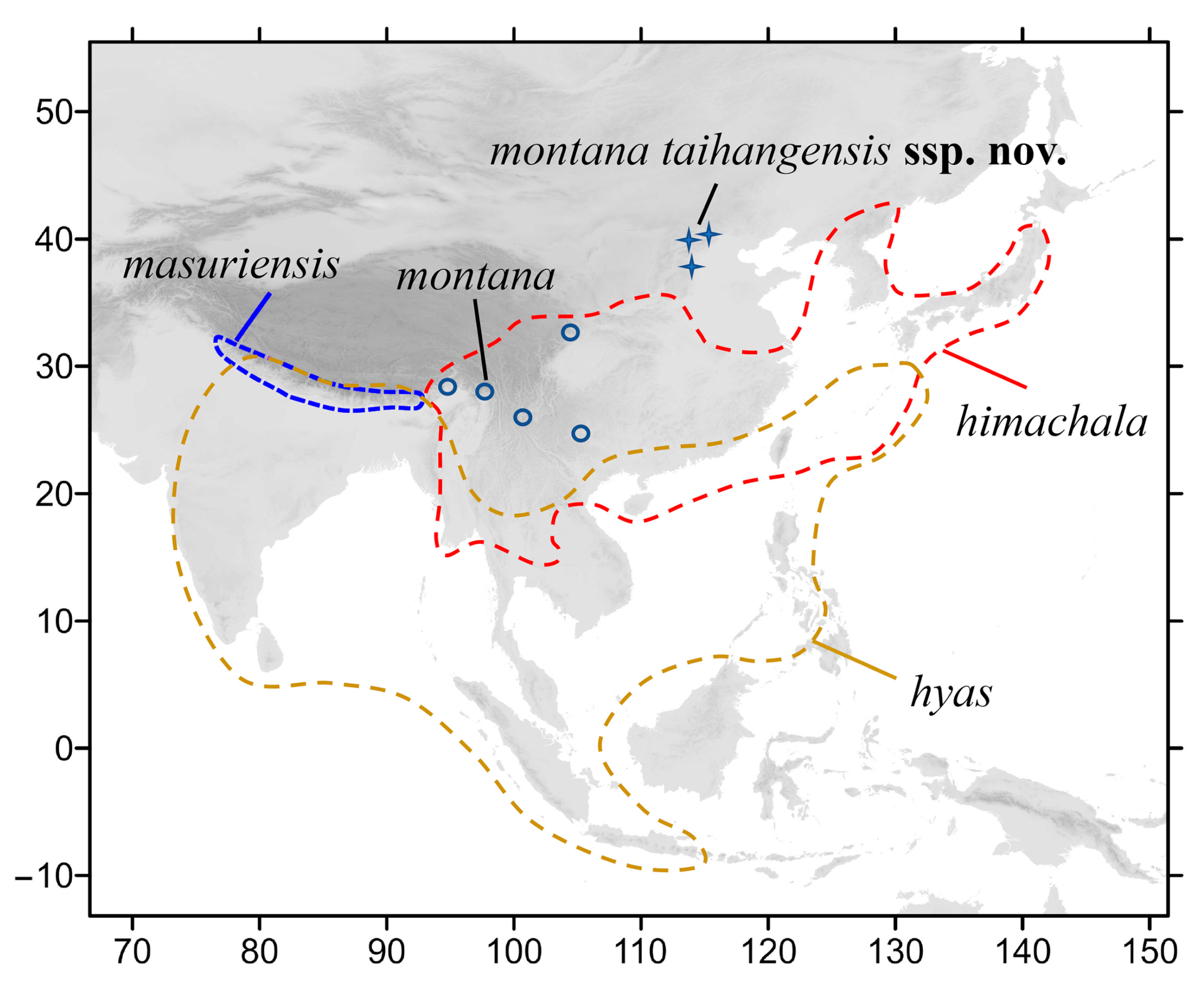
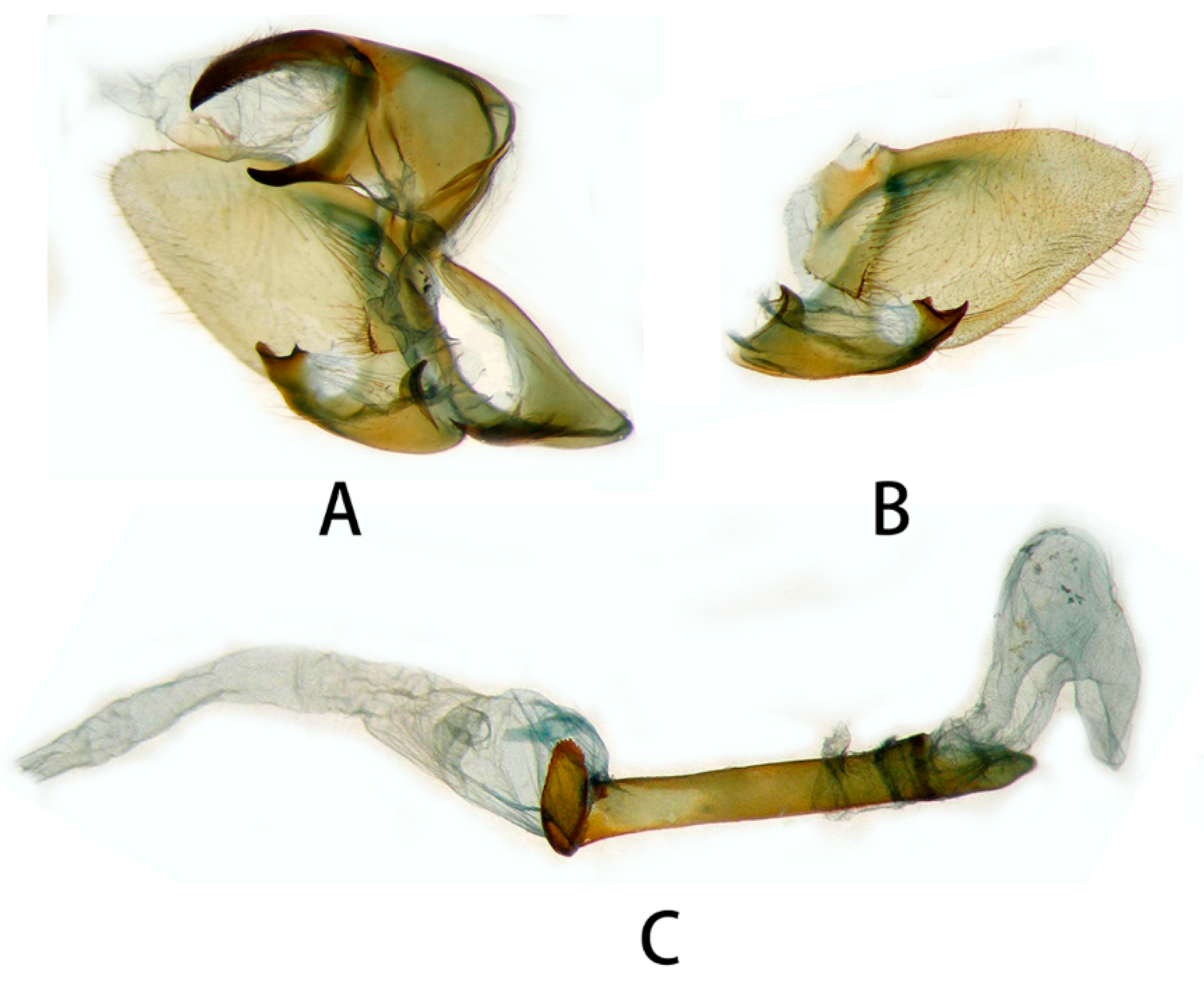
3.2.1. Neogurelca montana montana (Rothschild & Jordan, 1915)
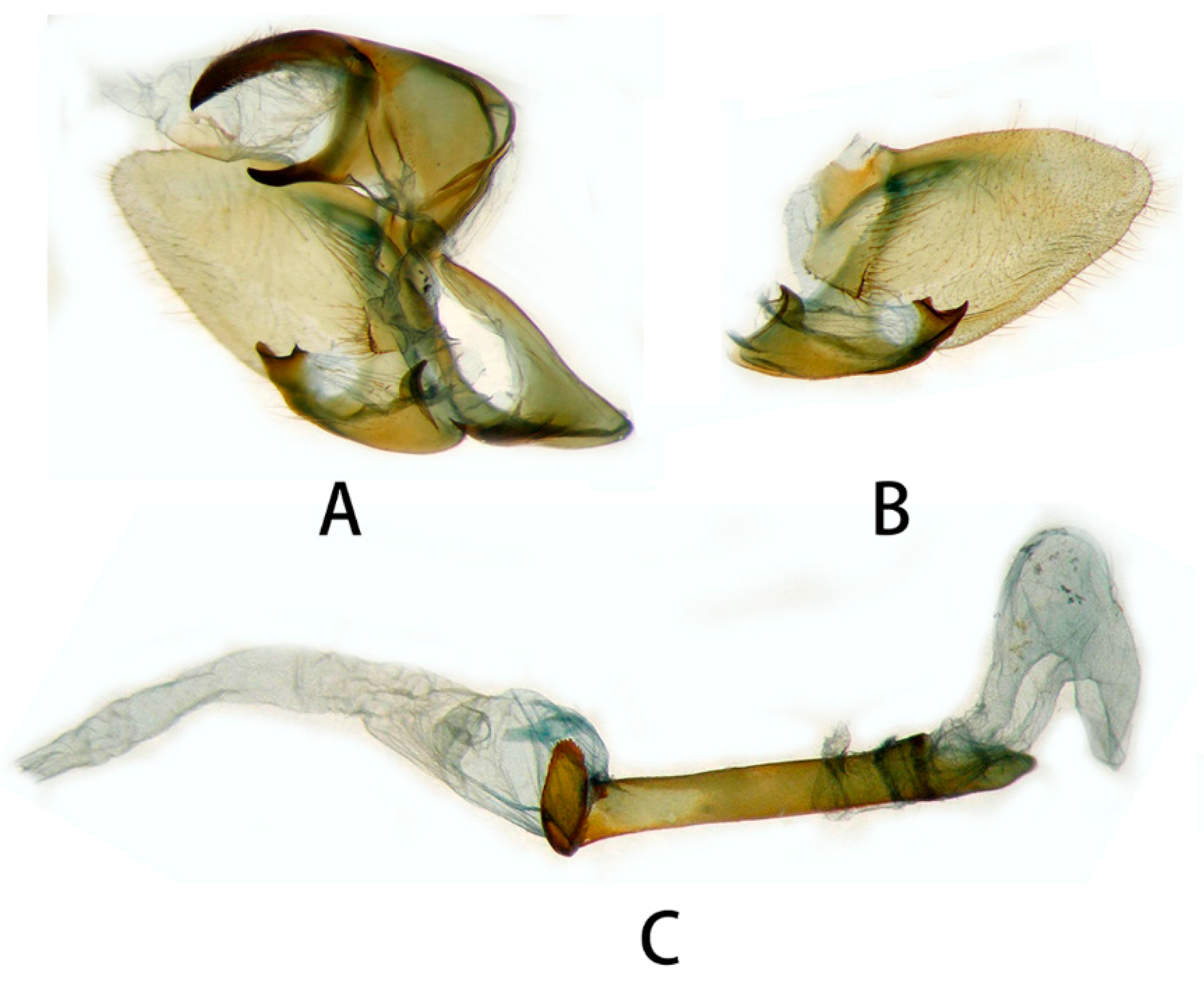
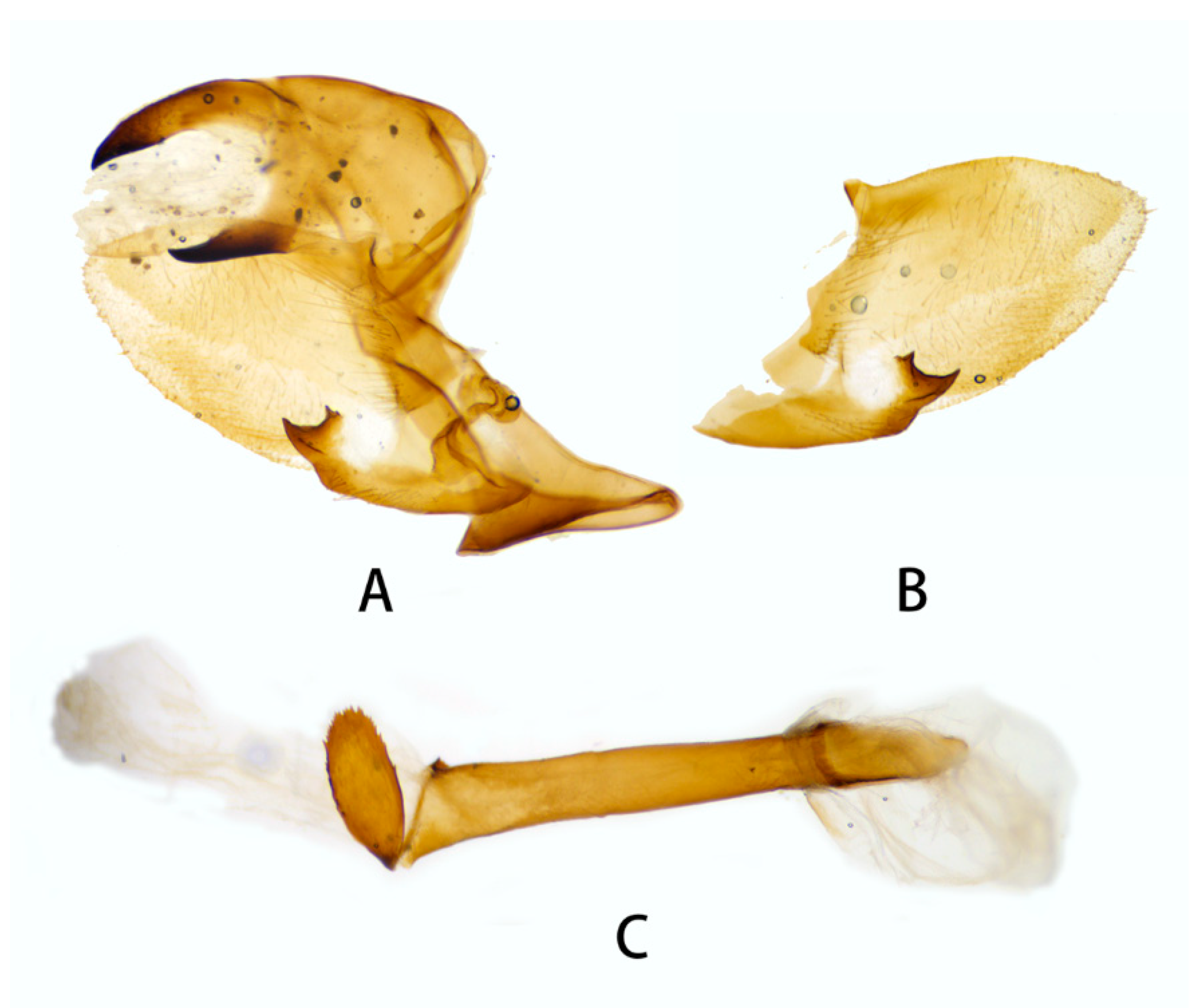
3.2.2. Neogurelca montana taihangensis Xu & He ssp. nov.
3.2.3. Neogurelca himachala (Butler, 1876)
3.2.4. Neogurelca hyas (Walker, 1856)
3.2.5. Neogurelca masuriensis (Butler, 1875)
3.2.6. Diagnostic Characters
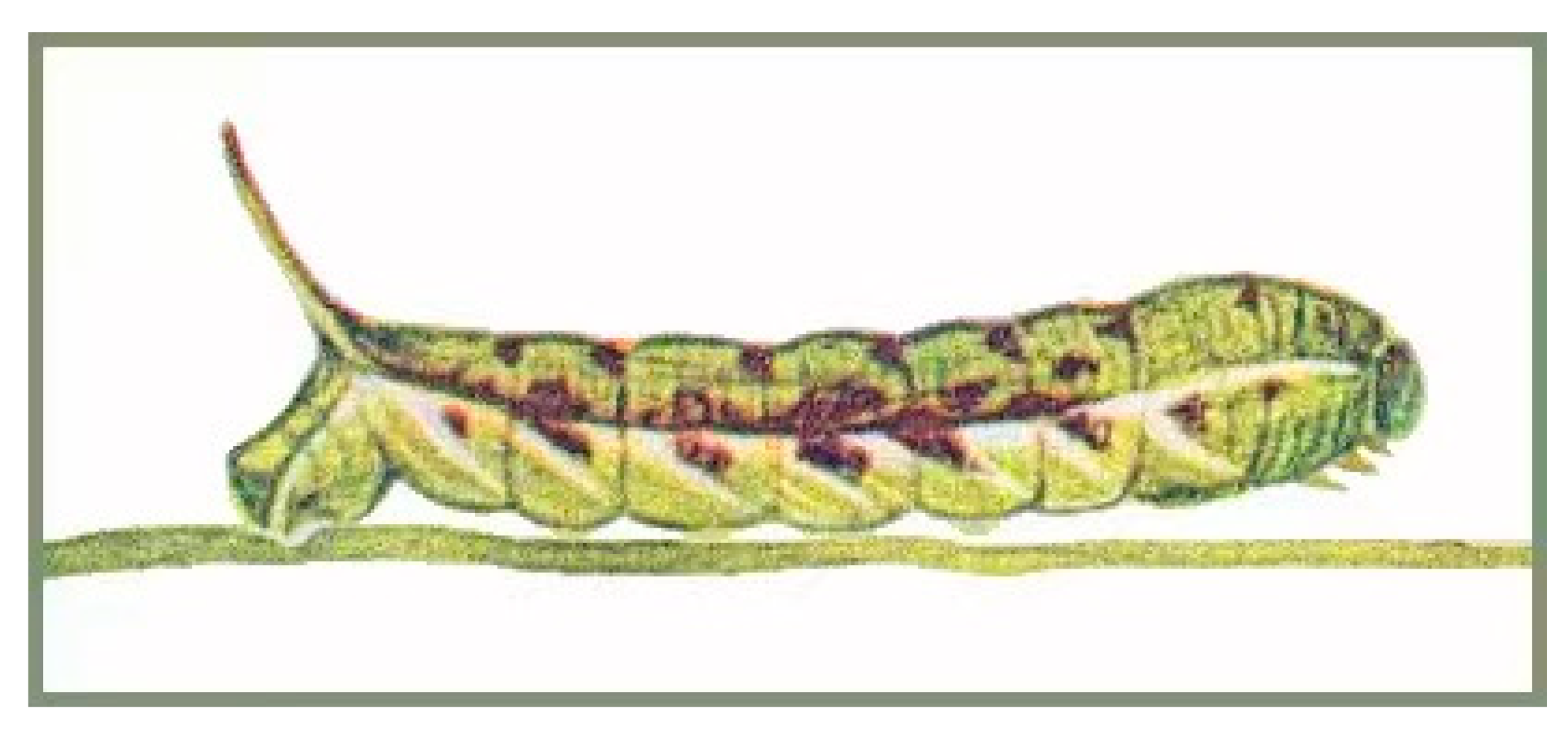
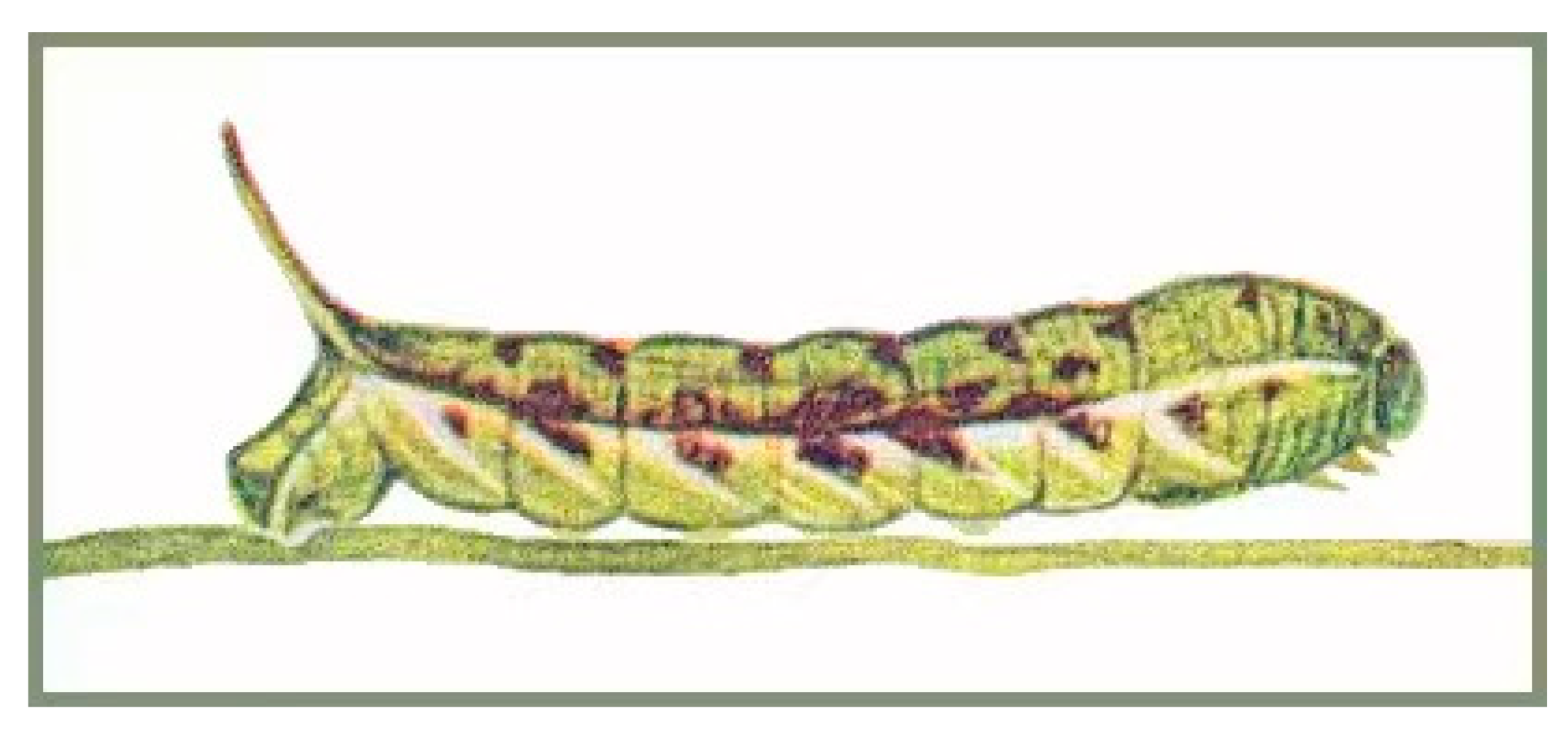
3.3. Life Histories of N. montana taihangensis ssp. nov. and N. himachala
3.3.1. N. montana taihangensis ssp. nov.
3.3.2. Neogurelca himachala
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. List of Neogurelca Specimens Examined
References
- Rothschild, L.W.; Jordan, K. Thirteen new Sphingidae. Novit. Zool. 1915, 22, 281–291. [Google Scholar]
- Hogenes, W.; Treadaway, C.G.T. New hawk moths from the Philippines (Lepidoptera, Sphingidae). Nachrichten Des Entomologischen. Ver. Apollo 1993, 13, 533–552. [Google Scholar]
- Clark, B.P. Some undescribed Sphingidae. Proc. N. Engl. Zoölogical Club 1919, 6, 99–114. [Google Scholar]
- Clark, B.P. Thirty-Three New Sphingidae. Proc. N. Engl. Zoölogical Club 1923, 8, 47–77. [Google Scholar] [CrossRef]
- Haxaire, J. A third species of the genus Neogurelca Hogenes & Treadaway, 1993 for the American continent: Neogurelca serranoi sp. n. (Lepidoptera Sphingidae). Eur. Entomol. 2021, 13, 29–42. [Google Scholar]
- Walker, F. List of the Specimens of Lepidopterous Insects in the Collection of the British Museum; Trustees of the British Museum: London, UK, 1855; Volume 5. [Google Scholar]
- Butler, A.G. Descriptions of New Species of Sphingidae. Proc. Zool. Soc. Lond. 1875, 1875, 238–261. [Google Scholar]
- Butler, A.G. Descriptions of several new species of Sphingidae. Proc. Zool. Soc. Lond. 1876, 1875, 621–623. [Google Scholar]
- Oberthür, C. Nouveaux Lépidoptères du Thibet. Études D’entomologie 1886, 11, 13–38. [Google Scholar]
- Leech, J.H. Butterflies from China, Japan, and Corea: Lycaenidae, Papilionidae, and Hesperiidae; RH Porter: London, UK, 1893. [Google Scholar]
- Mell, R. Biologie und Systematik der Südchinesischen Sphingiden: Zugleich ein Versuch einer Biologie Tropischer Lepidopteren Überhaupt; Friedländer & Sohn: Berlin, Germany, 1922; Volume 1. [Google Scholar]
- Closs, A. Neue Formen aus der Familie der Sphingidae. Int. Entomol. Z. 1917, 11, 153–154. [Google Scholar]
- Jordan, K. On some Oriental Sphingidae. Novit. Zool. 1929, 35, 85–88. [Google Scholar]
- Yu, G.-Y. Moths in Beijing; Science Press: Beijing, China, 2015. [Google Scholar]
- Di, J. The Illustrated Handbook of Insect in Hebei, China II; Science Press: Beijing, China, 2021; Volume II. [Google Scholar]
- Xu, Z.-H.; Guo, S.-B.; Peng, J.-Y. Insect Resources of Xiaowutai Mountain [Xiaowutaishan Kunchong Ziyuan] II; China Forestry Publishing House: Beijing, China, 2013; Volume II. [Google Scholar]
- Kawahara, A.Y.; Mignault, A.A.; Regier, J.C.; Kitching, I.J.; Mitter, C. Phylogeny and biogeography of hawkmoths (Lepidoptera: Sphingidae): Evidence from five nuclear genes. PLoS ONE 2009, 4, e5719. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-B.; Melichar, T.; He, J.-B.; Zhang, C.; Zhang, X.-Y.; Feng, D.-U.; Hu, S.-J. A new species of Rhodambulyx Mell, 1939 (Lepidoptera: Sphingidae) from Southwest Chongqing, China. Zootaxa 2022, 5105, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/Nt. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [PubMed]
- Bertheau, C.; Schuler, H.; Krumboeck, S.; Arthofer, W.; Stauffer, C. Hit or miss in phylogeographic analyses: The case of the cryptic NUMTs. Mol. Ecol. Resour. 2011, 11, 1056–1059. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Ivshin, N.; Krutov, V.; Romanov, D. Three new taxa of the genus Cechetra Zolotuhin & Ryabov, 2012 (Lepidoptera, Sphingidae) from South-East Asia with notes on other species of the genus. Zootaxa 2018, 4450, 1–25. [Google Scholar] [PubMed]
- Cheng, R.; Xue, D.-Y.; Galsworthy, A.; Han, H.-X. Complete mitochondrial genomes throw light on budding speciation in three Biston species (Lepidoptera, Geometridae). Zool. Scr. 2017, 46, 73–84. [Google Scholar] [CrossRef]
- Nässig, W.A.; Naumann, S.; Rougerie, R. Evidence for the existence of three species in the genus Archaeoattacus (Lepidoptera: Saturniidae). J. Res. Lepid. 2010, 43, 37–47. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.-L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.-X.; Wang, G.-T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.; Larget, B.; Alfaro, M. Bayesian phylogenetic model selection using reversible jump Markov chain Monte Carlo. Mol. Biol. Evol. 2004, 21, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lewis, P.O.; Fan, Y.; Kuo, L.; Chen, M.-H. Improving marginal likelihood estimation for Bayesian phylogenetic model selection. Syst. Biol. 2011, 60, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Godfray, H.C.J. Species-level para-and polyphyly in DNA barcode gene trees: Strong operational bias in European Lepidoptera. Syst. Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef]
- Hu, S.-J.; Cotton, A.M.; Condamine, F.L.; Duan, K.; Wang, R.-J.; Hsu, Y.-F.; Zhang, X.; Cao, J. Revision of Pazala Moore, 1888: The Graphium (Pazala) mandarinus (Oberthür, 1879) group, with treatments of known taxa and descriptions of new species and new subspecies (Lepidoptera: Papilionidae). Zootaxa 2018, 4441, 401–446. [Google Scholar] [CrossRef]
- Bell, T.R.D.; Scott, F.B. The Fauna of British India, Including Ceylon and Burma. Moths; Taylor and Francis: London, UK, 1937; Volume 5. [Google Scholar]
- Sphingidae; CSTR: 19715.10.6_Sphingidae [Internet]. National Animal Collection Resource Center, Institute of Zoology, Chinese Academy of Sciences. 2023. Available online: http://museum.ioz.ac.cn/DataSggregation.aspx?lk=6_Sphingidae (accessed on 11 October 2023).
- Irungbam, J.S.; Irungbam, M.J. Contributions to the knowledge of moths of Bombycoidea Latreille, 1802 (Lepidoptera: Heterocera) of Bhutan with new records. J. Threat. Taxa 2019, 11, 14022–14050. [Google Scholar] [CrossRef]
- Eberhard, W.G. Sexual Selection and Animal Genitalia; Harvard University Press: Cambridge, MA, USA, 1985. [Google Scholar]
- Ah-King, M.; Barron, A.B.; Herberstein, M.E. Genital evolution: Why are females still understudied? PLoS Biol. 2014, 12, e1001851. [Google Scholar] [CrossRef]
- Hosken, D.J.; Stockley, P. Sexual selection and genital evolution. Trends Ecol. Evol. 2004, 19, 87–93. [Google Scholar] [CrossRef]
- Sánchez, V.; Hernández-Baños, B.E.; Cordero, C. The evolution of a female genital trait widely distributed in the Lepidoptera: Comparative evidence for an effect of sexual coevolution. PLoS ONE 2011, 6, e22642. [Google Scholar] [CrossRef]
- Komárek, S. Mimicry, Aposematism and Related Phenomena: Mimetism in Nature and the History of Its Study; Lincom Europa: Munich, Germany, 2003. [Google Scholar]
- Wallace, A.R. Darwinism: An Exposition of the Theory of Natural Selection with Some of Its Applications; Cosimo, Inc.: New York, NY, USA, 2007. [Google Scholar]
- Wang, S.-T.; Teng, D.-Q.; Li, X.-Y.; Yang, P.-W.; Da, W.; Zhang, Y.-M.; Zhang, Y.-B.; Liu, G.-C.; Zhang, X.-S.; Wan, W.-T. The evolution and diversification of oakleaf butterflies. Cell 2022, 185, 3138–3152. [Google Scholar] [CrossRef]
- Ma, T.-J.; Bai, J.; Li, T.-T.; Chen, S.; Ma, X.-D.; Yin, J.; Jiang, X.-S. Light-driven dynamic surface wrinkles for adaptive visible camouflage. Proc. Natl. Acad. Sci. USA 2021, 118, e2114345118. [Google Scholar] [CrossRef] [PubMed]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon (Sample Code) | Locality | Collecting Date | GenBank No. | Bold ID |
|---|---|---|---|---|
| N. himachala (NHI02) | Wuhan, Hubei, China | 2018-XI | OR235183 | |
| N. himachala (NHI03) | Wuhan, Hubei, China | 2018-XI | OR235184 | |
| N. himachala (NHI04) | Wuhan, Hubei, China | 2018-XI | OR235185 | |
| N. himachala (NHI06) | Baotianman, Henan, China | 2021-IX | OR235186 | |
| N. himachala (NHI10) | Niubeiliang, Shaanxi, China | 2017-VII | OR235187 | |
| N. himachala | Zhejiang, China | 2003-I | IJK-04-0004 | |
| N. hyas (NHA01) | Lingao, Hainan, China | 2023-VI | OR235191 | |
| N. hyas (NHA02) | Lingao, Hainan, China | 2023-VI | OR235192 | |
| N. hyas | Lomblen Island, Indonesia | 2006-XI | BC-Mel 0555 | |
| N. hyas | Leyte Islands, Philippines | 2003-VIII | BC-Hax3980 | |
| N. montana montana (NMO01) | Shangri-La, Yunnan, China | 2017-VIII | OR235188 | |
| N. montana montana (NMO02) | Cangshan, Dali, Yunnan, China | 2020-XI | OR235189 | |
| N. montana montana (NMO03) | BingZhongluo, Yunnan, China | 2021-VII | OR235190 | |
| N. montana taihangensis ssp. nov. (BHS01) | Baihuashan, Beijing, China | 2018-X | OR235181 | |
| N. montana taihangensis ssp. nov. (BHS04) | Yunmengshan, Beijing, China | 2018-VII | OR235182 | |
| Sphingonaepiopsis pumilio | Sumatra, Indonesia | 2004-XI | BC-Mel 0221 | |
| Sphingonaepiopsis gorgoniades | Ukraine | 2005-I | IJK-05-0003 |
| 1 | 2 | 3 | 4 | |
|---|---|---|---|---|
| 1. Neogurelca montana taihangensis ssp. nov. | ||||
| 2. Neogurelca montana montana | 0.9 | |||
| 3. Neogurelca himachala | 5.9 | 5.6 | ||
| 4. Neogurelca hyas | 9.2 | 8.7 | 10.0 |
| Taxon | Assessment | Tangles |
|---|---|---|
| 1. Neogurelca montana taihangensis ssp. nov. | monophyletic | — |
| 2. Neogurelca montana montana | monophyletic | — |
| 3a. Neogurelca himachala | monophyletic | — |
| 3b. Neogurelca hyas | monophyletic | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.-B.; He, J.-B.; Yang, N.; Kitching, I.J.; Hu, S.-J. Review of the Narrow-Banded Hawkmoth, Neogurelca montana (Rothschild & Jordan, 1915) (Lepidoptera: Sphingidae) in China, with Morphological and Phylogenetic Analysis. Insects 2023, 14, 818. https://doi.org/10.3390/insects14100818
Xu Z-B, He J-B, Yang N, Kitching IJ, Hu S-J. Review of the Narrow-Banded Hawkmoth, Neogurelca montana (Rothschild & Jordan, 1915) (Lepidoptera: Sphingidae) in China, with Morphological and Phylogenetic Analysis. Insects. 2023; 14(10):818. https://doi.org/10.3390/insects14100818
Chicago/Turabian StyleXu, Zhen-Bang, Ji-Bai He, Nan Yang, Ian J. Kitching, and Shao-Ji Hu. 2023. "Review of the Narrow-Banded Hawkmoth, Neogurelca montana (Rothschild & Jordan, 1915) (Lepidoptera: Sphingidae) in China, with Morphological and Phylogenetic Analysis" Insects 14, no. 10: 818. https://doi.org/10.3390/insects14100818
APA StyleXu, Z.-B., He, J.-B., Yang, N., Kitching, I. J., & Hu, S.-J. (2023). Review of the Narrow-Banded Hawkmoth, Neogurelca montana (Rothschild & Jordan, 1915) (Lepidoptera: Sphingidae) in China, with Morphological and Phylogenetic Analysis. Insects, 14(10), 818. https://doi.org/10.3390/insects14100818