
Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples/Yellow Mealworm Colony
2.2. Diet Preparation
2.3. Larval Growth and Development Measurements (Feeding Trial)
2.4. Statistical Analysis
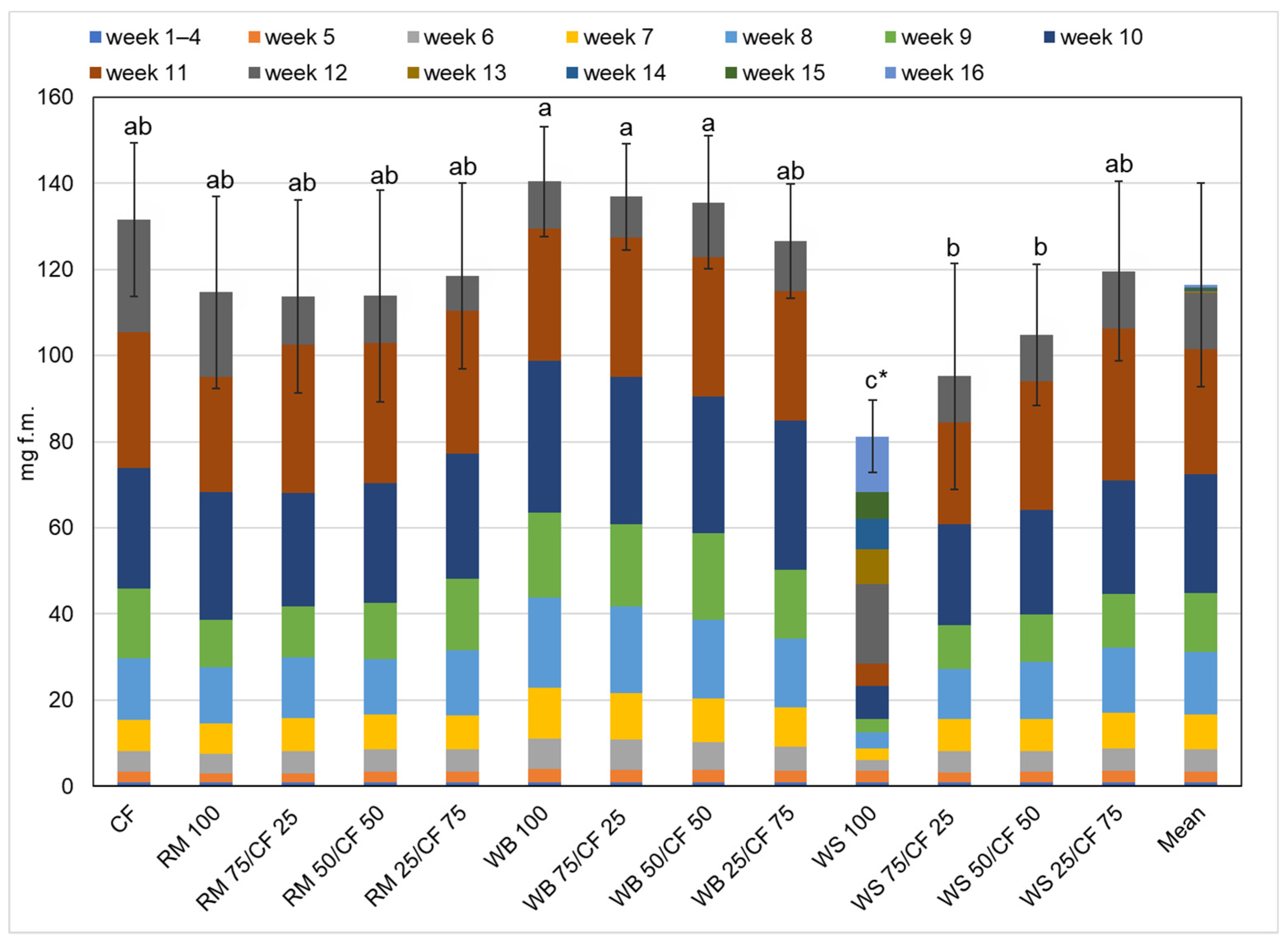
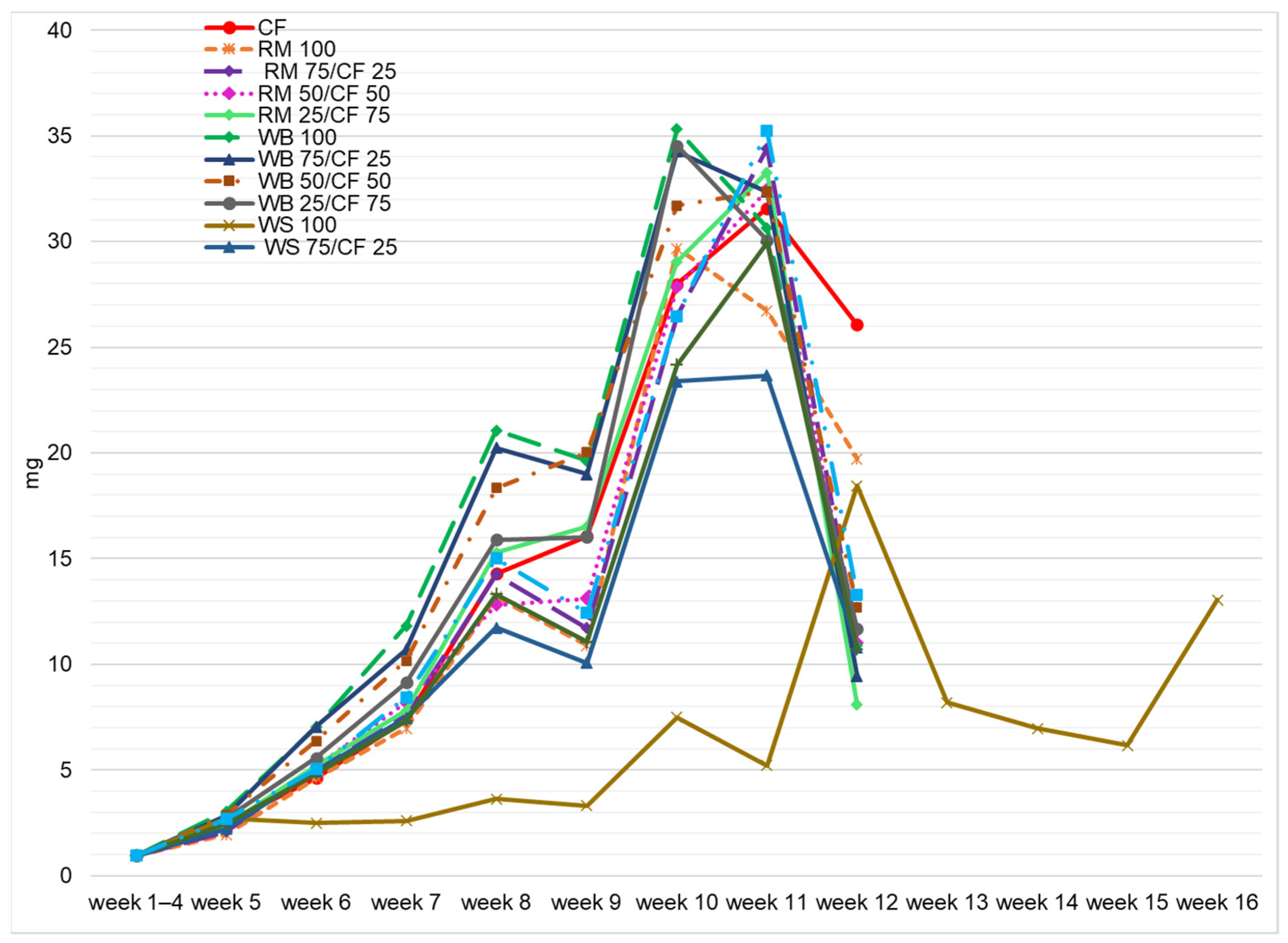
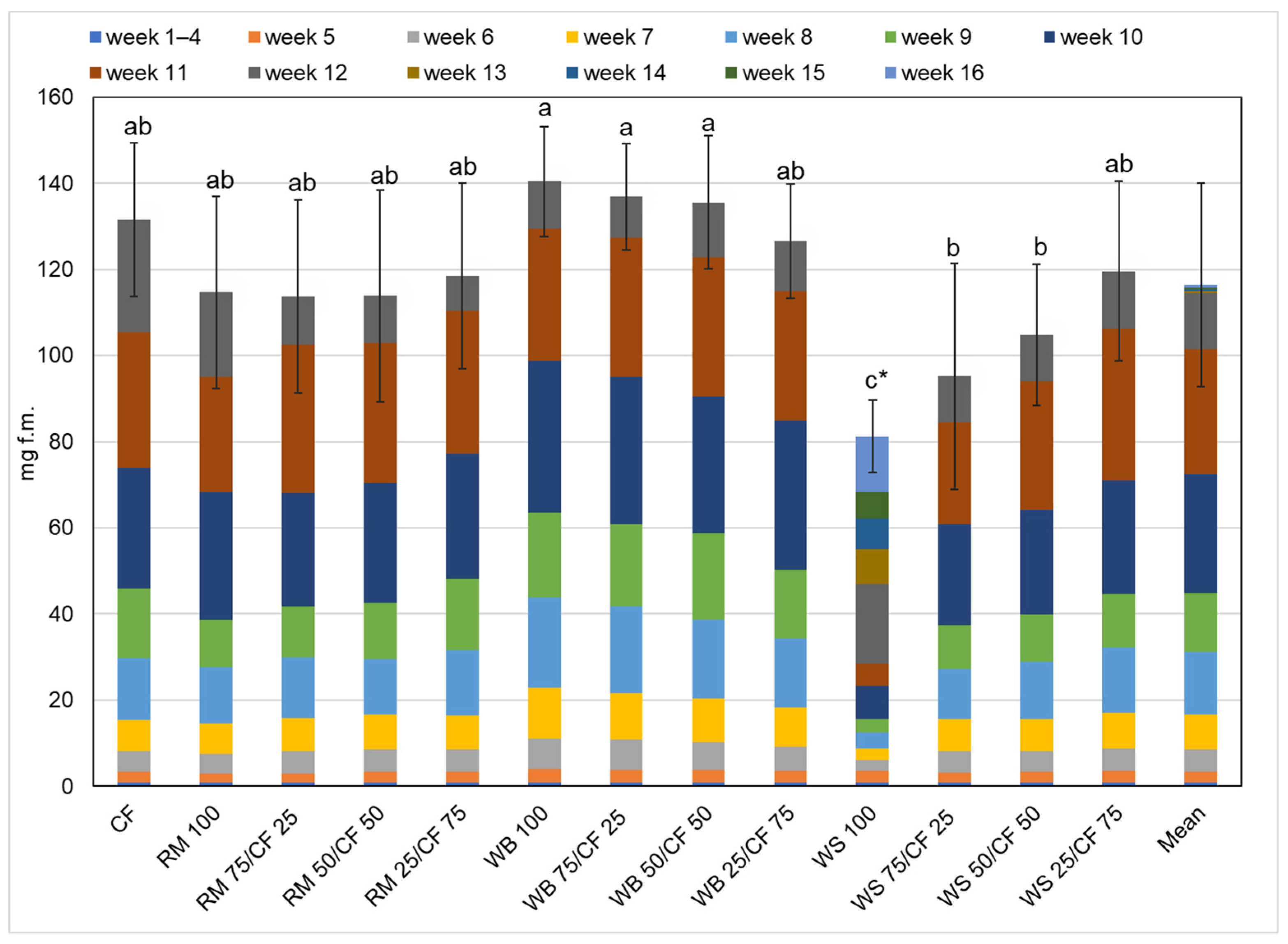
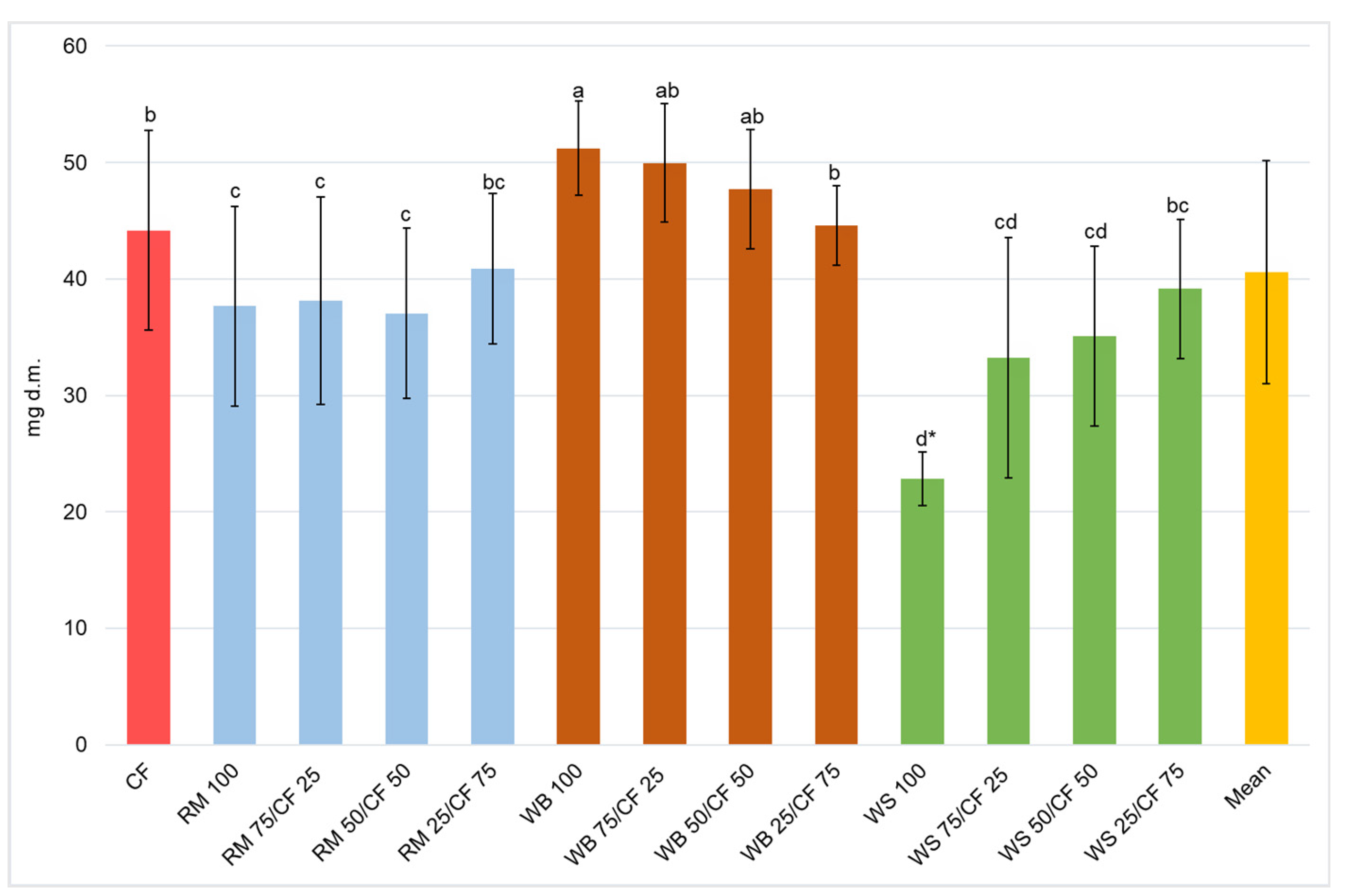
3. Results and Discussion
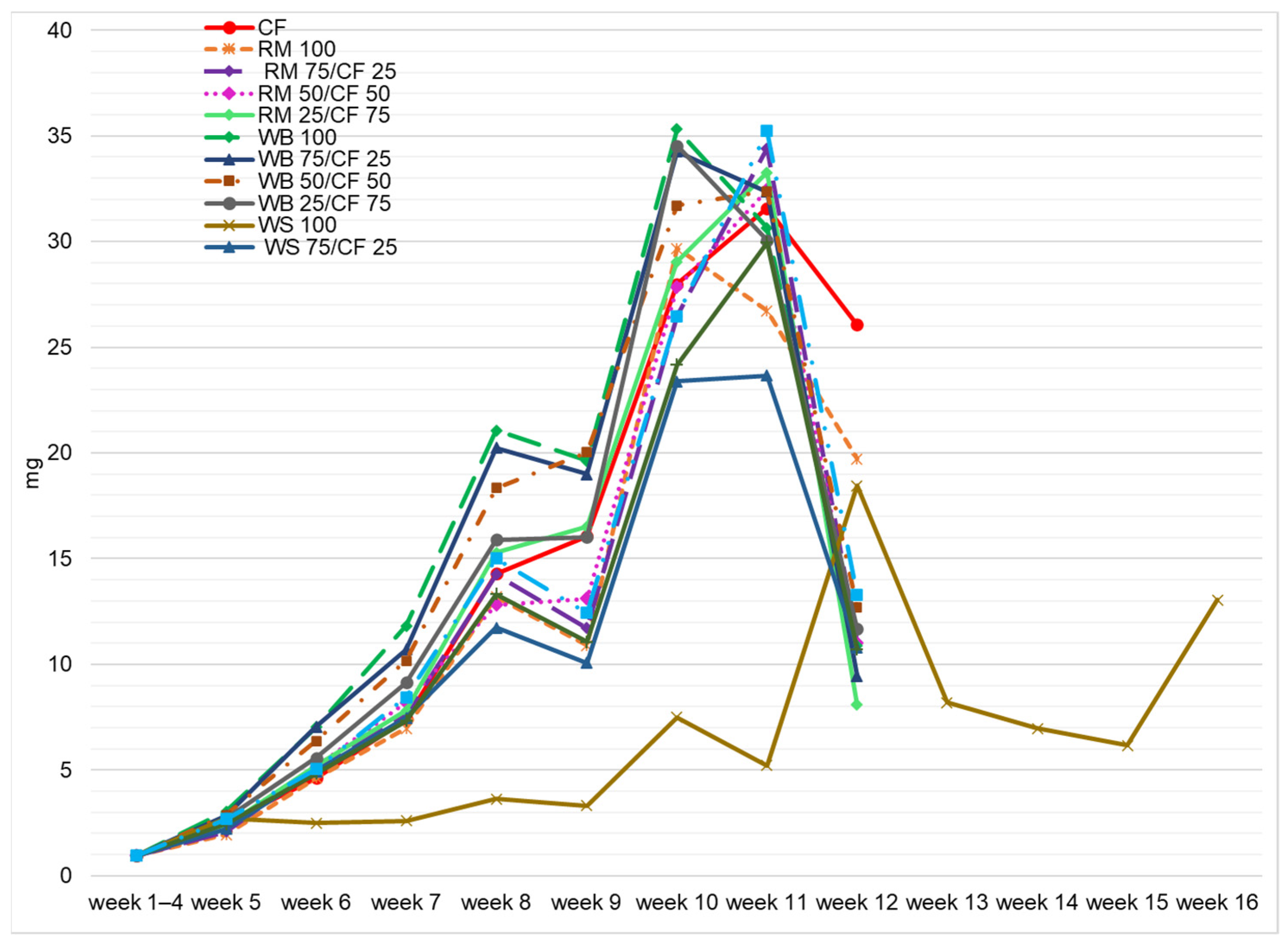
3.1. Larvae Development Time
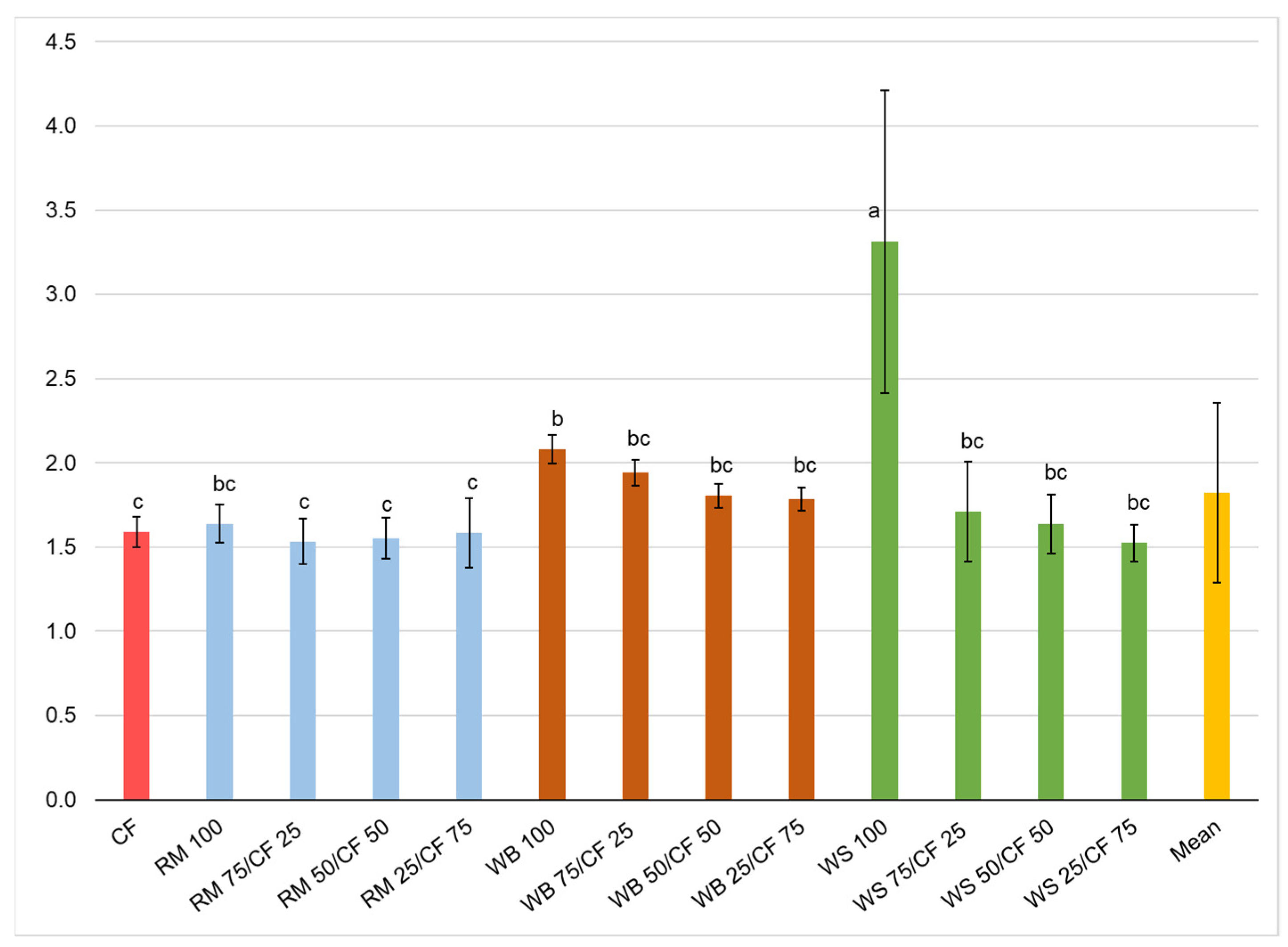
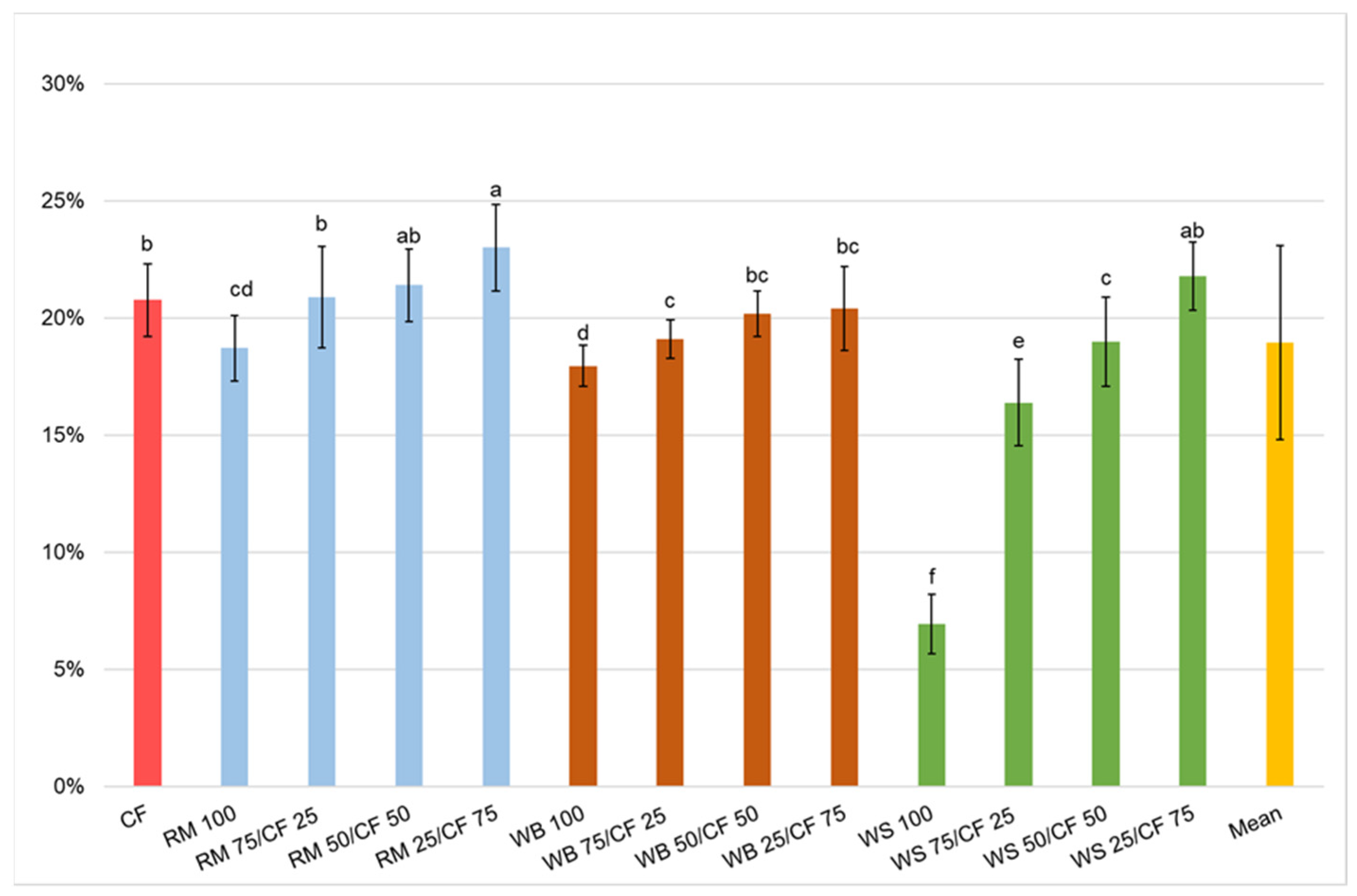
3.2. Growth Performance
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaza, S.; Yao, L.; Bhada-Tata, P.; Van Woerden, F. What a Waste 2.0: A Global Snapshot of Solid Waste Management to 2050; World Bank Publications: Washington, DC, USA, 2018. [Google Scholar]
- Gustafsson, J.; Cederberg, C.; Sonesson, U.; Emanuelsson, A. The Methodology of the FAO Study: Global Food Losses and Food Waste-extent, Causes and Prevention–FAO, 2011; SIK Institute för Livsmedel och Bioteknik: Boras, Sweden, 2013. [Google Scholar]
- World Health Organization. The State of Food Security and Nutrition in the World 2018: Building Climate Resilience for Food Security and Nutrition; FAO: Rome, Italy, 2018. [Google Scholar]
- Save Food (FAO). Global initiative on food loss and waste reduction. In Key Facts on Food Loss and Waste You Should Know; FAO: Rome, Italy, 2016. [Google Scholar]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Czachorowski, S.; Peni, D. Will yellow mealworm become a source of safe proteins for Europe? Agriculture 2020, 10, 233. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J.; González, E.A.; Hernández, A.R.; Pino, J.M. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 2002, 95, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0222043. [Google Scholar] [CrossRef] [PubMed]
- Van Broekhoven, S.; Oonincx, D.G.A.B.; van Huis, A.; van Loon, J.J.A. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Chen, G.; Qiao, L.; Li, J.; Liu, B.; Liu, Z.; Li, M.; Liu, X. Growth performance and nutritional profile of mealworms reared on corn stover, soybean meal, and distillers’ grains. Eur. Food Res. Technol. 2019, 245, 2631–2640. [Google Scholar] [CrossRef]
- Melis, R.; Braca, A.; Sanna, R.; Spada, S.; Mulas, G.; Fadda, M.L.; Sassu, M.M.; Serra, G.; Anedda, R. Metabolic response of yellow mealworm larvae to two alternative rearing substrates. Metabolomics 2019, 15, 113. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio molitor and Alphitobius diaperinus larvae on seed cleaning process byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D. Growth potential of yellow mealworm reared on industrial residues. Agriculture 2020, 10, 599. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J.; Peni, D.; Aljewicz, M. The influence of different diets on growth and nutritional composition of yellow mealworm. In Proceedings of the Insects to Feed the World, Quebec City, QC, Canada, 12–16 June 2022. [Google Scholar]
- Chemello, G.; Renna, M.; Caimi, C.; Guerreiro, I.; Oliva-Teles, A.; Enes, P.; Biasato, I.; Schiavone, A.; Gai, F.; Gasco, L. Partially defatted Tenebrio molitor larva meal in diets for grow-out rainbow trout, Oncorhynchus mykiss (Walbaum): Effects on growth performance, diet digestibility and metabolic responses. Animals 2020, 10, 229. [Google Scholar] [CrossRef]
- Biasato, I.; Gasco, L.; De Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Sterpone, L. Yellow mealworm larvae (Tenebrio molitor) inclusion in diets for male broiler chickens: Effects on growth performance, gut morphology, and histological findings. Poult. Sci. 2018, 97, 540–548. [Google Scholar] [CrossRef]
- Stolarski, M.J.; Warmiński, K.; Krzyżaniak, M.; Tyśkiewicz, K.; Olba-Zięty, E.; Graban, Ł.; Lajszner, W.; Załuski, D.; Wiejak, R.; Kamiński, P. How does extraction of biologically active substances with supercritical carbon dioxide affect lignocellulosic biomass properties? Wood Sci. Technol. 2020, 54, 519–546. [Google Scholar] [CrossRef]
- Miech, P.; Berggren, Å.; Lindberg, J.E.; Chhay, T.; Khieu, B.; Jansson, A. Growth and survival of reared Cambodian field crickets (Teleogryllus testaceus) fed weeds, agricultural and food industry by-products. J. Insects Food Feed 2016, 2, 285–292. [Google Scholar] [CrossRef]
- Waldbauer, G.P. The consumption and utilization of food by insects. In Advances in Insect Physiology; Academic Press: Cambridge, MA, USA, 1968; Volume 5, pp. 229–288. [Google Scholar] [CrossRef]
- Mlček, J.; Adámek, M.; Adámková, A.; Matyáš, J.; Bučková, M.; Mrázková, M.; Vícha, R.; Vychodil, R.; Knížková, I.; Volek, Z. Feed parameters influencing the breeding of mealworms (Tenebrio molitor). Sustainability 2021, 13, 12992. [Google Scholar] [CrossRef]
- Silva, L.B.; de Souza, R.G.; da Silva, S.R.; Feitosa, A.d.C.; Lopes, E.C.; Lima, S.B.P.; Dourado, L.R.B.; Pavan, B.E. Development of Tenebrio molitor (Coleoptera: Tenebrionidae) on poultry litter-based diets: Effect on chemical composition of larvae. J. Insect Sci. 2021, 21, 7. [Google Scholar] [CrossRef]
- Deruytter, D.; Coudron, C.L. The effects of density on the growth, survival and feed conversion of Tenebrio molitor larvae. J. Insects Food Feed 2022, 8, 141–146. [Google Scholar] [CrossRef]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth performance and nutrient composition of mealworms (Tenebrio Molitor) fed on fresh plant materials-supplemented diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef]
- Yang, S.-S.; Brandon, A.M.; Flanagan, J.C.A.; Yang, J.; Ning, D.; Cai, S.-Y.; Fan, H.-Q.; Wang, Z.-Y.; Ren, J.; Benbow, E. Biodegradation of polystyrene wastes in yellow mealworms (larvae of Tenebrio molitor Linnaeus): Factors affecting biodegradation rates and the ability of polystyrene-fed larvae to complete their life cycle. Chemosphere 2018, 191, 979–989. [Google Scholar] [CrossRef]
- Eberle, S.; Schaden, L.-M.; Tintner, J.; Stauffer, C.; Schebeck, M. Effect of temperature and photoperiod on development, survival, and growth rate of mealworms, Tenebrio molitor. Insects 2022, 13, 321. [Google Scholar] [CrossRef]
- Punzo, F.; Mutchmor, J.A. Effects of temperature, relative humidity and period of exposure on the survival capacity of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kansas Entomol. Soc. 1980, 53, 260–270. [Google Scholar]
- Kim, S.Y.; Kim, H.G.; Yoon, H.J.; Lee, K.Y.; Kim, N.J. Nutritional analysis of alternative feed ingredients and their effects on the larval growth of Tenebrio molitor (Coleoptera: Tenebrionidae). Entomol. Res. 2017, 47, 194–202. [Google Scholar] [CrossRef]
- Adamaki-Sotiraki, C.; Rumbos, C.I.; Athanassiou, C.G. Developmental plasticity among strains of the yellow mealworm, Tenebrio molitor L., under dry conditions. J. Insects Food Feed 2021, 8, 281–288. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Adamaki-Sotiraki, C.; Gourgouta, M.; Karapanagiotidis, I.T.; Asimaki, A.; Mente, E.; Athanassiou, C.G. Strain matters: Strain effect on the larval growth and performance of the yellow mealworm, Tenebrio molitor L. J. Insects Food Feed 2021, 7, 1195–1205. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Z.; Liu, H. Feasibility of feeding yellow mealworm (Tenebrio molitor L.) in bioregenerative life support systems as a source of animal protein for humans. Acta Astronaut. 2013, 92, 103–109. [Google Scholar] [CrossRef]
- House, H.L. Effects of low levels of the nutrient content of a food and of nutrient imbalance on the feeding and the nutrition of a phytophagous larva, Celerio euphorbiae (Linnaeus) (Lepidoptera: Sphingidae). Can. Entomol. 1965, 97, 62–68. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Ash (% d.m.) | Fibre Content (% d.m.) | Crude Fat (% d.m.) | Protein (% d.m.) | NFE (% d.m.) | |
|---|---|---|---|---|---|---|
| 1. | CF 100 | 12.89 | 3.23 | 2.38 | 20.11 | 61.39 |
| 2. | RM 100 | 7.39 | 11.12 | 1.78 | 35.52 | 44.19 |
| 3. | RM 75/CF 25 | 8.77 | 9.15 | 1.93 | 31.67 | 48.48 |
| 4. | RM 50/CF 50 | 10.14 | 7.18 | 2.08 | 27.82 | 52.78 |
| 5. | RM 25/CF 75 | 11.52 | 5.2 | 2.23 | 23.96 | 57.09 |
| 6. | WB 100 | 5.82 | 8.68 | 2.90 | 18.63 | 63.97 |
| 7. | WB 75/CF 25 | 7.59 | 7.32 | 2.77 | 19.00 | 63.32 |
| 8. | WB 50/CF 50 | 9.36 | 5.96 | 2.64 | 19.38 | 62.66 |
| 9. | WB 25/CF 75 | 11.12 | 4.59 | 2.51 | 19.74 | 62.04 |
| 10. | WS 100 | 9.49 | 22.64 | 0.76 | 10.56 | 56.55 |
| 11. | WS 75/CF 25 | 10.34 | 17.79 | 1.17 | 12.95 | 57.75 |
| 12. | WS 50/CF 50 | 11.19 | 12.94 | 1.57 | 15.34 | 58.96 |
| 13. | WS 25/CF 75 | 12.04 | 8.08 | 1.98 | 17.72 | 60.18 |
| Source of Variation | Survival | Dry Weight | Fresh Weight | FCR | ECI |
|---|---|---|---|---|---|
| df | 12 | ||||
| F | 1.7 | 9.416 | 6.234 | 25.219 | 60.44 |
| p | 0.07 | <0.001 | <0.001 | <0.001 | <0.001 |
| No. | Diet | Survival (%) | Larval Period (Days) * |
|---|---|---|---|
| 1. | CF (control) | 96.0 ± 2.6 | 76.8 ± 3.2 |
| 2. | RM 100 | 95.3 ± 2.8 | 78.5 ± 0.5 |
| 3. | RM 75/CF 25 | 96.9 ± 1.4 | 76.0 ± 1.0 |
| 4. | RM 50/CF 50 | 96.7 ± 2.4 | 76.0 ± 1.0 |
| 5. | RM 25/CF 75 | 96.6 ± 1.9 | 76.0 ± 1.0 |
| 6. | WB 100 | 92.0 ± 6.7 | 74.7 ± 0.3 |
| 7. | WB 75/CF 25 | 96.0 ± 3.7 | 76.0 ± 1.0 |
| 8. | WB 50/CF 50 | 95.8 ± 2.5 | 76.0 ± 1.0 |
| 9. | WB 25/CF 75 | 95.8 ± 2.3 | 73.0 ± 2.0 |
| 10. | WS 100 | 95.8 ± 3.2 | 115.0 ± 1.0 |
| 11. | WS 75/CF 25 | 96.7 ± 2.4 | 81.5 ± 4.5 |
| 12. | WS 50/CF 50 | 96.2 ± 1.5 | 76.5 ± 1.5 |
| 13. | WS 25/CF 75 | 97.7 ± 2.2 | 75.0 ± 0.0 |
| Mean | 96.0 ± 3.1 | 79.3 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bordiean, A.; Krzyżaniak, M.; Stolarski, M.J. Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results. Insects 2022, 13, 810. https://doi.org/10.3390/insects13090810
Bordiean A, Krzyżaniak M, Stolarski MJ. Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results. Insects. 2022; 13(9):810. https://doi.org/10.3390/insects13090810
Chicago/Turabian StyleBordiean, Anna, Michał Krzyżaniak, and Mariusz Jerzy Stolarski. 2022. "Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results" Insects 13, no. 9: 810. https://doi.org/10.3390/insects13090810
APA StyleBordiean, A., Krzyżaniak, M., & Stolarski, M. J. (2022). Bioconversion Potential of Agro-Industrial Byproducts by Tenebrio molitor—Long-Term Results. Insects, 13(9), 810. https://doi.org/10.3390/insects13090810