

Mass Trapping and Larval Source Management for Mosquito Elimination on Small Maldivian Islands
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
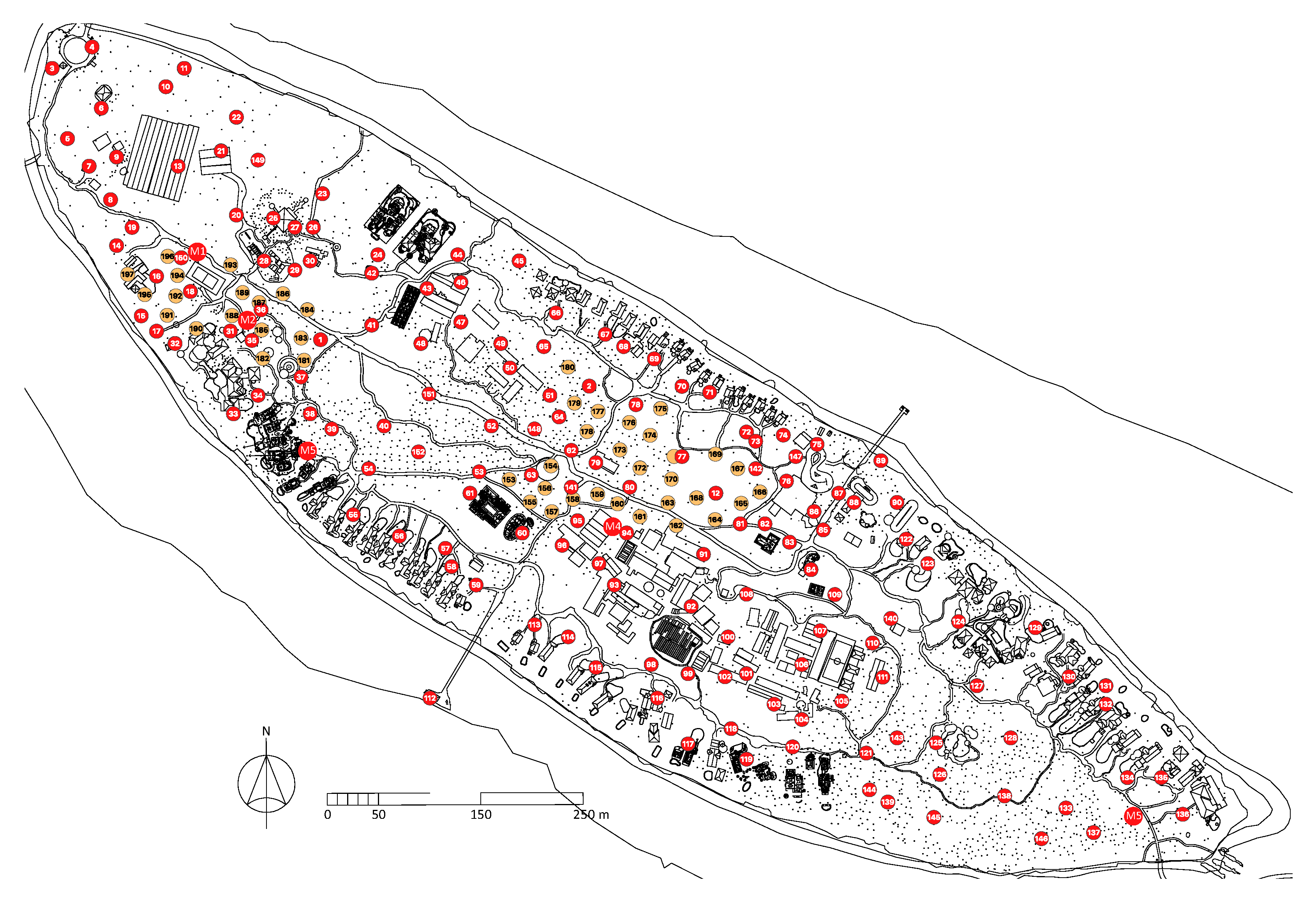
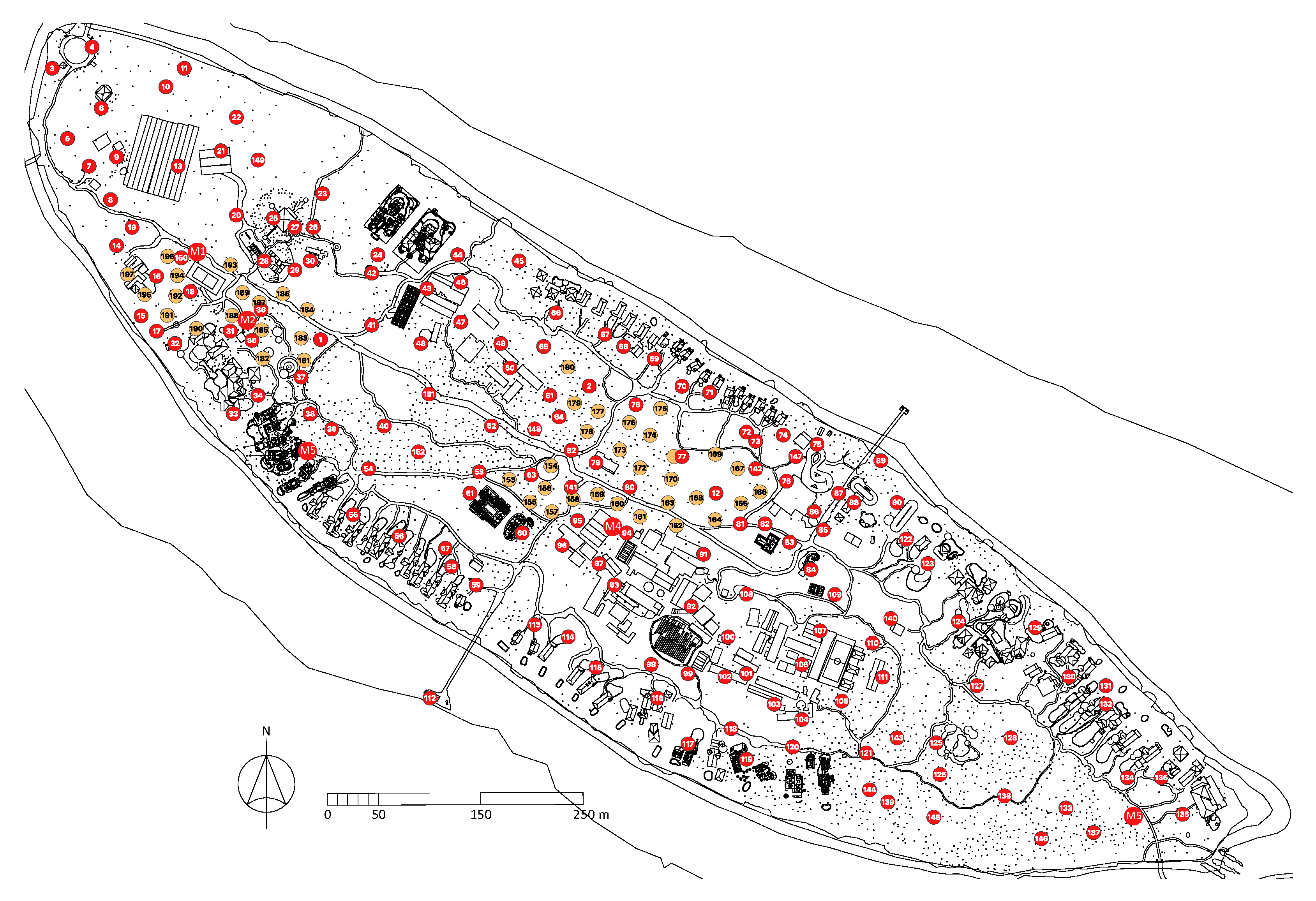
2.1. Study Island #1: Kunfunadhoo
2.2. Study Island #2: Medhufaru
2.3. Study Island #3: Thahigandu Kolhu
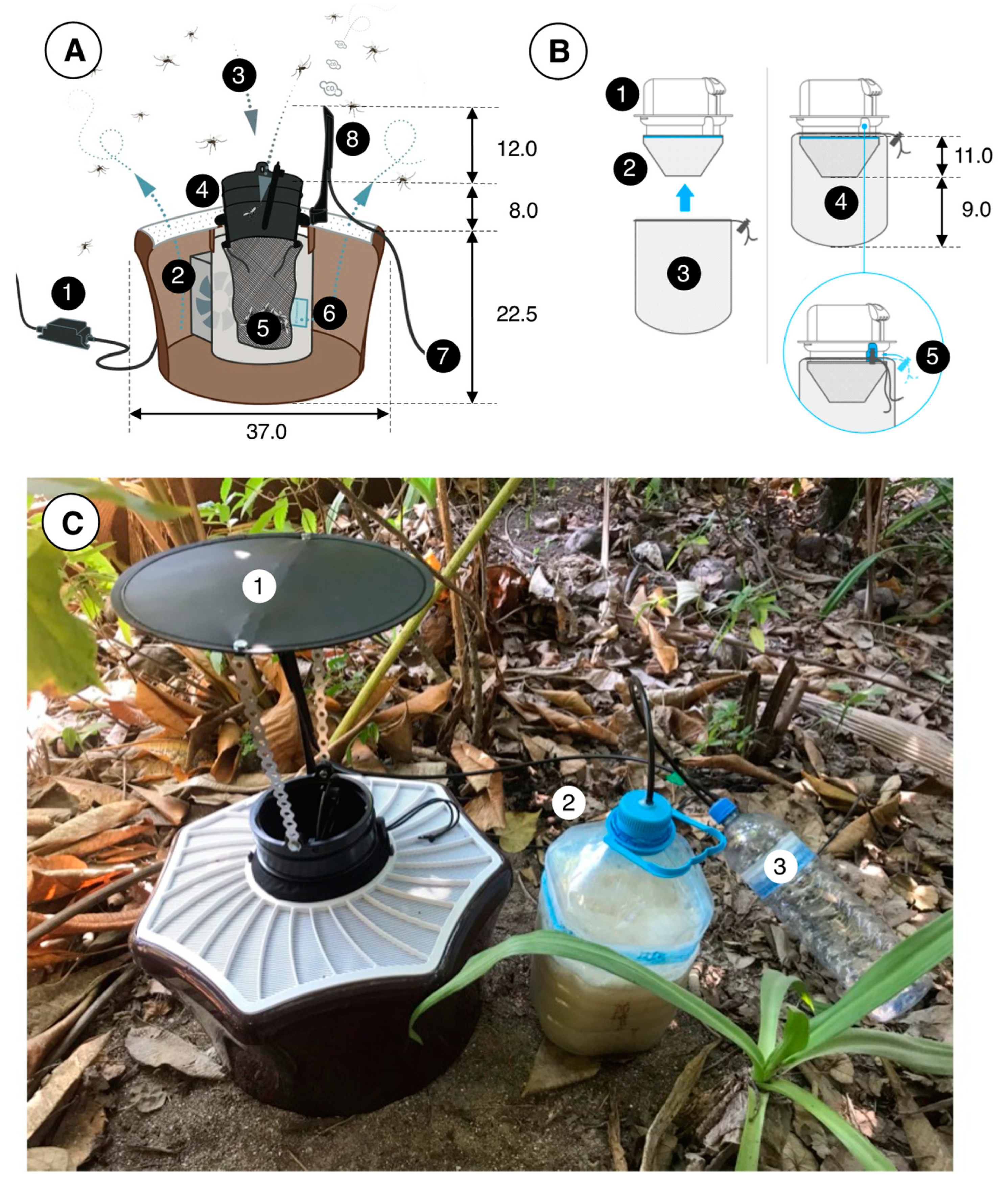
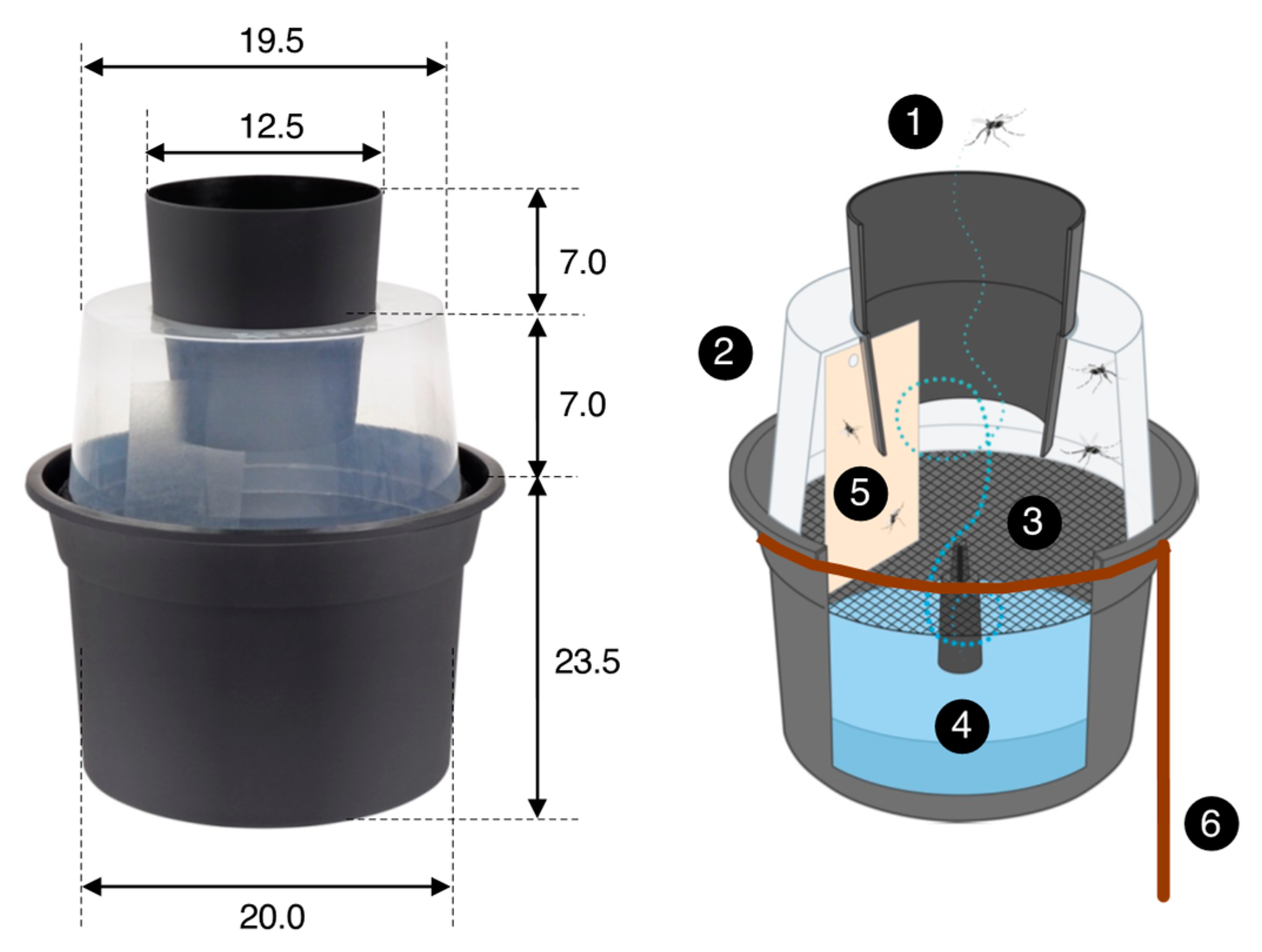
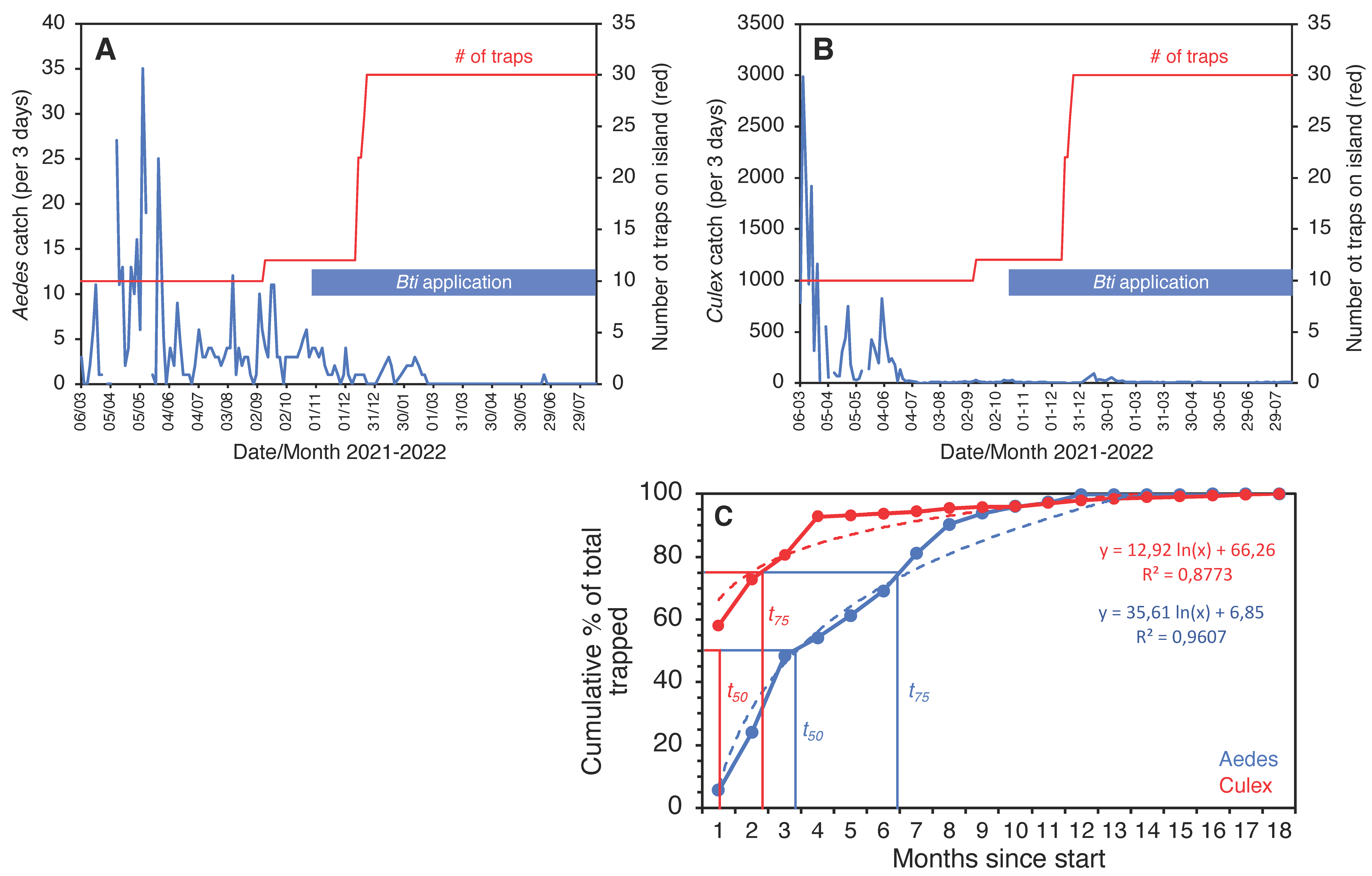
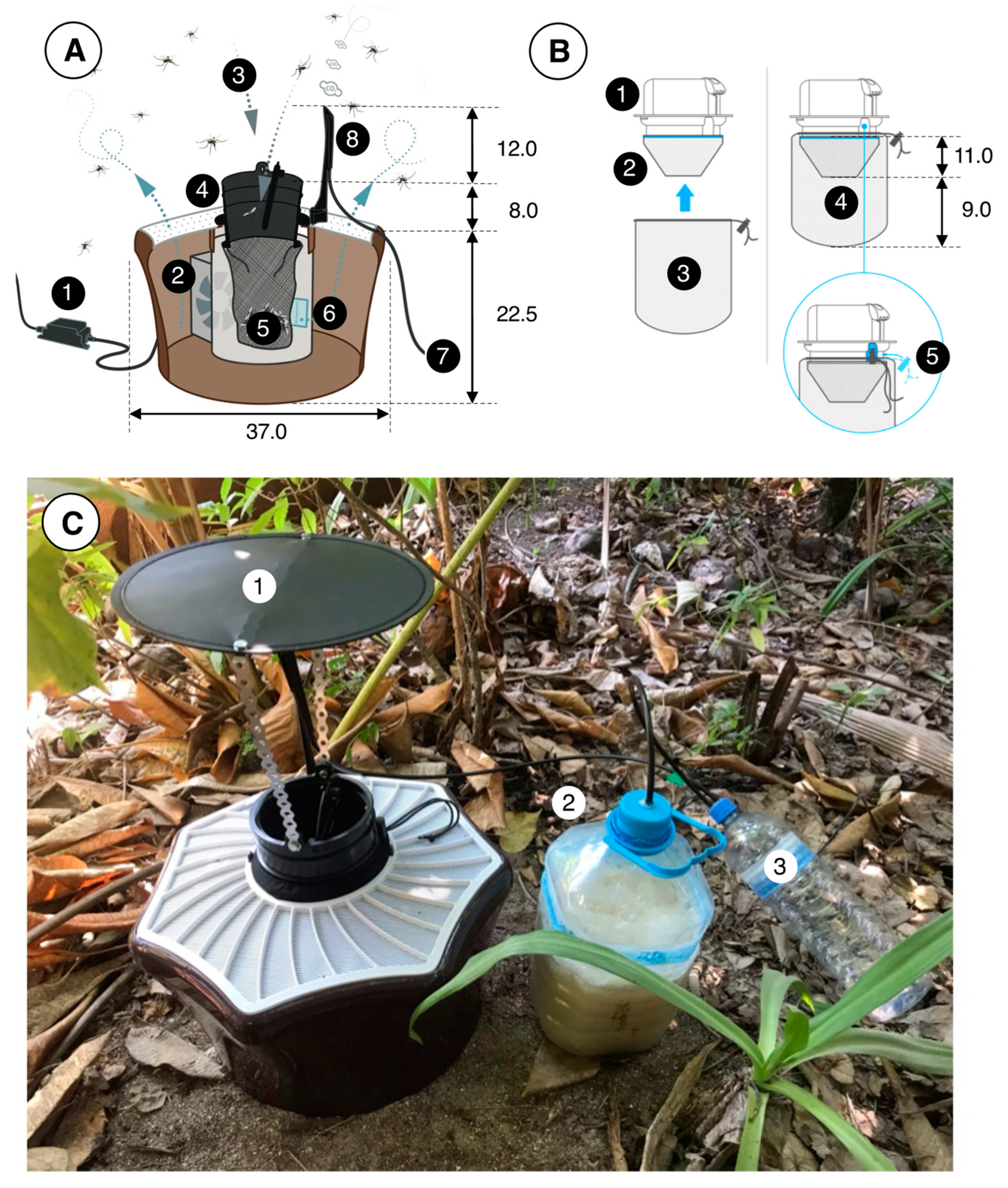
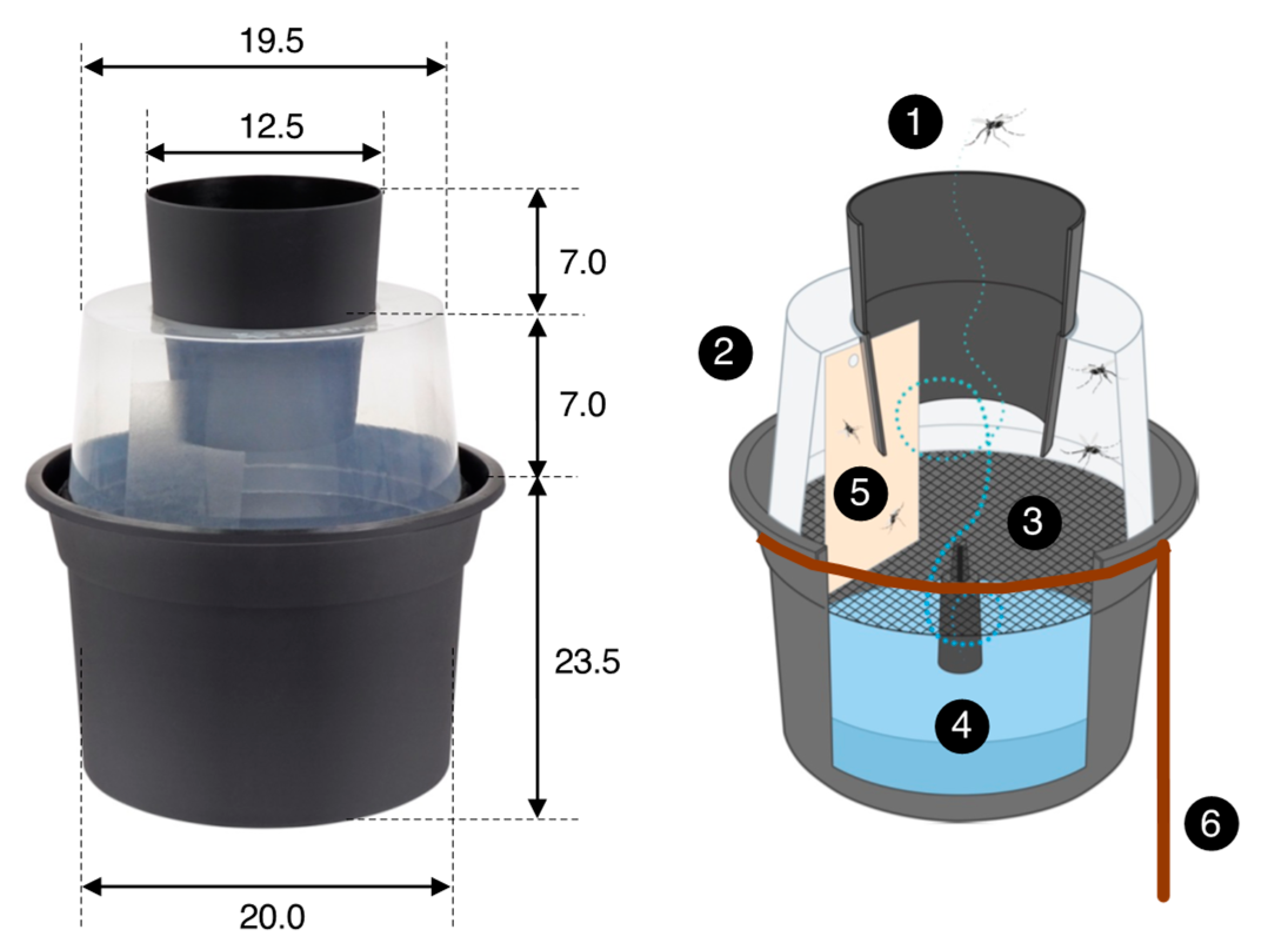
2.4. Mosquito Traps and Odor Baits
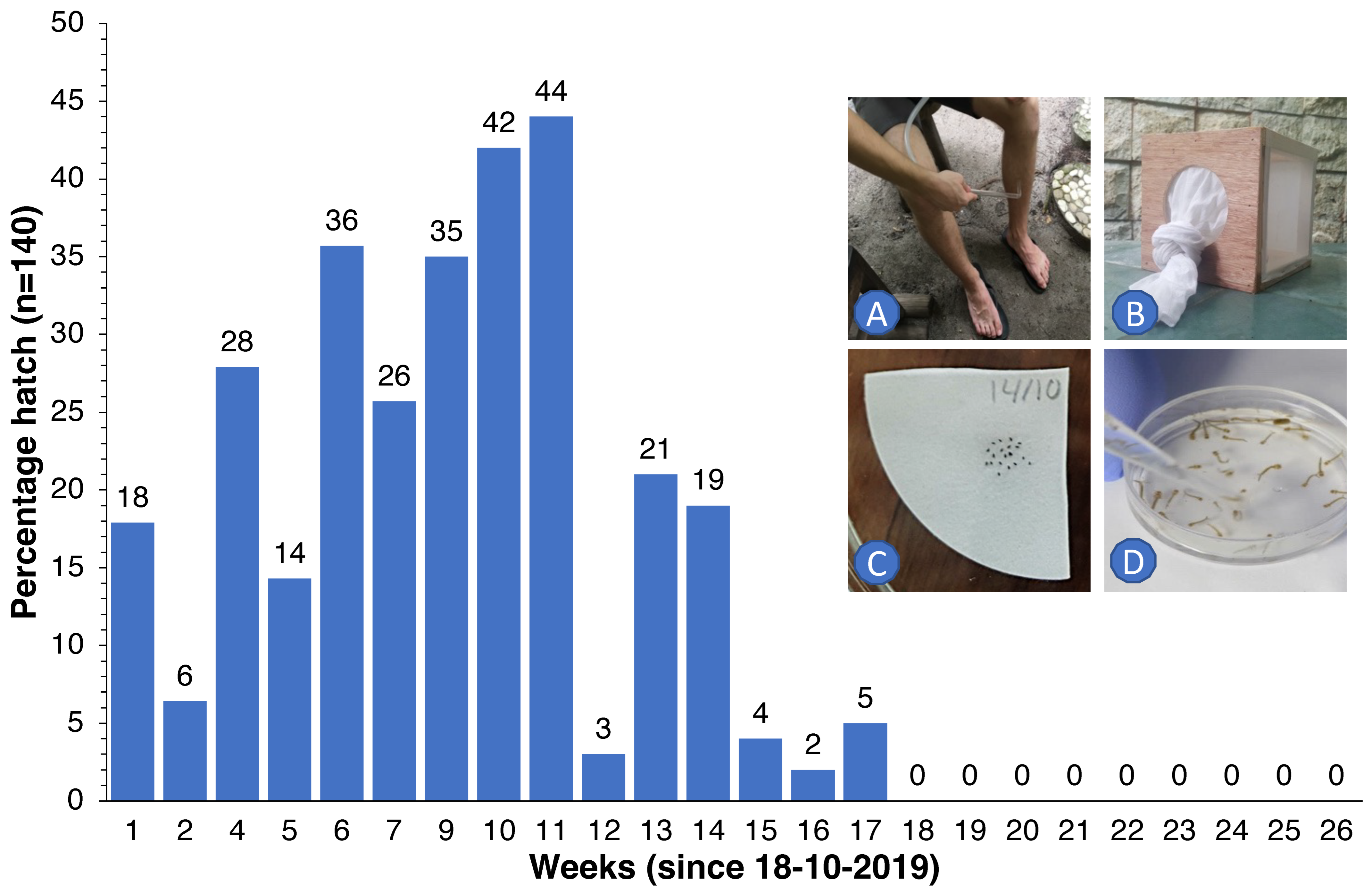
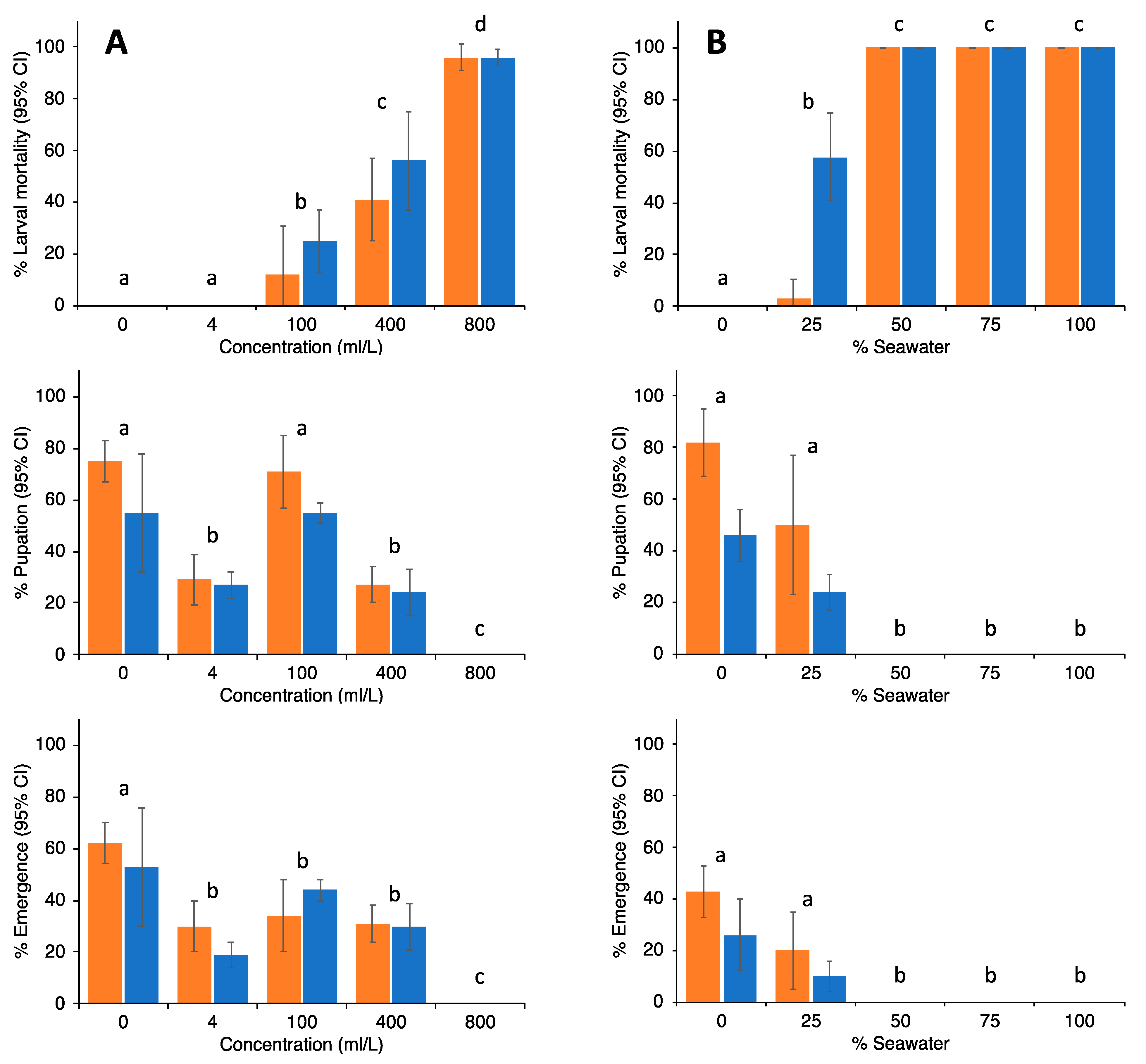
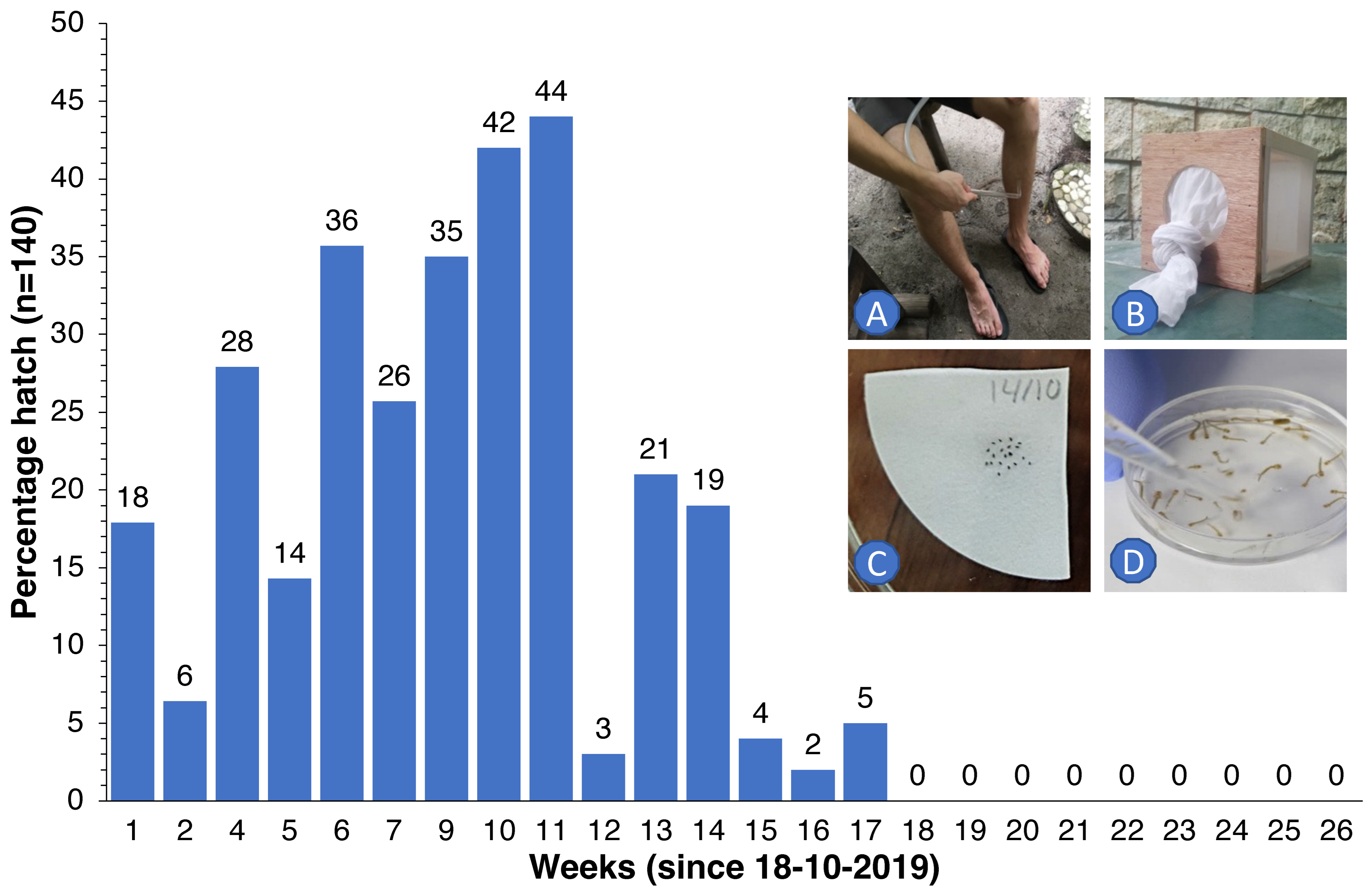
2.5. Egg Survival Experiments
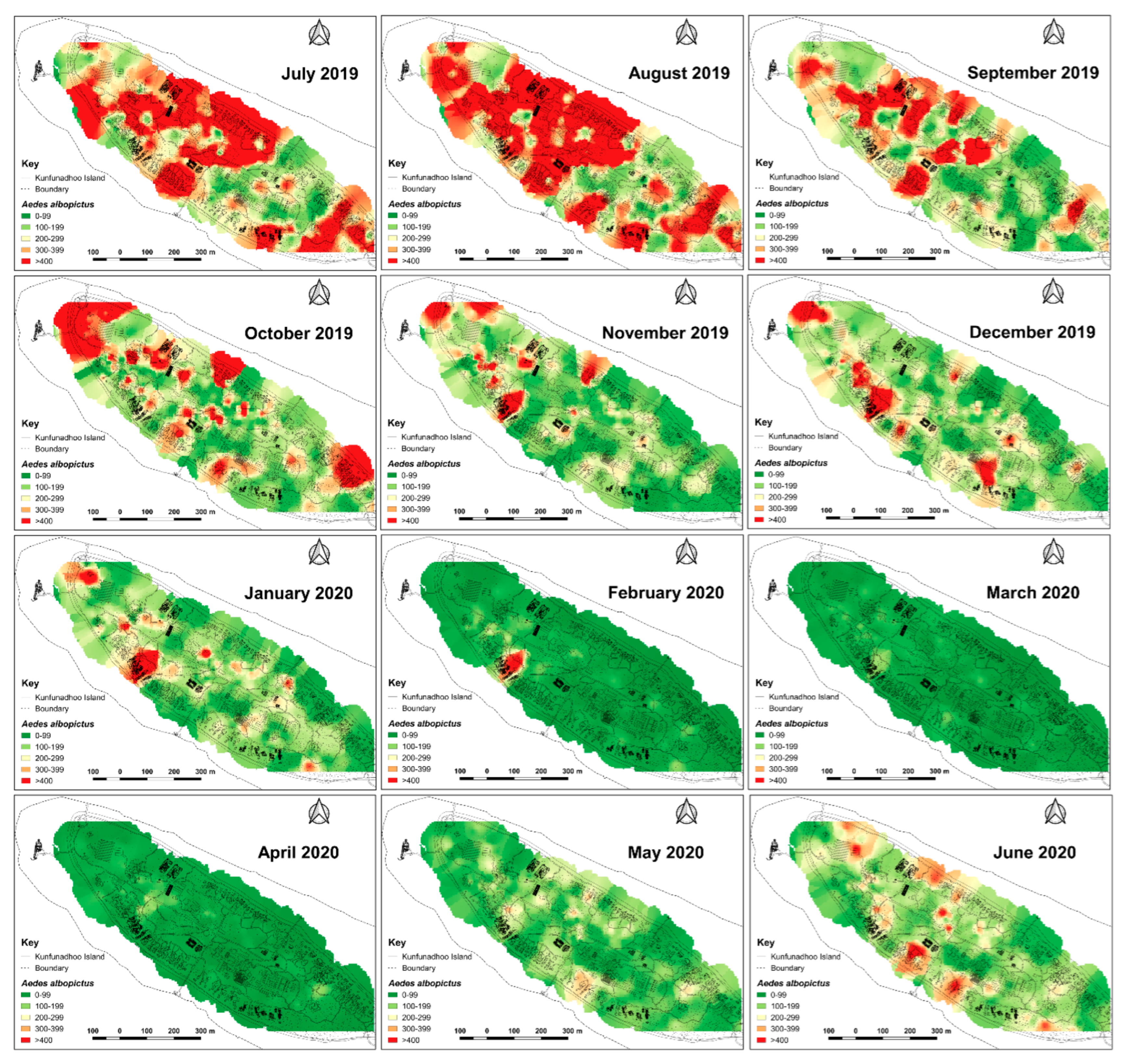
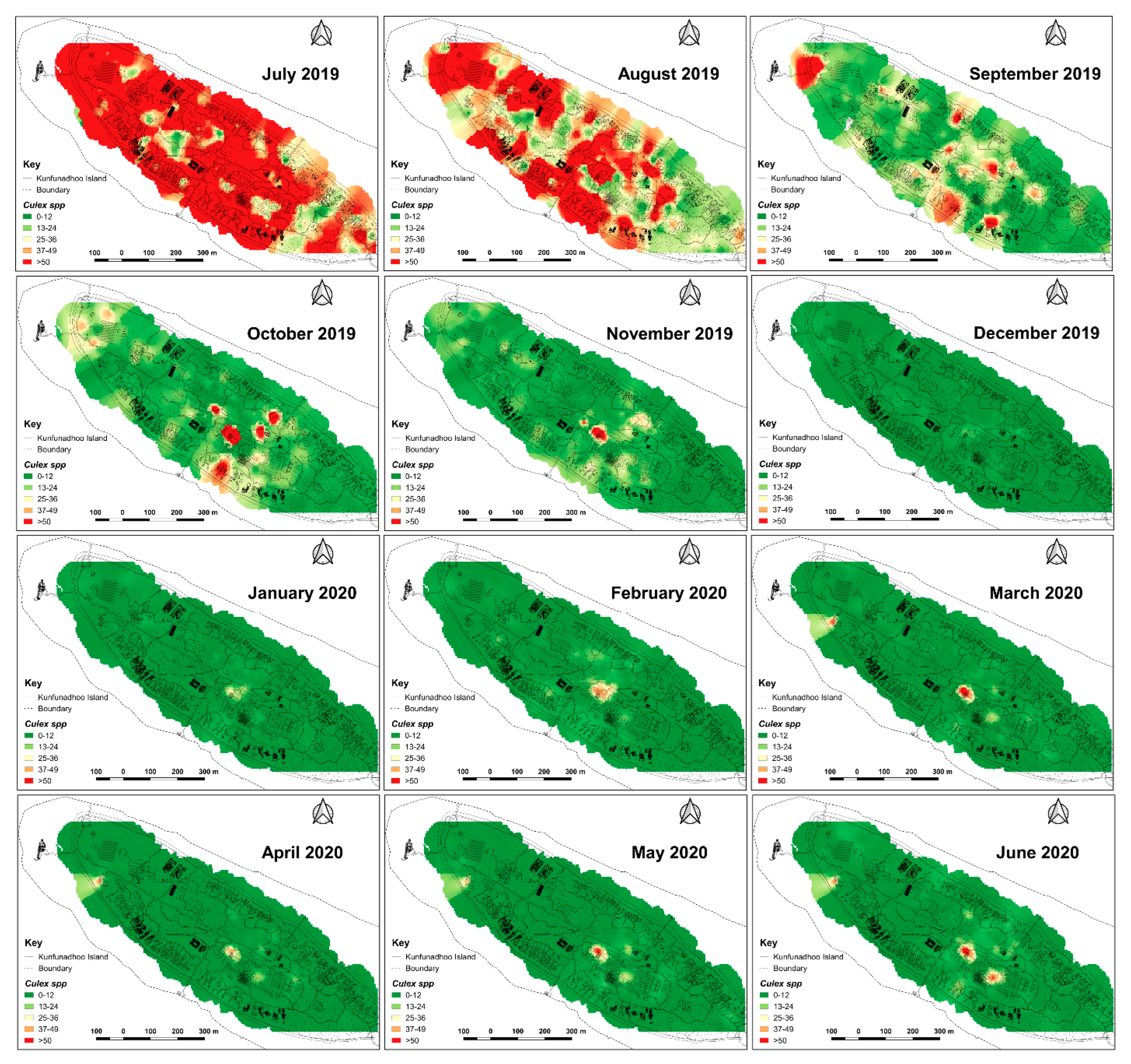
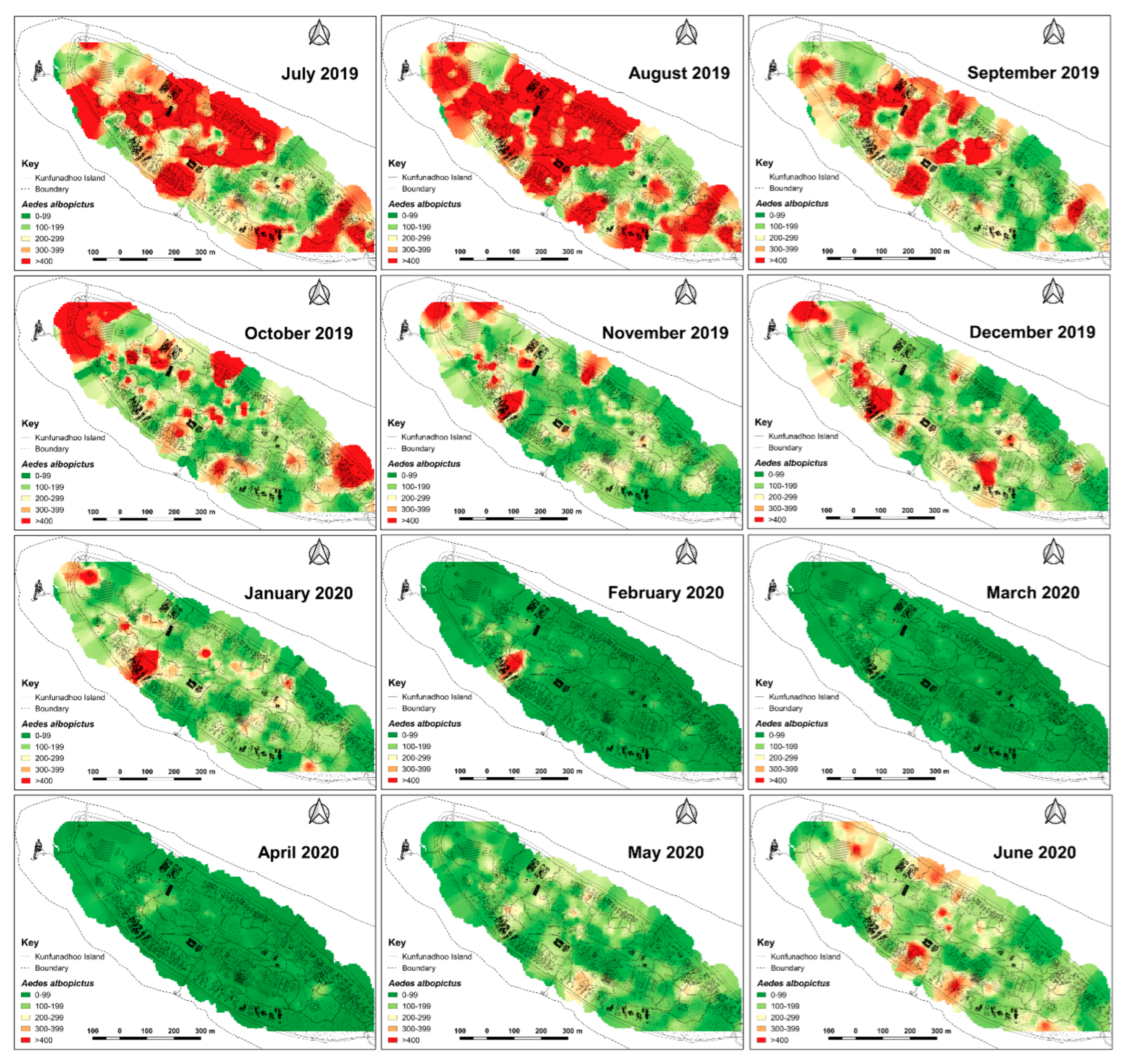
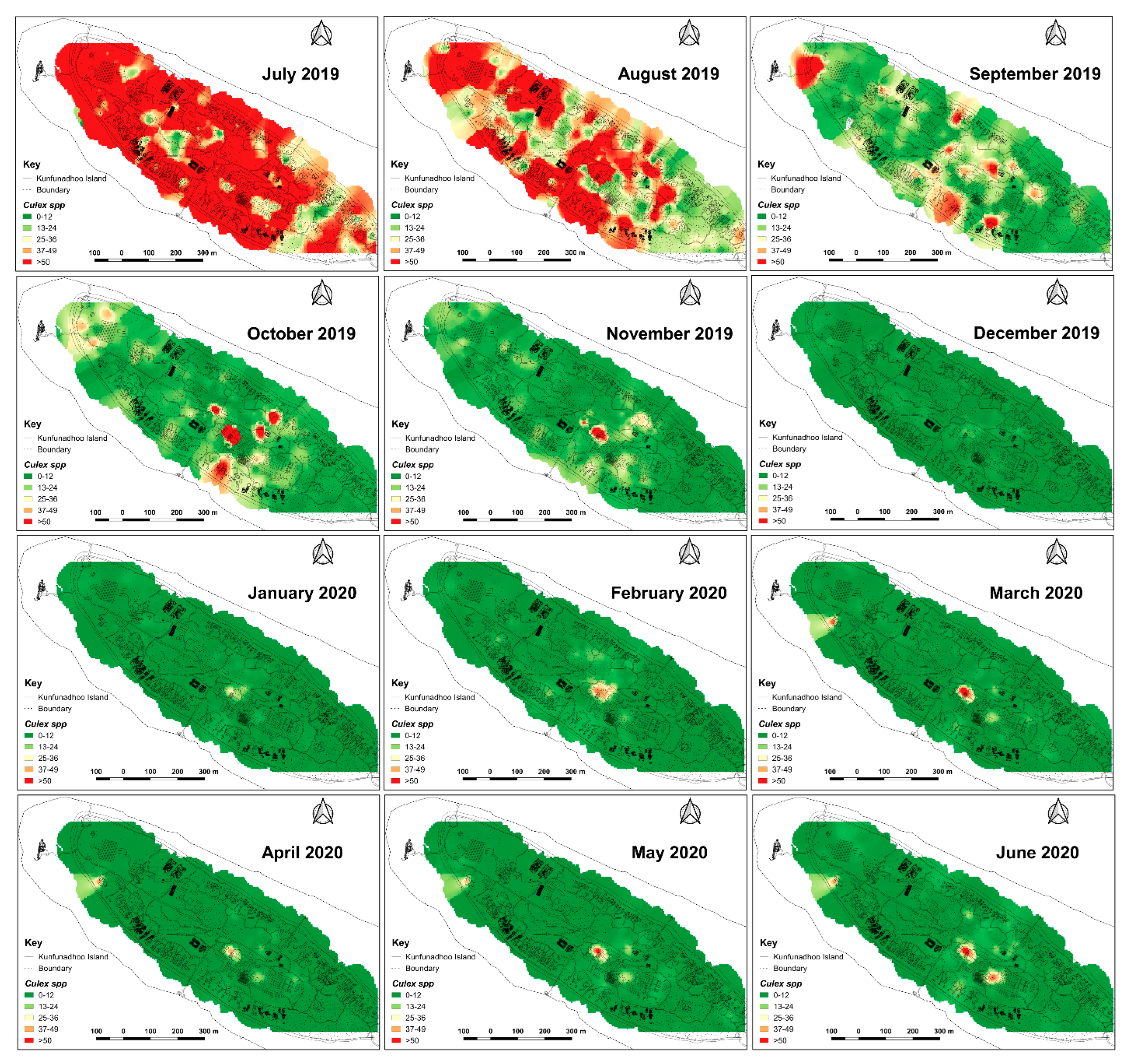
2.6. Heatmaps
2.7. Larval Source Management
2.8. Operations and Direct Cost Analysis
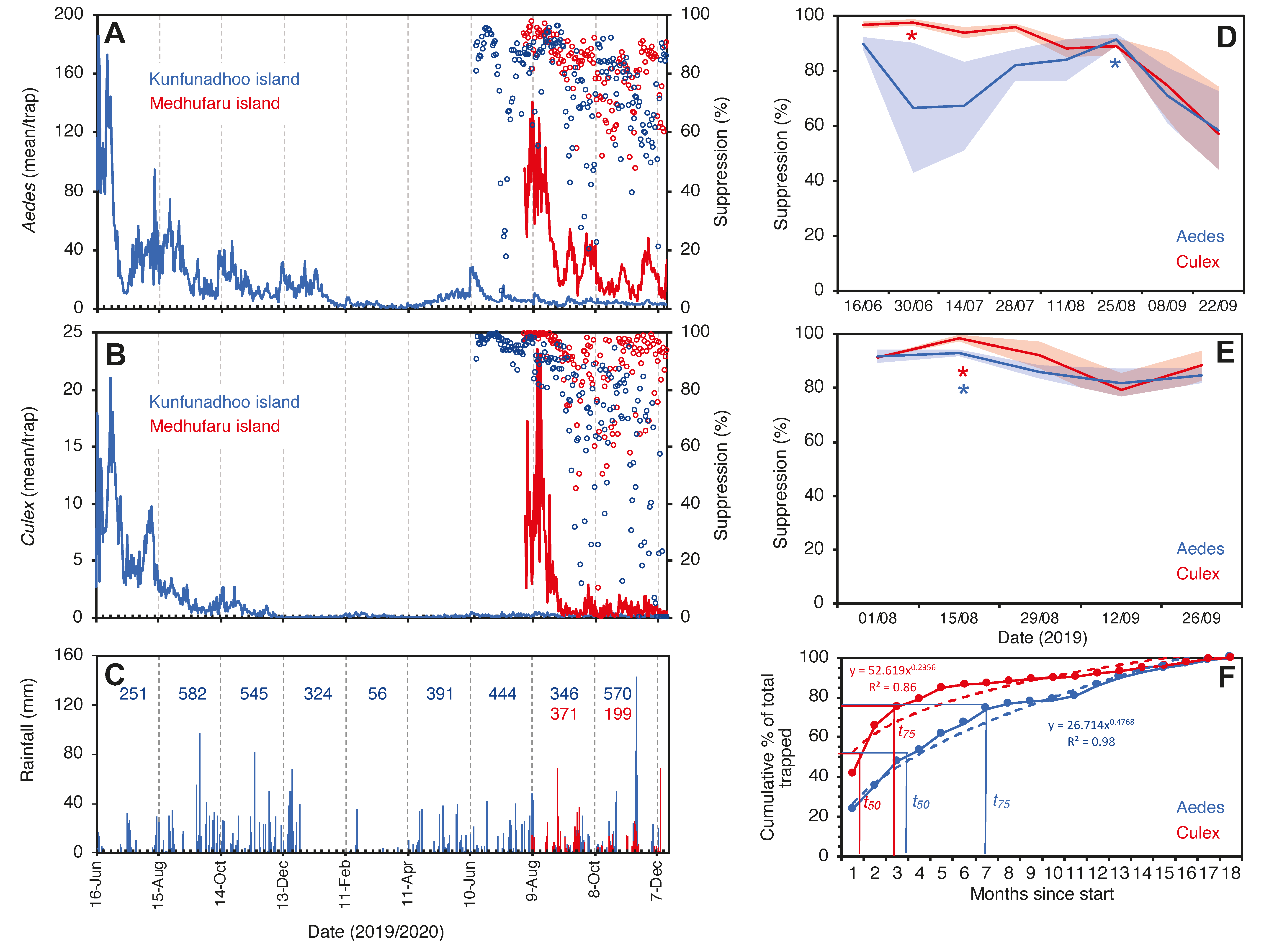
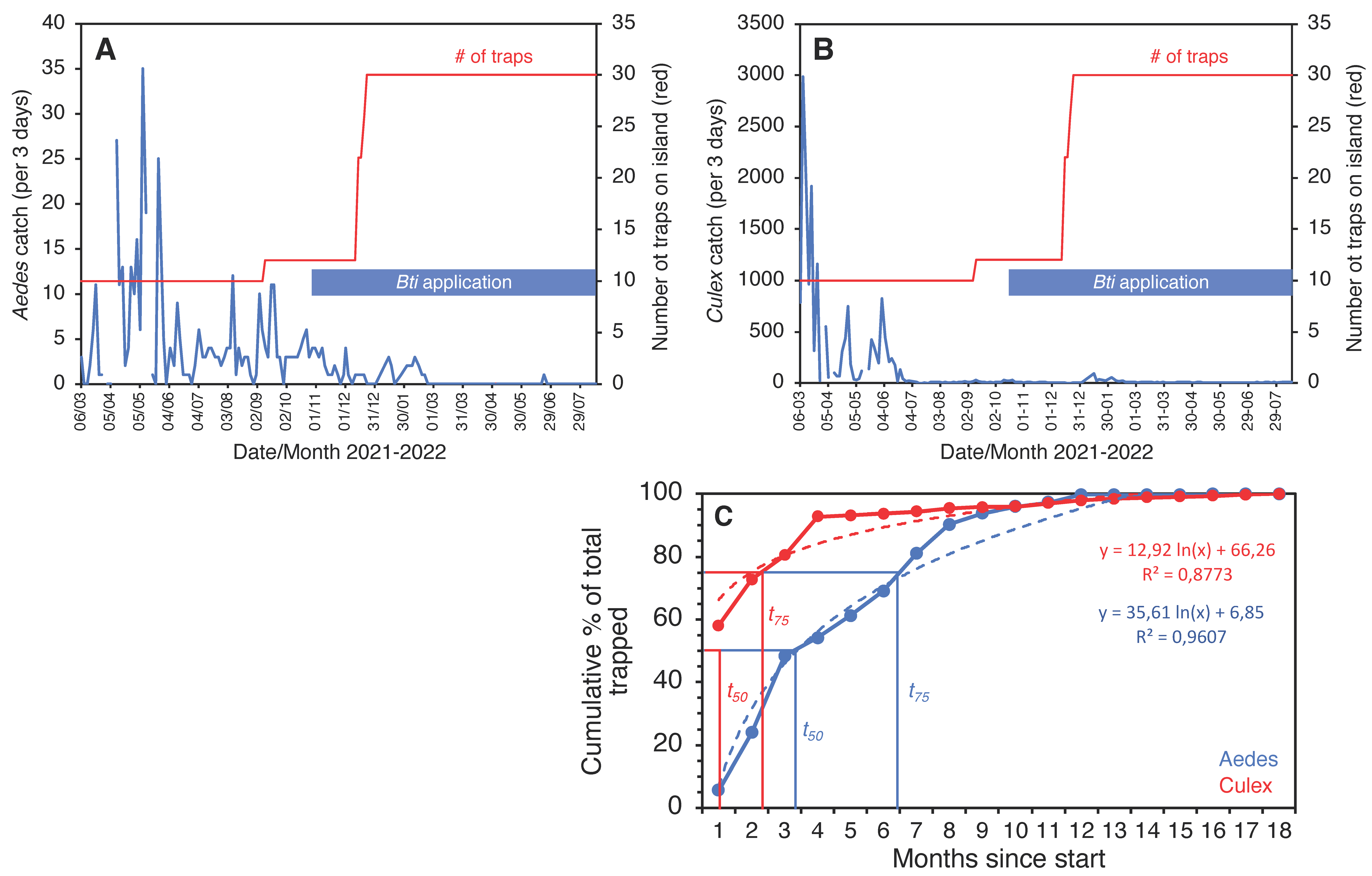
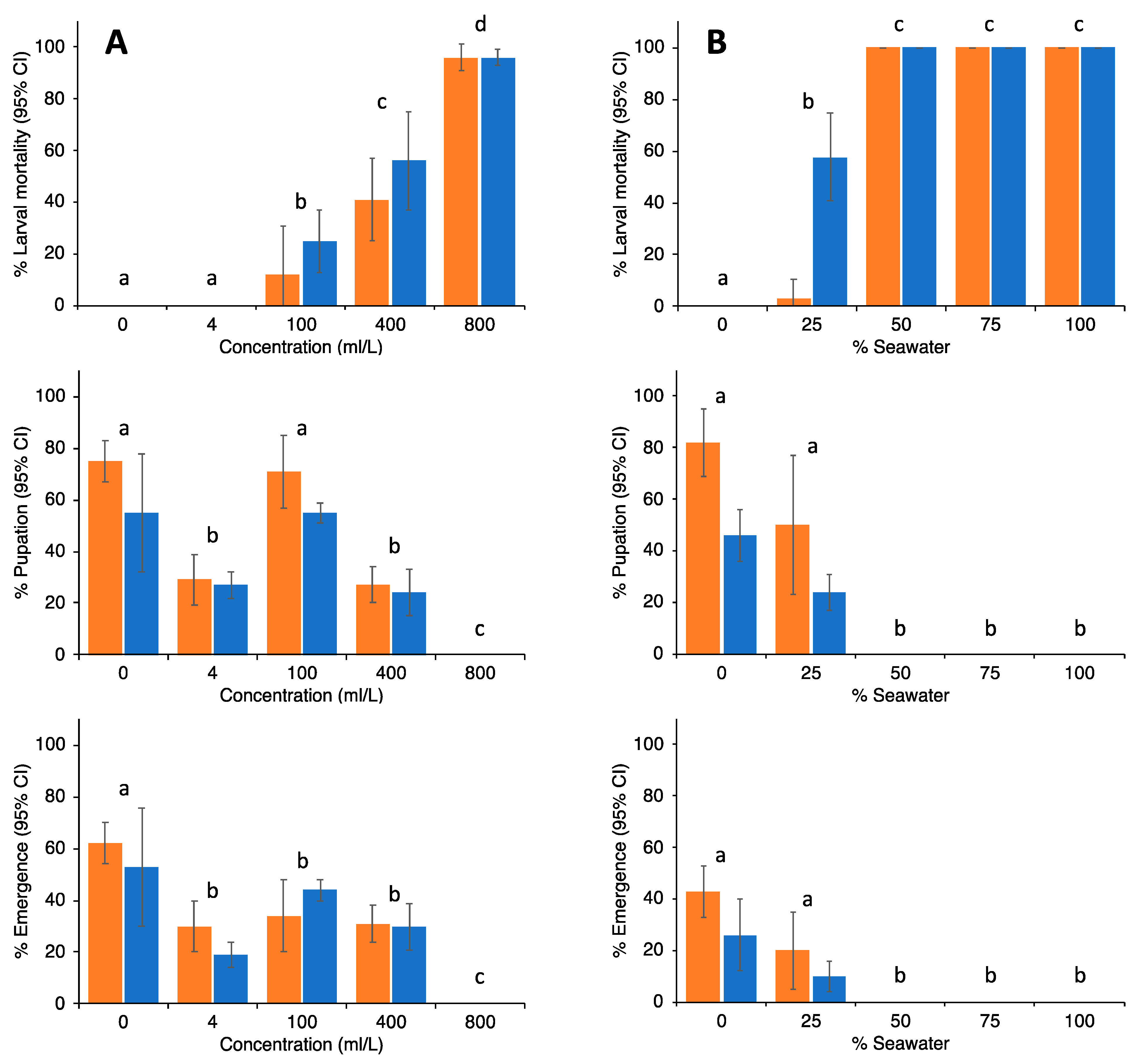
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-Ng, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C.; et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar] [CrossRef]
- Shaw, W.R.; Catteruccia, F. Vector biology meets disease control: Using basic research to fight vector-borne diseases. Nat. Microbiol. 2019, 4, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and sterile insect techniques combined eliminate mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S.; et al. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables large-scale suppression of wild populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef]
- Caputo, B.; Ienco, A.; Cianci, D.; Pombi, M.; Petrarca, V.; Baseggio, A.; Devine, G.J.; Della Torre, A. The “auto-dissemination” approach: A novel concept to fight Aedes albopictus in urban areas. PLoS Negl. Trop. Dis. 2012, 6, e1793. [Google Scholar] [CrossRef] [PubMed]
- Unlu, I.; Rochlin, I.; Suman, D.S.; Wang, Y.; Chandel, K.; Gaugler, R. Large-scale operational pyriproxyfen autodissemination deployment to suppress the immature Asian Tiger Mosquito (Diptera: Culicidae) populations. J. Med. Entomol. 2020, 57, 1120–1130. [Google Scholar] [CrossRef]
- Homan, T.; Hiscox, A.; Mweresa, C.K.; Masiga, D.; Mukabana, W.R.; Oria, P.; Maire, M.; Aurelio, D.P.; Silkey, M.; Alaii, J.; et al. The effect of mass mosquito trapping on malaria transmission and disease burden (SolarMal): A stepped-wedge cluster-randomised trial. Lancet 2016, 388, 1193–1201. [Google Scholar] [CrossRef]
- Martinet, J.-P.; Ferté, H.; Failloux, A.-B.; Schaffner, F.; Depaquit, J. Mosquitoes of North-Western Europe as potential vectors of arboviruses: A review. Viruses 2019, 11, 1059. [Google Scholar] [CrossRef] [Green Version]
- Pereira-dos-Santos, T.; Roiz, D.; Lourenço-de-Oliveira, R.; Paupy, C. A Systematic Review: Is Aedes albopictus an efficient bridge vector for zoonotic arboviruses? Pathogens 2020, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Maldives Ministry of Tourism. Tourism Yearbook 2021. Available online: https://www.tourism.gov.mv/dms/document/2f11c02edec48b0fa37014122e7c39e6.pdf (accessed on 18 August 2022).
- World Health Organization (WHO). Malaria-Free Maldives (WHO, 2016). Available online: http://apps.who.int/iris/bitstream/10665/250397/1/9789290225140-eng.pdf (accessed on 17 August 2022).
- World Health Organization (WHO). Finishing off Lymphatic Filariasis: How Maldives Eliminated a Once-Prominent Scourge (WHO, 2017). Available online: https://www.who.int/southeastasia/news/feature-stories/detail/finishing-off-lymphatic-filariasis-how-maldives-eliminated-a-once-prominent-scourge (accessed on 17 August 2022).
- Abdulla, A.A.; Rasheeda, F.; Ahmed, I.N.; Aboobakur, M. An evaluation of the surveillance system for dengue virus infections in Maldives. WHO South-East Asia J. Public Health 2014, 3, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Yoosuf, A.A.; Shiham, I.; Mohamed, A.J.; Ali, G.; Luna, J.M.; Pandav, R.; Gongal, G.N.; Nisaluk, A.; Jarman, R.G.; Gibbons, R.V. First report of chikungunya from the Maldives. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, E.M.; Huhtamo, E.; Smura, T.; Kallio-Kokko, H.; Raassina, M.; Vapalahti, O. Zika virus infection in a traveller returning from the Maldives, June 2015. Eurosurveillance 2016, 21, 30107. [Google Scholar] [CrossRef]
- Bangert, M.; Latheef, A.T.; Pant, S.D.; Ahmed, I.N.; Saleem, S.; Rafeeq, F.N.; Abdulla, M.; Shamah, F.; Jamsheed Mohamed, A.; Fitzpatrick, C.; et al. Economic analysis of dengue prevention and case management in the Maldives. PLoS Negl. Trop. Dis. 2018, 12, e0006796. [Google Scholar] [CrossRef]
- Biogents, A.G. Product Catalogue. Available online: https://eu.biogents.com/wp-content/uploads/Biogents-Product-Catalog-en.pdf (accessed on 18 August 2022).
- Smallegange, R.C.; Schmied, W.H.; van Roey, K.J.; Verhulst, N.O.; Spitzen, J.; Mukabana, W.R.; Takken, W. Sugar-fermenting yeast as an organic source of carbon dioxide to attract the malaria mosquito Anopheles gambiae. Malar J. 2010, 9, 292. [Google Scholar] [CrossRef]
- Aldridge, R.L.; Britch, S.C.; Allan, S.A.; Tsikolia, M.; Carolina Calix, M.; Bernier, U.R.; Linthicum, K.J. Comparison of volatiles and mosquito capture efficacy for three carbohydrate sources in a yeast-fermentation CO2 generator. J. Am. Mosq. Control Assoc. 2016, 32, 282–291. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, J.; Li, T.; Liu, Q.; Gong, Z.; Hou, J. Effect of different carbon dioxide (CO2) flows on trapping Aedes albopictus with BG traps in the field in Zhejiang Province, China. PLoS ONE 2020, 15, e0243061. [Google Scholar] [CrossRef] [PubMed]
- Acree, F., Jr.; Turner, R.B.; Gouck, H.K.; Beroza, M.; Smith, N. L-Lactic acid: A mosquito attractant isolated from humans. Science 1968, 161, 1346–1347. [Google Scholar] [CrossRef]
- Allan, S.A.; Bernier, U.R.; Kline, D.L. Laboratory evaluation of lactic acid on attraction of Culex spp. (Diptera: Culicidae). J. Vector Ecol. 2010, 35, 318–324. [Google Scholar] [CrossRef]
- Eiras, A.E.; Buhagiar, T.S.; Ritchie, S.A. Development of the Gravid Aedes Trap for the capture of adult female container–exploiting mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2014, 51, 200–209. [Google Scholar] [CrossRef]
- Harwood, J.F.; Rama, V.; Hash, J.M.; Gordon, S.W. The attractiveness of the Gravid Aedes Trap to dengue vectors in Fiji. J. Med. Entomol. 2017, 55, 481–484. [Google Scholar] [CrossRef]
- Mendki, M.J.; Ganesan, K.; Prakash, S.; Suryanarayana, M.V.S.; Malhotra, R.C.; Rao, K.M.; Vaidyanathaswamy, R. Heneicosane: An oviposition-attractant pheromone of larval origin in Aedes aegypti mosquito. Curr. Sci. 2000, 78, 1295–1296. [Google Scholar]
- Seenivasagan, T.; Sharma, K.R.; Sekhar, K.; Ganesan, K.; Prakash, S.; Vijayaraghavan, R. Electroantennogram, flight orientation, and oviposition responses of Aedes aegypti to the oviposition pheromone n-heneicosane. Parasitol. Res. 2009, 104, 827–833. [Google Scholar] [CrossRef]
- Denlinger, D.L. Dormancy in tropical insects. Annu. Rev. Entomol. 1986, 31, 239–264. [Google Scholar] [CrossRef]
- Sota, T. Response to selection for desiccation resistance in Aedes albopictus eggs (Diptera: Culicidae). Appl. Entomol. Zool. 1993, 28, 161–168. [Google Scholar] [CrossRef]
- World Health Organization. Larval Source Management: A Supplementary Measure for Malaria Vector Control. An Operational Manual. World Health Organization: Geneva, Switzerland, 2013; Available online: https://apps.who.int/iris/bitstream/handle/10665/85379/9789241505604_eng.pdf (accessed on 17 August 2022).
- Okumu, F.O.; Knols, B.G.J.; Fillinger, U. Larvicidal effects of a neem (Azadirachta indica) oil formulation on the malaria vector Anopheles gambiae. Malar. J. 2007, 6, 63. [Google Scholar] [CrossRef]
- Zheng, M.L.; Zhang, D.J.; Damiens, D.D.; Lees, R.S.; Gilles, J.R.L. Standard operating procedures for standardized mass rearing of the dengue and chikungunya vectors Aedes aegypti and Aedes albopictus (Diptera: Culicidae)-II-Egg storage and hatching. Parasit. Vectors 2015, 26, 348. [Google Scholar] [CrossRef]
- Amos, B.A.; Ritchie, S.A.; Cardé, R.T. Attraction versus capture: Efficiency of BG-Sentinel trap under semi-field conditions and characterizing response behaviors for female Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 884–892. [Google Scholar] [CrossRef]
- Akhoundi, M.; Jourdain, F.; Chandre, F.; Delaunay, P.; Roiz, D. Effectiveness of a field trap barrier system for controlling Aedes albopictus: A “removal trapping” strategy. Parasit. Vectors 2018, 11, 101. [Google Scholar] [CrossRef]
- Fang, J. A world without mosquitoes. Nature 2010, 466, 432–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, C.M.; Bonds, J.A.S.; Quinlan, M.M.; Mumford, J.D. Effects of the removal or reduction in density of the malaria mosquito, Anopheles gambiae s.l., on interacting predators and competitors in local ecosystems. Med. Vet. Entomol. 2019, 33, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef] [PubMed]
- Duarte, C.M.; Agusti, S.; Barbier, E.; Britten, G.L.; Castilla, J.C.; Gattuso, J.P.; Fulweiler, R.W.; Hughes, T.P.; Knowlton, N.; Lovelock, C.E.; et al. Rebuilding marine life. Nature 2020, 580, 39–51. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahir, A.; Kahamba, N.F.; Knols, T.O.; Jackson, G.; Patty, N.F.A.; Shivdasani, S.; Okumu, F.O.; Knols, B.G.J. Mass Trapping and Larval Source Management for Mosquito Elimination on Small Maldivian Islands. Insects 2022, 13, 805. https://doi.org/10.3390/insects13090805
Jahir A, Kahamba NF, Knols TO, Jackson G, Patty NFA, Shivdasani S, Okumu FO, Knols BGJ. Mass Trapping and Larval Source Management for Mosquito Elimination on Small Maldivian Islands. Insects. 2022; 13(9):805. https://doi.org/10.3390/insects13090805
Chicago/Turabian StyleJahir, Akib, Najat F. Kahamba, Tom O. Knols, Gordon Jackson, Nila F. A. Patty, Sonu Shivdasani, Fredros O. Okumu, and Bart G. J. Knols. 2022. "Mass Trapping and Larval Source Management for Mosquito Elimination on Small Maldivian Islands" Insects 13, no. 9: 805. https://doi.org/10.3390/insects13090805
APA StyleJahir, A., Kahamba, N. F., Knols, T. O., Jackson, G., Patty, N. F. A., Shivdasani, S., Okumu, F. O., & Knols, B. G. J. (2022). Mass Trapping and Larval Source Management for Mosquito Elimination on Small Maldivian Islands. Insects, 13(9), 805. https://doi.org/10.3390/insects13090805