
Fitness of the Papaya Mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae), after Transferring from Solanum tuberosum to Carica papaya, Ipomoea batatas, and Alternanthera philoxeroides


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Cultivation of Host Plants
2.2. Paracoccus marginatus
2.3. Life Table Study of P. marginatus
2.4. Life Table Data Analysis
2.5. Population Projection
3. Results
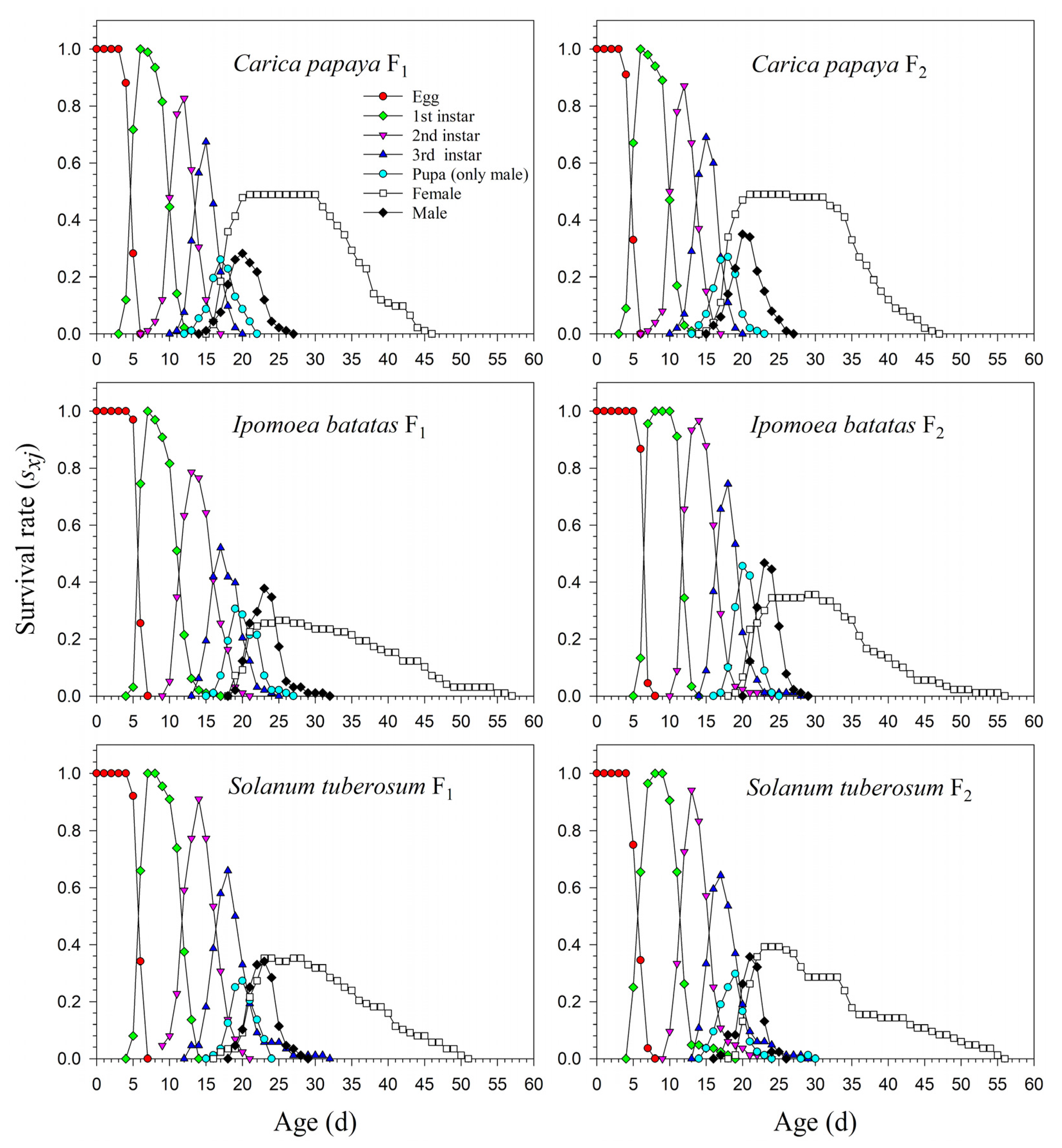
3.1. Development and Fecundity of P. marginatus after Host Plant Shifting
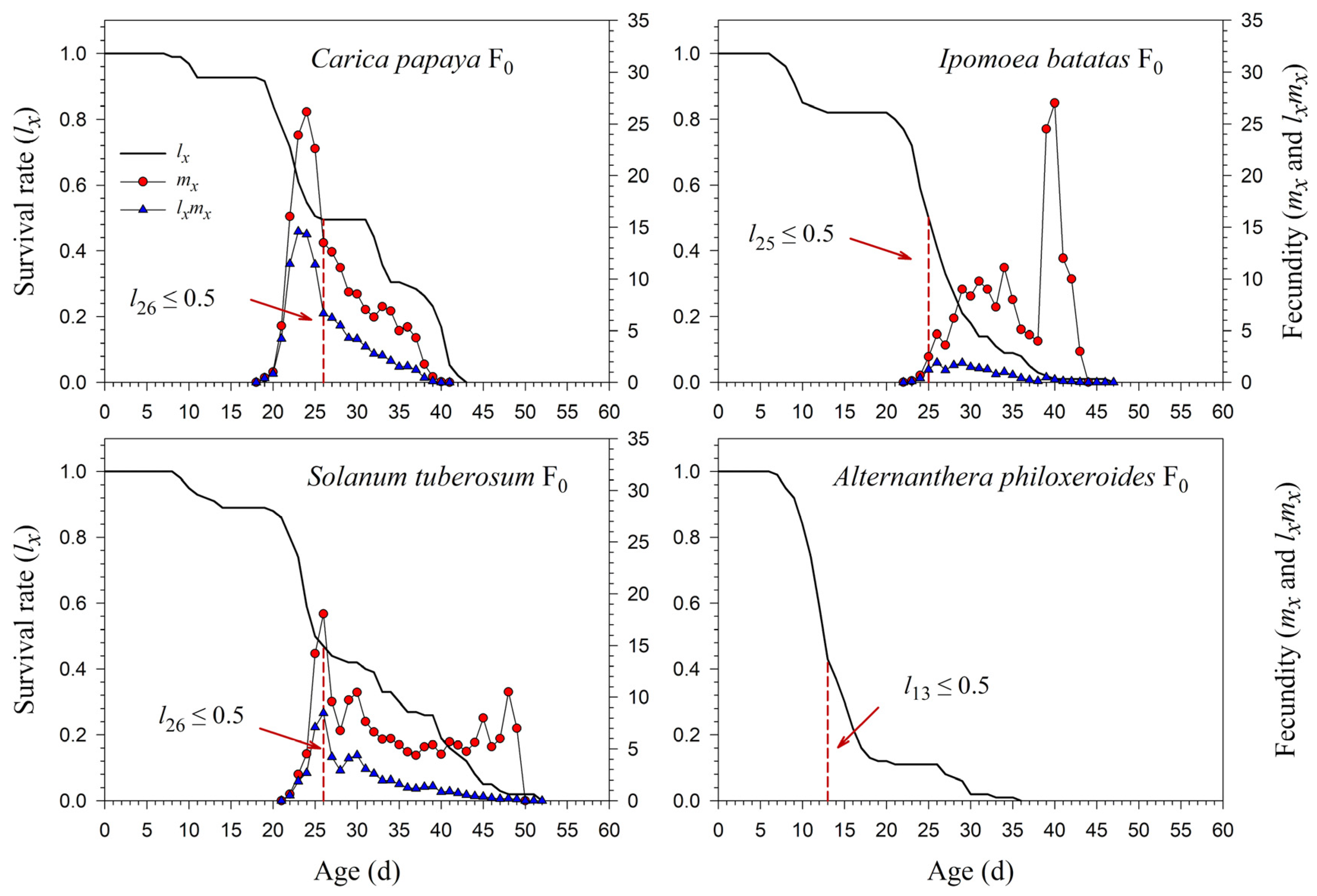
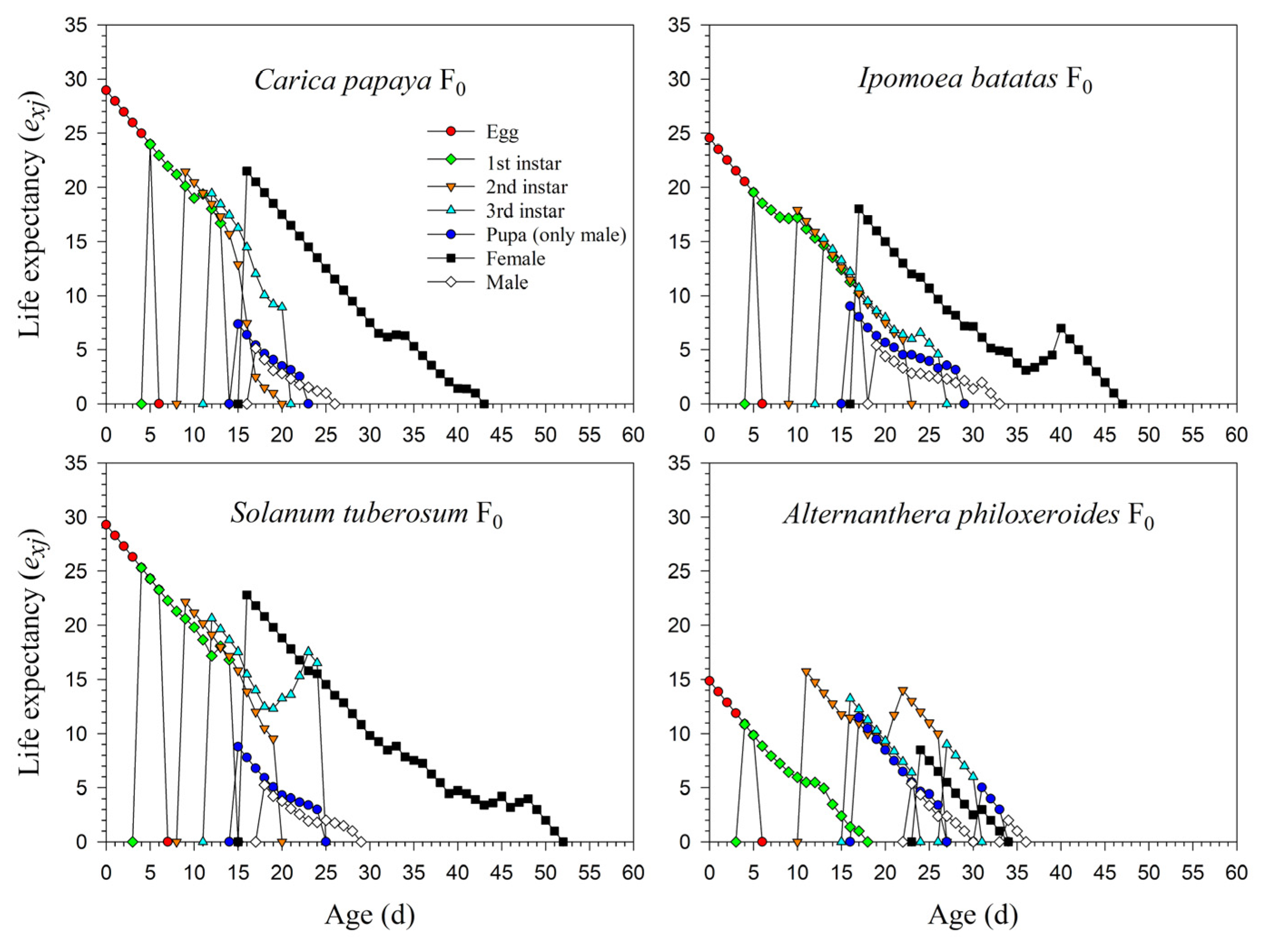
3.2. Population Parameters of P. marginatus after Host Plant Shifting
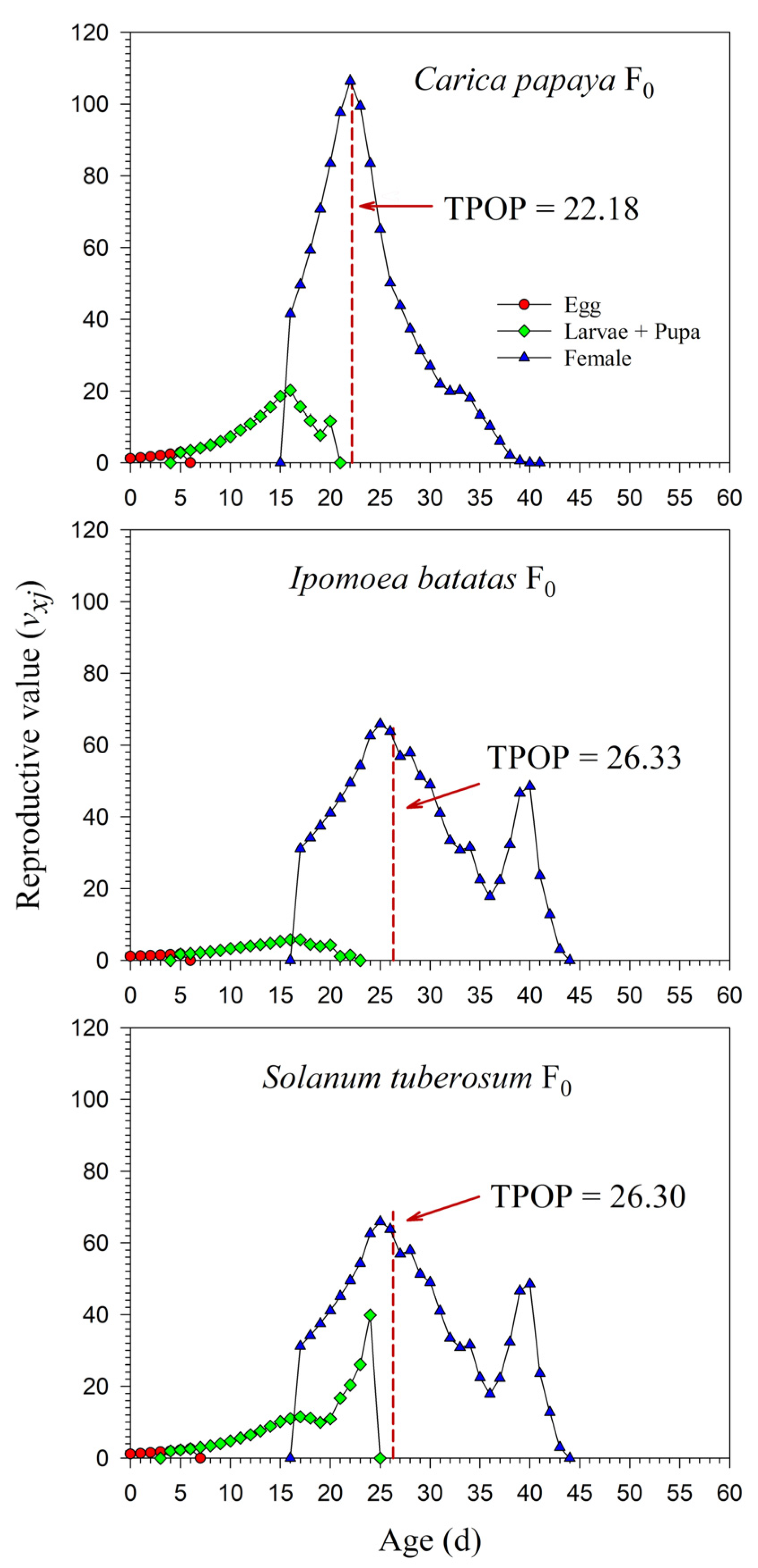
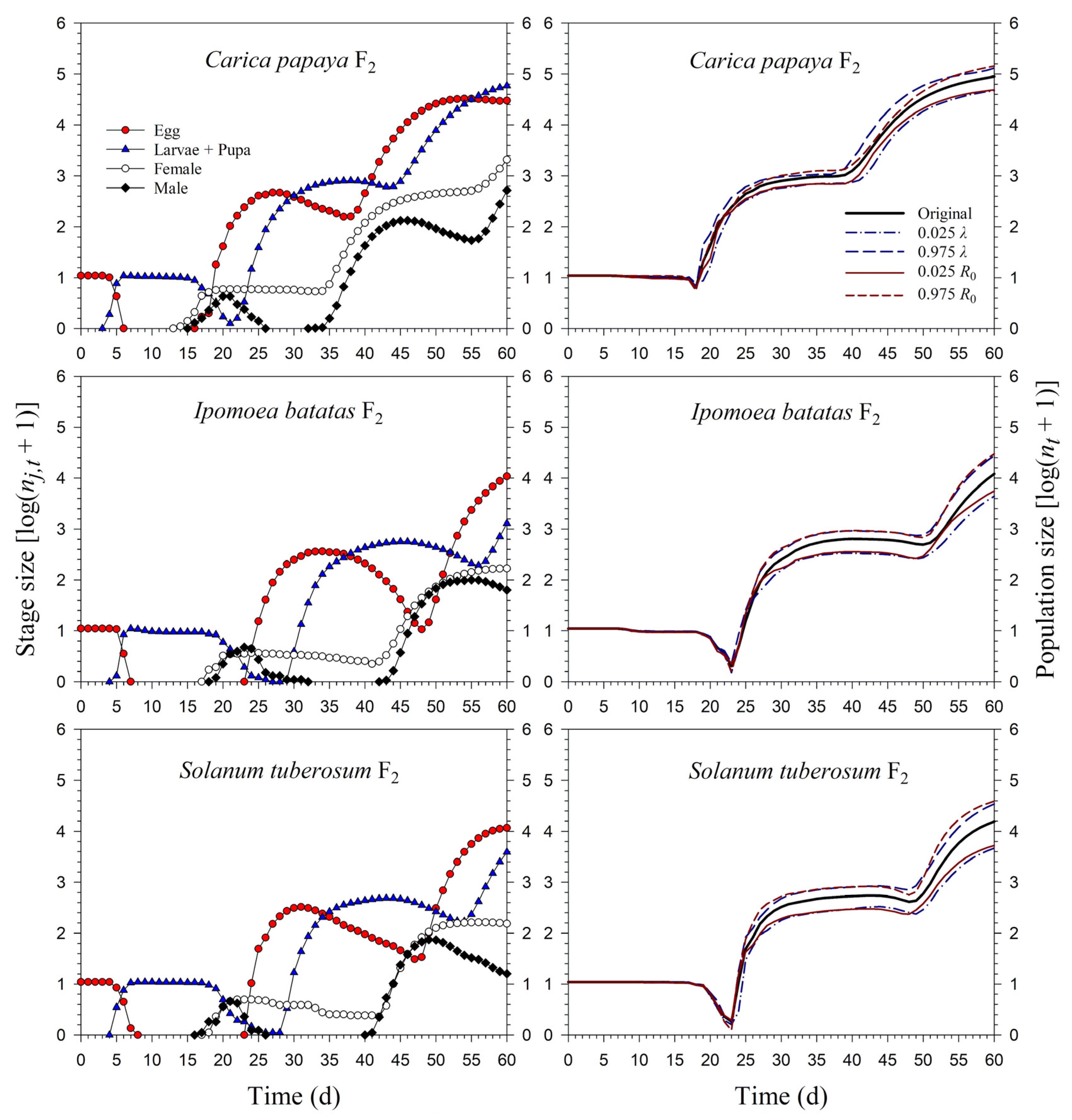
3.3. Population Projection of P. marginatus
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, F.Z.; Liu, Z.H.; Shen, H.; Yu, F.; Ma, J.; Hu, X.N.; Zeng, L. Morphological and molecular identification of Paracoccus marginatus (Hemiptera: Pseudococcidae) in Yunnan, China. Fla. Entomol. 2014, 97, 1469–1473. [Google Scholar] [CrossRef]
- Miller, D.R.; Williams, D.J. Notes on a new mealybug (Hemiptera: Coccoidea: Pseudococcidae) pest in Florida and the Caribbean: The papaya mealybug, Paracoccus marginatus Williams and Granara de Willink. Insecta Mundi. 1999, 13, 179–181. [Google Scholar]
- Finch, E.A.; Beale, T.; Chellappan, M.; Goergen, G.; Gadratagi, B.G.; Khan, M.A.; Rehman, M.A.; Rwomushana, I.; Sarma, A.K.; Wyckhuys, K.A.; et al. The potential global distribution of the papaya mealybug, Paracoccus marginatus, a polyphagous pest. Pest Manag. Sci. 2021, 77, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, J.U.; George, M.; Ajesh, G.; Jithine, J.R.; Lekshmi, N.R.; Deepasree, M.I. A review on Paracoccus marginatus Williams, papaya mealybug (Hemiptera: Pseudococcidae). J. Entomol. Zool. Stud. 2016, 4, 528–533. [Google Scholar]
- Huang, F.; Zhang, Z.J.; Li, W.D.; Lin, W.C.; Zhang, P.J.; Zhang, J.M.; Bei, Y.W.; He, Y.P.; Lu, Y.B. Host plant probing analysis reveals quick settlement of the solenopsis mealybug during host shift. J. Econ. Entomol. 2014, 107, 1419–1425. [Google Scholar] [CrossRef]
- Lu, H.; Yang, P.C.; Xu, Y.Y.; Luo, L.; Zhu, J.J.; Cui, N.; Kang, L.; Cui, F. Performances of survival, feeding behavior, and gene expression in aphids reveal their different fitness to host alteration. Sci. Rep. 2016, 6, 19344. [Google Scholar] [CrossRef]
- Ning, S.Y.; Zhang, W.C.; Sun, Y.; Feng, J.N. Development of insect life tables: Comparison of two demographic methods of Delia antiqua (Diptera: Anthomyiidae) on different hosts. Sci. Rep. 2017, 7, 4821. [Google Scholar] [CrossRef]
- Milanović, S.; Janković-Tomanić, M.; Kostić, I.; Kostić, M.; Morina, F.; Živanović, B.; Lazarević, J. Behavioural and physiological plasticity of gypsy moth larvae to host plant switching. Entomol. Exp. Et Appl. 2016, 158, 152–162. [Google Scholar] [CrossRef]
- Mody, K.; Unsicker, S.B.; Linsenmair, K.E. Fitness related diet-mixing by intraspecific host-plant-switching of specialist insect herbivores. Ecology 2007, 88, 1012–1020. [Google Scholar] [CrossRef]
- Fan, Y.J.; Fen, L.; Abd, A.A.H.M.; Yi, X.Q.; Zhang, M.; Nicolas, D.; Gao, X.W. The damage risk evaluation of Aphis gossypii on wheat by host shift and fitness comparison in wheat and cotton. J. Integr. Agric. 2018, 17, 631–639. [Google Scholar] [CrossRef]
- Amarasekare, K.G.; Mannion, C.M.; Osborne, L.S.; Epsky, N.D. Life history of Paracoccus marginatus (Hemiptera: Pseudococcidae) on four host plant species under laboratory conditions. Environ. Entomol. 2008, 37, 630–635. [Google Scholar] [CrossRef]
- Nisha, R.; Kennedy, J.S. Life cycle of papaya mealybug Paracoccus marginatus Williams and Granara de Willink on different host plants vis-à-vis divergent natural selection. J. Entomol. Zool. Stud. 2017, 5, 91–102. [Google Scholar]
- Huang, Y.B.; Chi, H. Assessing the application of the jackknife and bootstrap techniques to the estimation of the variability of the net reproductive rate and gross reproductive rate: A case study in Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). J. Agric. For. 2012, 61, 37–45. [Google Scholar]
- Mou, D.F.; Lee, C.C.; Smith, C.L.; Chi, H. Using viable eggs to accurately determine the demographic and predation potential of Harmonia dimidiata (Coleoptera: Coccinellidae). J. Appl. Entomol. 2015, 139, 579–591. [Google Scholar] [CrossRef]
- Chi, H.; Liu, S. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; National Chung Hsing University: Taichung, Taiwan, 2022. [Google Scholar]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman & Hall: New York, NY, USA, 1993. [Google Scholar]
- Chi, H.; You, M.S.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Özgökçe, M.S.; Güncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 102–123. [Google Scholar] [CrossRef]
- Wei, M.F.; Chi, H.; Guo, Y.F.; Li, X.W.; Zhao, L.L.; Ma, R.Y. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri (Rosales: Rosaceae) and P. communis pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef]
- Euler, L. De aptissima figura rotarum dentibus tribuenda. Novi Comment. Acad. Sci. Petropolitanae 1760, 17, 299–316. [Google Scholar]
- Lotka, A.J. Studies on the mode of growth of material aggregates. Am. J. Sci. 1907, 24, 199–216. [Google Scholar] [CrossRef]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Fisher, R.A. The Genetical Theory of Natural Selection; Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Timing of control based on the stage structure of pest populations: A simulation approach. J. Econ. Entomol. 1990, 83, 1143–1150. [Google Scholar] [CrossRef]
- Chi, H. Timing-MSChart: A Computer Program for the Population Projection Based on Age-Stage, Two-Sex Life Table; National Chung Hsing University: Taichung, Taiwan, 2022. [Google Scholar]
- Huang, H.W.; Chi, H.; Smith, C.L. Linking demography and consumption of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae) fed on Solanum photeinocarpum: With a new method to project the uncertainty of population growth and consumption. J. Econ. Entomol. 2018, 111, 1–9. [Google Scholar] [CrossRef]
- Fu, J.W.; Shi, M.Z.; Wang, T.; Li, J.Y.; Zheng, L.Z.; Wu, G. Demography and population projection of flea beetle, Agasicles hygrophila (Coleoptera: Chrysomelidae), fed on alligator weed under elevated CO2. J. Econ. Entomol. 2016, 109, 1116–1124. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.M.; Chi, H.; Wang, R.C.; Wang, Y.P.; Xu, Y.Y.; Li, X.D.; Yin, P.; Zheng, F.Q. Demography and uncertainty of population growth of Conogethes punctiferalis (Lepidoptera: Crambidae) reared on five host plants with discussion on some life history statistics. J. Econ. Entomol. 2018, 111, 2143–2152. [Google Scholar] [CrossRef]
- Mastoi, M.I.; Adam, N.A.; Muhamad, R.; Ghani, I.A.; Gilal, A.A.; Khan, J.; Bhatti, A.R.; Zia, A.; Sahito, J.G.M. Efficiency of Acerophagus papayae on different host stage combinations of papaya mealybug, Paracoccus marginatus. Pak. J. Agric. Sci. 2018, 55, 375–379. [Google Scholar]
- Shi, M.Z.; Li, J.Y.; Ding, B.; Fu, J.W.; Zheng, L.Z.; Chi, H. Indirect effect of elevated CO2 on population parameters and growth of Agasicles hygrophila (Coleoptera: Chrysomelidae), a biocontrol agent of alligatorweed (Amaranthaceae). J. Econ. Entomol. 2019, 112, 1120–1129. [Google Scholar] [CrossRef]
- Jha, R.K.; Chi, H.; Tang, L.C. Effects of survival rate and fecundity on the intrinsic rate of increase of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Formos. Entomol. 2012, 32, 223–235. [Google Scholar]
- Seni, A.; Sahoo, A.K. Biology of Paracoccus marginatus Williams and Granara de Willink (Hemiptera: Pseudococcidae) on papaya, parthenium and brinjal plants. Res. Crop. 2015, 16, 722–727. [Google Scholar] [CrossRef]
- He, L.M.; Wu, Q.L.; Gao, X.W.; Wu, K.M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Karimi-Pormehr, M.S.; Borzoui, E.; Naseri, B.; Dastjerdi, H.R.; Mansouri, S.M. Two-sex life table analysis and digestive physiology of Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) on different barley cultivars. J. Stored Prod. Res. 2018, 75, 64–71. [Google Scholar] [CrossRef]
- Lewontin, R.C. Selection for Colonizing Ability; Baker, H.G., Stebbins, G.L., Eds.; The Genetics of Colonizing Species; Academic Press: San Diego, CA, USA, 1965; pp. 77–94. [Google Scholar]
- Quezada, G.R.; Seehausen, M.L.; Bauce, É. Adaptation of an outbreaking insect defoliator to chronic nutritional stress. J. Evolution. Biol. 2015, 28, 347–355. [Google Scholar] [CrossRef]
- Meihls, L.N.; Higdon, M.L.; Siegfried, B.D.; Miller, N.J.; Sappington, T.W.; Ellersieck, M.R.; Spencerc, T.A.; Hibbard, B.E. Increased survival of western corn rootworm on transgenic corn within three generations of on-plant greenhouse selection. Proc. Natl. Acad. Sci. USA 2008, 105, 19177–19182. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Nemati-Kalkhoran, M.; Razmjou, J.; Borzoui, E.; Naseri, B. Comparison of life table parameters and digestive physiology of Rhyzopertha dominica (Coleoptera: Bostrichidae) fed on various barley cultivars. J. Insect Sci. 2018, 18, 31. [Google Scholar] [CrossRef]
- Jaleel, W.; Tao, X.B.; Wang, D.S.; Lu, L.H.; He, Y.R. Using two-sex life table traits to assess the fruit preference and fitness of Bactrocera dorsalis (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2936–2945. [Google Scholar] [CrossRef]
- Cipollini, D.; Peterson, D.L. The potential for host switching via ecological fitting in the emerald ash borer-host plant system. Oecologia 2018, 187, 507–519. [Google Scholar] [CrossRef]
- Tan, S.L. Sweet potato-Ipomoea batatas-a great health food. Agric. Sci. 2015, 3, 15–28. [Google Scholar]
- Singh, A.; Yemmireddy, V. Fate of Salmonella spp. in fresh-cut papaya (Carica papaya L.) at different storage temperature and relative humidity. LWT Food Sci. Technol. 2021, 148, 111810. [Google Scholar] [CrossRef]
- Tian, W.J.; He, G.D.; Qin, L.J.; Li, D.D.; Meng, L.L.; Huang, Y.; He, T.B. Genome-wide analysis of the NRAMP gene family in potato (Solanum tuberosum): Identification, expression analysis and response to five heavy metals stress. Ecotoxicol. Environ. Saf. 2021, 208, 111661. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage (d) | Generation | Carica papaya | Ipomoea batatas | Solanum tuberosum | Alternanthera philoxeroides | ||||
|---|---|---|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | n | Mean ± SE | n | Mean ± SE | ||
| Egg | F0 | 95 | 5.18 ± 0.04 aA | 100 | 5.21 ± 0.04 aC | 100 | 5.20 ± 0.08 aB | 100 | 5.23 ± 0.05 a |
| F1 | 92 | 5.16 ± 0.06 bA | 98 | 6.22 ± 0.05 aB | 88 | 6.26 ± 0.06 aA | - | - | |
| F2 | 100 | 5.24 ± 0.06 cA | 90 | 6.91 ± 0.04 aA | 84 | 6.13 ± 0.09 bA | - | - | |
| Female nymph | F0 | 47 | 12.36 ± 0.16 cA | 17 | 14.29 ± 0.41 bA | 45 | 14.04 ± 0.30 bA | 2 | 18.50 ± 0.50 a |
| F1 | 45 | 11.98 ± 0.18 bA | 30 | 14.00 ± 0.31 aA | 40 | 15.07 ± 0.47 aA | - | - | |
| F2 | 49 | 12.24 ± 0.16 bA | 33 | 13.70 ± 0.34 aA | 40 | 14.32 ± 0.35 aA | - | - | |
| Male nymph | F0 | 39 | 13.92 ± 0.21 dA | 65 | 17.18 ± 0.30 bA | 44 | 15.43 ± 0.20 cA | 9 | 20.22 ± 1.05 a |
| F1 | 40 | 13.50 ± 0.26 bA | 53 | 15.53 ± 0.24 aB | 44 | 15.20 ± 0.19 aAB | - | - | |
| F2 | 44 | 13.64 ± 0.21 cA | 51 | 15.55 ± 0.15 aB | 38 | 14.61 ± 0.24 bB | - | - | |
| Female adult | F0 | 47 | 20.09 ± 0.55 aA | 17 | 15.65 ± 1.23 bA | 45 | 19.87 ± 0.94 abA | 2 | 8.00 ± 1.00 c |
| F1 | 45 | 19.18 ± 0.56 aA | 30 | 18.53 ± 1.63 aA | 40 | 13.97 ± 1.27 bB | - | - | |
| F2 | 49 | 19.67 ± 0.56 aA | 33 | 17.12 ± 1.22 aA | 40 | 14.28 ± 1.44 bB | - | - | |
| Male adult | F0 | 39 | 3.28 ± 0.18 aA | 65 | 3.48 ± 0.14 aA | 44 | 3.41 ± 0.20 aA | 9 | 3.67 ± 0.62 a |
| F1 | 40 | 3.48 ± 0.17 aA | 53 | 3.21 ± 0.14 aAB | 44 | 3.11 ± 0.15 aAB | - | - | |
| F2 | 44 | 3.50 ± 0.16 aA | 51 | 3.00 ± 0.13 bB | 38 | 2.87 ± 0.13 bB | - | - | |
| Population Parameter | Generation | Carica papaya | Ipomoea batatas | Solanum tuberosum | Alternanthera philoxeroides |
|---|---|---|---|---|---|
| Preadult survival rate (sa) | F0 | 0.905 ± 0.030 aA | 0.820 ± 0.038 aB | 0.890 ± 0.031 aA | 0.110 ± 0.031 b |
| F1 | 0.924 ± 0.028 abA | 0.847 ± 0.036 bB | 0.955 ± 0.022 aA | - | |
| F2 | 0.930 ± 0.026 aA | 0.933 ± 0.026 aA | 0.929 ± 0.028 aA | - | |
| Proportion of female adults (Nf/N) | F0 | 0.495 ± 0.051 aA | 0.170 ± 0.038 bB | 0.450 ± 0.050 aA | 0.023 ± 0.012 c |
| F1 | 0.489 ± 0.052 aA | 0.306 ± 0.047 bA | 0.455 ± 0.053 aA | - | |
| F2 | 0.490 ± 0.050 aA | 0.367 ± 0.051 aA | 0.476 ± 0.055 aA | - | |
| Proportion of male adults (Nm/N) | F0 | 0.411 ± 0.050 bA | 0.650 ± 0.048 aA | 0.440 ± 0.049 bA | 0.090 ± 0.050 c |
| F1 | 0.435 ± 0.052 aA | 0.541± 0.050 aA | 0.500 ± 0.053 aA | - | |
| F2 | 0.440 ± 0.049 aA | 0.567 ± 0.052 aA | 0.452 ± 0.054 aA | - | |
| Fecundity (F) (hatch eggs/female) | F0 | 202.70 ± 17.68 aA | 98.93 ± 23.04 bB | 127.46 ± 18.79 bA | 0 |
| F1 | 202.46 ± 24.08 aA | 229.50 ± 41.84 aA | 127.38 ± 25.40 bA | - | |
| F2 | 215.27 ± 20.32 aA | 203.76 ± 35.72 abA | 121.51 ± 25.35 bA | - |
| Population Parameter | Generation | Carica papaya | Ipomoea batatas | Solanum tuberosum |
|---|---|---|---|---|
| Net reproductive rate (R0) (offspring) | F0 | 100.26 ± 13.56 aA | 16.82 ± 5.31 cB | 57.36 ± 10.53 bA |
| F1 | 98.97 ±15.76 aA | 70.32 ±16.63 abA | 57.98 ± 13.31 bA | |
| F2 | 105.51 ± 4.98 aA | 74.64 ±5.09 abA | 57.88 ±13.74 bA | |
| Intrinsic rate of increase (r) (d−1) | F0 | 0.1778 ± 0.0053 aA | 0.0926 ± 0.0109 cB | 0.1376 ± 0.0069 bA |
| F1 | 0.1747 ± 0.0064 aA | 0.1357 ± 0.0080 bA | 0.1389 ± 0.0084 bA | |
| F2 | 0.1824 ± 0.0064 aA | 0.1435 ± 0.0075 bA | 0.1361 ± 0.0085 bA | |
| Finite rate of increase (λ) (d−1) | F0 | 1.1945 ± 0.0064 aA | 1.0970 ± 0.0119 cB | 1.1475 ± 0.0079 bA |
| F1 | 1.1909 ± 0.0076 aA | 1.1454 ± 0.0091 bA | 1.1490 ± 0.0097 bA | |
| F2 | 1.2000 ± 0.0076 aA | 1.1543 ± 0.0086 bA | 1.1459 ± 0.0097 bA | |
| Mean generation time (T) (d) | F0 | 25.92 ± 0.30 bA | 30.48 ± 1.08 aAB | 29.43 ± 0.61 aA |
| F1 | 26.30 ± 0.46 cA | 31.34 ± 0.42 aA | 29.23 ± 0.58 bA | |
| F2 | 25.56 ± 0.40 bA | 30.05 ± 0.39 aB | 29.81 ± 0.61 aA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuai, H.-Y.; Shi, M.-Z.; Li, J.-Y.; Zheng, L.-Z.; Fu, J.-W. Fitness of the Papaya Mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae), after Transferring from Solanum tuberosum to Carica papaya, Ipomoea batatas, and Alternanthera philoxeroides. Insects 2022, 13, 804. https://doi.org/10.3390/insects13090804
Chuai H-Y, Shi M-Z, Li J-Y, Zheng L-Z, Fu J-W. Fitness of the Papaya Mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae), after Transferring from Solanum tuberosum to Carica papaya, Ipomoea batatas, and Alternanthera philoxeroides. Insects. 2022; 13(9):804. https://doi.org/10.3390/insects13090804
Chicago/Turabian StyleChuai, Hui-Yu, Meng-Zhu Shi, Jian-Yu Li, Li-Zhen Zheng, and Jian-Wei Fu. 2022. "Fitness of the Papaya Mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae), after Transferring from Solanum tuberosum to Carica papaya, Ipomoea batatas, and Alternanthera philoxeroides" Insects 13, no. 9: 804. https://doi.org/10.3390/insects13090804
APA StyleChuai, H.-Y., Shi, M.-Z., Li, J.-Y., Zheng, L.-Z., & Fu, J.-W. (2022). Fitness of the Papaya Mealybug, Paracoccus marginatus (Hemiptera: Pseudococcidae), after Transferring from Solanum tuberosum to Carica papaya, Ipomoea batatas, and Alternanthera philoxeroides. Insects, 13(9), 804. https://doi.org/10.3390/insects13090804