
Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae)

 ,
,  , ,
, ,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
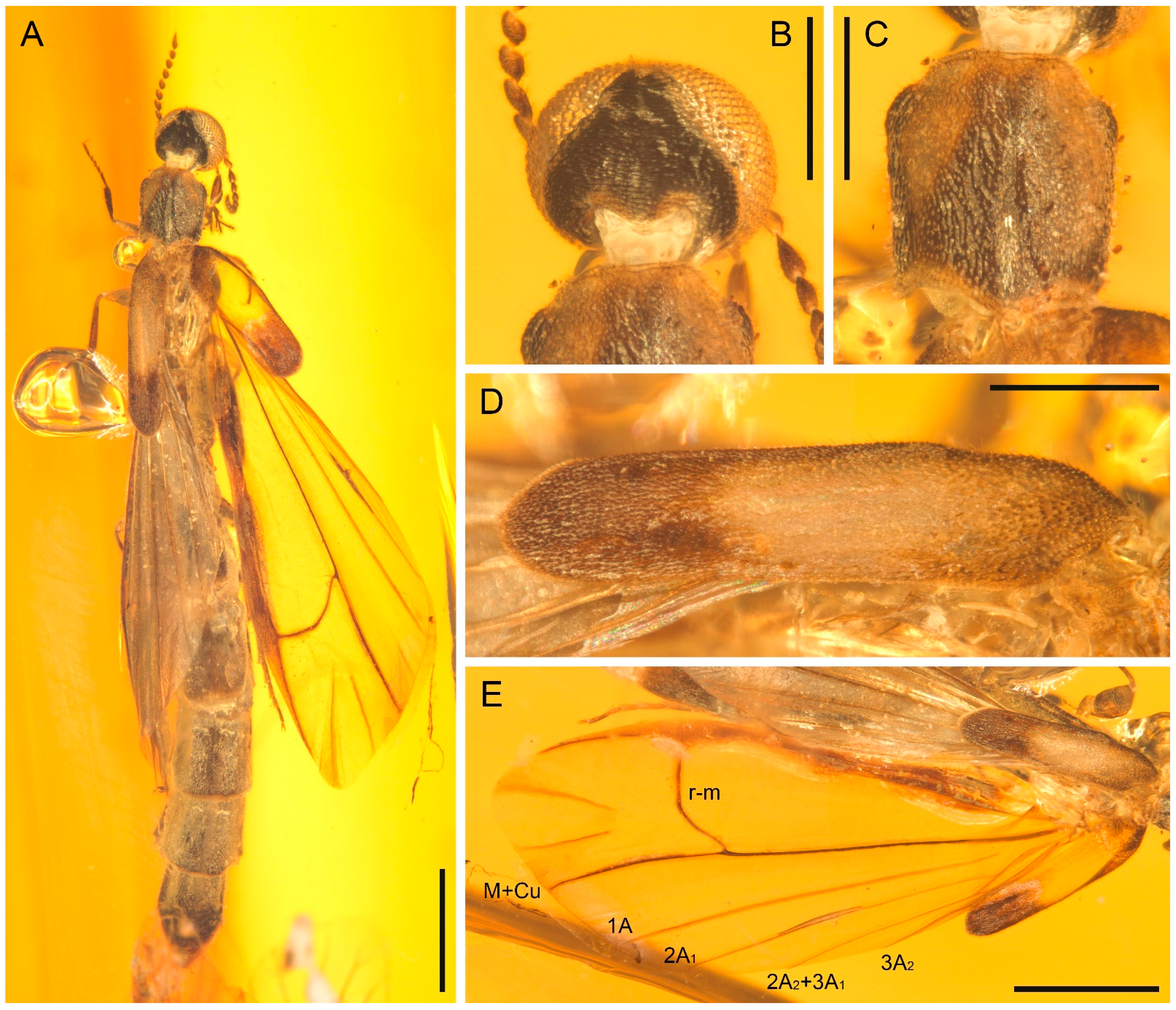
3. Systematic Palaeontology
- Order Coleoptera Linnaeus, 1758
- Suborder Polyphaga Emery, 1886
- Superfamily Lymexyloidea Fleming, 1821
- Family Lymexylidae Fleming, 1821
- Subfamily Atractocerinae Laporte, 1840
- Genus Cretoquadratus Chen, 2019
- Type species. Cretoquadratus engeli Chen, 2019.
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paulus, H. Urtea graeca nov. gen. et nov. spec., der erste Vertreter der tropischen Atractocerinae in Europa sowie eine Beschreibung von Hymaloxylon aspoecki nov. spec. aus Yunnan (China) (Coleoptera, Cucujiformia, Lymexylidae, Atractocerinae nov. status). Denisia 2004, 13, 277–290. [Google Scholar]
- Wolf-Schwenninger, K. The oldest fossil record of Lymexylidae (Insecta: Coleoptera) from the Lower Cretaceous Crato formation of Brazil. Insect Syst. Evol. 2011, 42, 205–212. [Google Scholar] [CrossRef]
- Kirejtshuk, A. A new genus and species of timber beetle (Coleoptera: Lymexylidae) from the Baltic amber. Paleontol. J. 2008, 42, 63–65. [Google Scholar] [CrossRef]
- Chen, X. A new genus and species of Lymexylidae (Insecta: Coleoptera) from mid-Cretaceous amber of northern Myanmar. Paleontol. J. 2019, 53, 1203–1205. [Google Scholar] [CrossRef]
- Yamamoto, S. Fossil evidence of elytra reduction in ship-timber beetles. Sci. Rep. 2019, 9, 4938. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Nazarenko, V.Y.; Vasilenko, D.V.; Perkovsky, E.E. First fossil species of ship-timber beetles (Coleoptera, Lymexylidae) from Eocene Rovno amber (Ukraine). Foss. Rec. 2022, 25, 65–74. [Google Scholar] [CrossRef]
- Nazarenko, V.; Perkovsky, E.; Vasilenko, D. New genus and species of Lymexylidae (Coleoptera) from Baltic amber. Paleontol. J. 2020, 54, 1395–1399. [Google Scholar] [CrossRef]
- Chen, X.-Y.; Zhang, H.-C. A new fossil record of Lymexylidae (Insecta: Coleoptera) from mid-Cretaceous amber of northern Myanmar. Zootaxa 2020, 4878, 195–200. [Google Scholar] [CrossRef]
- Perkovsky, E. Anancites nom. nov., a replacement name for Adamas Chen et Wang, 2020 (Coleoptera, Lymexylidae). Paleontol. J. 2022, 56, 327. [Google Scholar] [CrossRef]
- Crowson, R. The phylogeny of Coleoptera. Annu. Rev. Entomol. 1960, 5, 111–134. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Newton, A.F. Evolution and classification of beetles. Annu. Rev. Ecol. Syst. 1982, 13, 261–290. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J.A.; Ślipiński, A.; Moulton, M.; Shockley, F.W.; Giorgi, A.; Lord, N.P.; Mckenna, D.D.; Tomaszewska, W.; Forrester, J.; Miller, K.B. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst. Entomol. 2015, 40, 745–778. [Google Scholar] [CrossRef]
- Mckenna, D.D.; Wild, A.L.; Kanda, K.; Bellamy, C.L.; Beutel, R.G.; Caterino, M.S.; Farnum, C.W.; Hawks, D.C.; Ivie, M.A.; Jameson, M.L. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 2015, 40, 835–880. [Google Scholar] [CrossRef]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; John, O.S.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef]
- Bocak, L.; Barton, C.; Crampton-Platt, A.; Chesters, D.; Ahrens, D.; Vogler, A.P. Building the Coleoptera tree-of-life for >8000 species: Composition of public DNA data and fit with Linnaean classification. Syst. Entomol. 2014, 39, 97–110. [Google Scholar] [CrossRef]
- Gunter, N.L.; Levkaničová, Z.; Weir, T.H.; Ślipiński, A.; Cameron, S.L.; Bocak, L. Towards a phylogeny of the Tenebrionoidea (Coleoptera). Mol. Phylogenet. Evol. 2014, 79, 305–312. [Google Scholar] [CrossRef]
- Zhang, S.-Q.; Che, L.-H.; Li, Y.; Liang, D.; Pang, H.; Ślipiński, A.; Zhang, P. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 2018, 9, 205. [Google Scholar] [CrossRef]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef]
- Cai, C.; Tihelka, E.; Giacomelli, M.; Lawrence, J.F.; Slipinski, A.; Kundrata, R.; Yamamoto, S.; Thayer, M.K.; Newton, A.F.; Leschen, R.A.; et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. R. Soc. Open Sci. 2022, 9, 211771. [Google Scholar] [CrossRef]
- Wheeler, Q.D. Revision of the genera of Lymexylidae (Coleoptera, Cucujiformia). Bull. Am. Mus. Nat. Hist. 1986, 183, 113–210. [Google Scholar]
- Kirejtshuk, A.G.; Azar, D. Current knowledge of Coleoptera (Insecta) from the Lower Cretaceous Lebanese amber and taxonomical notes for some Mesozoic groups. Terr. Arthropod Rev. 2013, 6, 103–134. [Google Scholar] [CrossRef]
- Solórzano Kraemer, M.M.; Delclòs, X.; Clapham, M.E.; Arillo, A.; Peris, D.; Jäger, P.; Stebner, F.; Peñalver, E. Arthropods in modern resins reveal if amber accurately recorded forest arthropod communities. Proc. Natl. Acad. Sci. USA 2018, 115, 6739–6744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G., Jr.; Lambert, J.B.; Wu, Y. Araucarian source of fossiliferous Burmese amber: Spectroscopic and anatomical evidence. J. Bot. Res. Inst. Tex. 2007, 1, 449–455. [Google Scholar]
- Wolfe, A.P.; Tappert, R.; Muehlenbachs, K.; Boudreau, M.; McKellar, R.C.; Basinger, J.F.; Garrett, A. A new proposal concerning the botanical origin of Baltic amber. Proc. R. Soc. B 2009, 276, 3403–3412. [Google Scholar] [CrossRef]
- Wolfe, A.P.; McKellar, R.C.; Tappert, R.; Sodhi, R.N.; Muehlenbachs, K. Bitterfeld amber is not Baltic amber: Three geochemical tests and further constraints on the botanical affinities of succinite. Rev. Palaeobot. Palynol. 2016, 225, 21–32. [Google Scholar] [CrossRef]
- Perkovsky, E.E.; Zosimovich, V.Y.; Vlaskin, A.P. Rovno amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Fu, Y.-Z.; Li, Y.-D.; Su, Y.-T.; Cai, C.-Y.; Huang, D.-Y. Application of confocal laser scanning microscopy to the study of amber bioinclusions. Palaeoentomology 2021, 4, 266–278. [Google Scholar] [CrossRef]
- Kurosawa, Y. Revisional notes on the family Lymexylonidae (Coleoptera) in eastern and southeastern Asia. Bull. Natl. Sci. Mus. Ser. A 1985, 11, 109–119. [Google Scholar]
- Casari, S.A.; Albertoni, F.F. First instar larva of Atractocerus brasiliensis (Lepeletier & Audinet-Serville, 1825) (Lymexylidae, Atractocerinae). Pap. Avulsos Zool. 2013, 53, 359–372. [Google Scholar]
- Lawrence, J.F. The Australian Lymexylidae (Coleoptera: Tenerionoidea) with one new genus and two genera new to Australia. Zootaxa 2020, 4895, 211–238. [Google Scholar] [CrossRef]
- Francke-Grosmann, H. Ectosymbiosis in wood-inhabiting insects. In Symbiosis. Volume II. Associations of Invertebrates, Birds, Ruminants, and Other Biota; Henry, S., Ed.; Academic Press: New York, NY, USA, 1967; pp. 141–205. [Google Scholar]
- Toki, W. A single case study of mycetangia-associated fungi and their abilities to assimilate wood-associated carbon sources in the ship timber beetle Elateroides flabellicornis (Coleoptera: Lymexylidae) in Japan. Symbiosis 2021, 83, 173–181. [Google Scholar] [CrossRef]
- Casari, S.A.; Teixeira, É.P. Larva of Atractocerus brasiliensis (Lepeletier & Audinet-Serville, 1825) (Lymexylidae, Atractocerinae). Pap. Avulsos Zool. 2011, 51, 197–205. [Google Scholar]
- Peris, D.; Delclòs, X.; Jordal, B. Origin and evolution of fungus farming in wood-boring Coleoptera—A palaeontological perspective. Biol. Rev. 2021, 96, 2476–2488. [Google Scholar] [PubMed]
- Kirkendall, L.R.; Biedermann, P.H.; Jordal, B.H. Evolution and diversity of bark and ambrosia beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: London, UK, 2015; pp. 85–156. [Google Scholar]
- Peris, D. Coleoptera in amber from Cretaceous resiniferous forests. Cretac. Res. 2020, 113, 104484. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Azar, D.; Beaver, R.A.; Mandelshtam, M.Y.; Nel, A. The most ancient bark beetle known: A new tribe, genus and species from Lebanese amber (Coleoptera, Curculionidae, Scolytinae). Syst. Entomol. 2009, 34, 101–112. [Google Scholar] [CrossRef]
- Pistone, D.; Gohli, J.; Jordal, B.H. Molecular phylogeny of bark and ambrosia beetles (Curculionidae: Scolytinae) based on 18 molecular markers. Syst. Entomol. 2018, 43, 387–406. [Google Scholar] [CrossRef]
- Peris, D.; Rust, J. Cretaceous beetles (Insecta: Coleoptera) in amber: The palaeoecology of this most diverse group of insects. Zool. J. Linn. Soc. 2020, 189, 1085–1104. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.-D.; Peris, D.; Yamamoto, S.; Hsiao, Y.; Newton, A.F.; Cai, C.-Y. Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae). Insects 2022, 13, 768. https://doi.org/10.3390/insects13090768
Li Y-D, Peris D, Yamamoto S, Hsiao Y, Newton AF, Cai C-Y. Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae). Insects. 2022; 13(9):768. https://doi.org/10.3390/insects13090768
Chicago/Turabian StyleLi, Yan-Da, David Peris, Shûhei Yamamoto, Yun Hsiao, Alfred F. Newton, and Chen-Yang Cai. 2022. "Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae)" Insects 13, no. 9: 768. https://doi.org/10.3390/insects13090768
APA StyleLi, Y.-D., Peris, D., Yamamoto, S., Hsiao, Y., Newton, A. F., & Cai, C.-Y. (2022). Revisiting the Raractocetus Fossils from Mesozoic and Cenozoic Amber Deposits (Coleoptera: Lymexylidae). Insects, 13(9), 768. https://doi.org/10.3390/insects13090768