

Where Does Honey Bee (Apis mellifera L.) Pollen Come from? A Study of Pollen Collected from Colonies at Ornamental Plant Nurseries

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
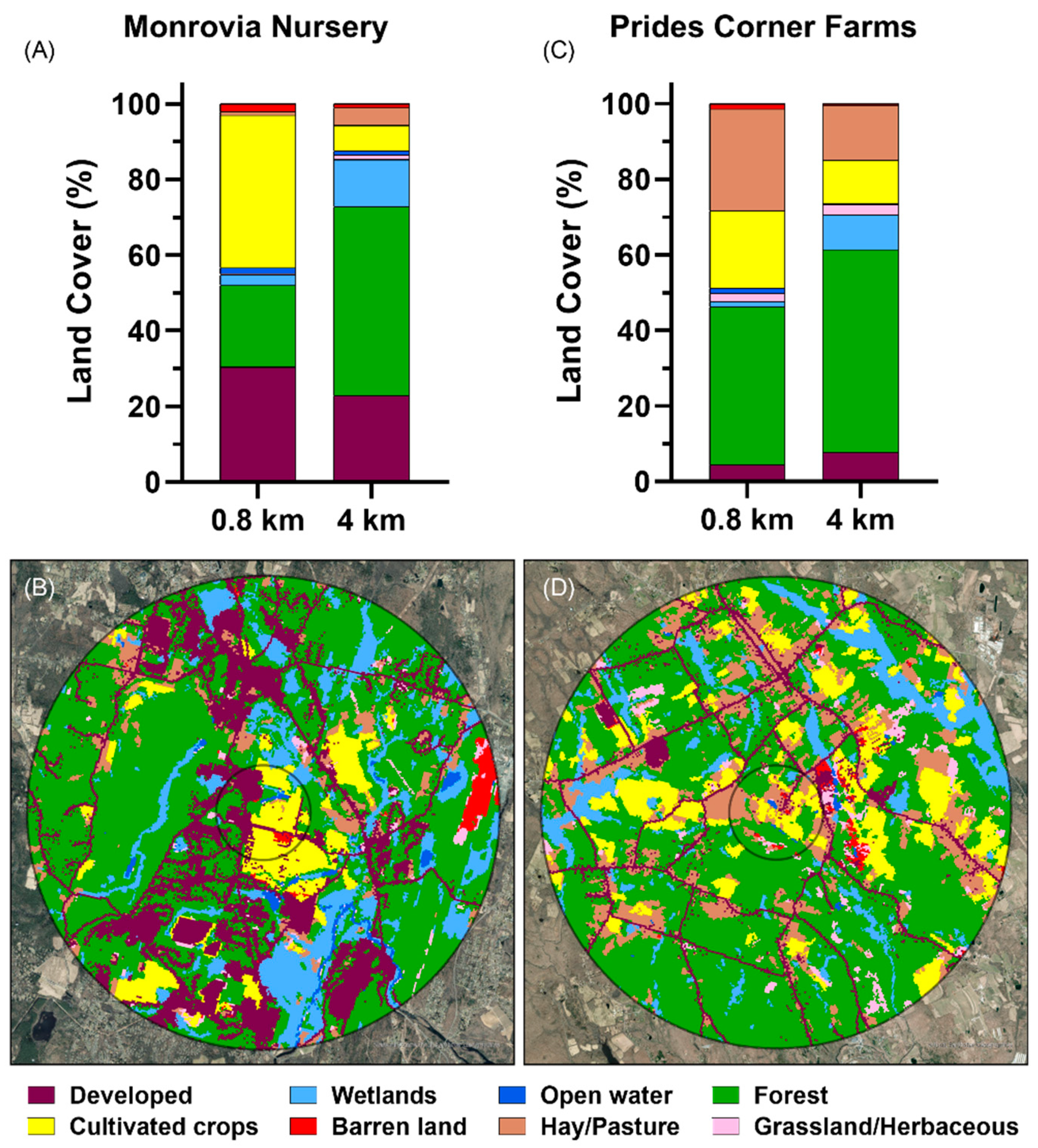
3.1. Land Cover Composition at the Nursery Sites
3.2. Percent of Total Pollen Volume from Genera Grown at the Nursery
3.3. Genera in Pollen Samples at Each Nursery
3.4. Major Plant Sources of Pollen through the Season
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monrovia Nursery | Prides Corner Farms | ||||
|---|---|---|---|---|---|
| Genus or Family | cf | Grain volume (µm3) | Genus or family | cf | Grain volume (µm3) |
| Acer | 4920 | Achillea | cf | 7238 | |
| Amaranthus | 6371 | Actaea | 22,449 | ||
| Ambrosia/Xanthium | 3054 | Aesculus | 3351 | ||
| Amorpha | 3393 | Ajuga | 4581 | ||
| Andropogon | 22,093 | Alisma | cf | 22,449 | |
| Anthyllis | cf | 8143 | Allium | 6447 | |
| Antirrhinum | 4618 | Ambrosia/Xanthium | 3236 | ||
| Aquilegia | 3054 | Amorpha | 3393 | ||
| Asparagus | 2360 | Anthyllis | 10,619 | ||
| Astragalus | 5236 | Aquilegia | 4817 | ||
| Baptisia | 2121 | Aralia | 11,494 | ||
| Bellis | cf | 8181 | Arctium | cf | 3054 |
| Berberis | 22,449 | Asparagus | 2360 | ||
| Calystegia | 203,689 | Astragalus | cf | 1595 | |
| Capsella | cf | 1767 | Baptisia | 1227 | |
| Carum | cf | 1697 | Begonia | 295 | |
| Castanea | 636 | Bellis | cf | 8181 | |
| Centaurea | 11,494 | Buddleja | 733 | ||
| Cephalanthus | 3393 | Buxus | cf | 14,137 | |
| Chelidonium | 6371 | Callitriche | 697 | ||
| Chenopodium | 9203 | Campanula | 11,494 | ||
| Cirsium | 22,449 | Capsella | cf | 1767 | |
| Clematis | 8181 | Caragana | 1593 | ||
| Coreopsis | 11,494 | Cardamine | cf | 3054 | |
| Cucumis | 87,114 | Carya | 23,732 | ||
| Cucurbita | 530,241 | Castanea | 636 | ||
| Daucus | cf | 1882 | Celastrus | 6635 | |
| Dianthus | cf | 47,713 | Celosia | 28,731 | |
| Epilobium | cf | 523,600 | Centaurea | 11,494 | |
| Erigeron | cf | 6371 | Cephalanthus | 3393 | |
| Eupatorium/Eutrochium | 6648 | Chelidonium | 11,494 | ||
| Euphorbia | 8310 | Chenopodium | 6371 | ||
| Fagopyrum | 7238 | Cichorium | cf | 47,713 | |
| Fallopia | 3393 | Cirsium | 22,449 | ||
| Fraxinus | 4189 | Clematis | 5540 | ||
| Fuchsia | cf | 102,161 | Colutea | cf | 3485 |
| Funaria | moss | 2124 | Coreopsis | 11,494 | |
| Gaillardia | cf | 41,630 | Crocosmia | cf | 17,999 |
| Geranium | 77,952 | Cytisus | 3732 | ||
| Hedera | cf | 17,157 | Dahlia | cf | 15,599 |
| Heiracium | 7238 | Daucus | cf | 1882 | |
| Helianthus | cf | 18,817 | Digitalis | 4189 | |
| Hemerocallis | 44,899 | Echinops | 25,656 | ||
| Heuchera | 905 | Epilobium | 747,596 | ||
| Hydrangea | 1327 | Erigeron | cf | 6086 | |
| Hypericum | 509 | Eupatorium/Eutrochium | cf | 4398 | |
| Ilex | 10,688 | Fagopyrum | 32,071 | ||
| Impatiens | 18,817 | Fagus | 7202 | ||
| Iris | 59,362 | Funaria | moss | 1947 | |
| Lepidium | cf | 8181 | Gaillardia | 41,630 | |
| Liatris | cf | 11,494 | Galium | 2572 | |
| Linaria | 2547 | Gaura | cf | 696,912 | |
| Lotus | 1327 | Heiracium | cf | 7238 | |
| Lycium | 14,137 | Helianthus | 18,817 | ||
| Lysimachia | 5445 | Hemerocallis | 73,999 | ||
| Lythrum | 11,579 | Hippuris | 8181 | ||
| Medicago | 8585 | Humulus | 6371 | ||
| Melilotus | cf | 4920 | Hydrangea | 1327 | |
| Mentha | 25,656 | Hypericum | 509 | ||
| Nuphar | 17,974 | Ilex | 8181 | ||
| Nymphaea | 17,974 | Impatiens | 6283 | ||
| Paeonia | 10,263 | Iris | 35,278 | ||
| Parthenocissus | 13,932 | Lagerstroemia | 15,080 | ||
| Philadelphus | 1593 | Lepidium | 324 | ||
| Phlox | 47,713 | Lespedeza | 2681 | ||
| Phryma | 4817 | Liatris | cf | 11,494 | |
| Phytolacca | 10,263 | Lamium | cf | 9140 | |
| Pinus | 78,703 | Liquidambar | 28,731 | ||
| Plantago | 7588 | Lonicera | cf | 38,725 | |
| Polygonum | 11,494 | Lotus | 1327 | ||
| Pontederia | 6049 | Lupinus | 3563 | ||
| Potentilla/Dasiphora | 1593 | Lysimachia | 2356 | ||
| Primula | 11,451 | Lythrum | 8890 | ||
| Quercus | 6648 | Malus | cf | 6925 | |
| Raphanus | cf | 3902 | Medicago | 5052 | |
| Rhamnus | 3223 | Melilotus | 4920 | ||
| Rhododendron | 32,511 | Mikania | cf | 14,137 | |
| Rhus | 13,854 | Morus | 1767 | ||
| Robinia | 6097 | Myriophyllum | 41,630 | ||
| Rosa | 5231 | Nyssa | 13,547 | ||
| Rubus | 2686 | Onobrychis | 5089 | ||
| Rumex | 8084 | Pedicularis | 1327 | ||
| Sagitaria | 6371 | Persicaria | 33,510 | ||
| Salvia | 16,605 | Phlox | 47,713 | ||
| Saxifraga | 4189 | Plantago | 7156 | ||
| Scutellaria | 8181 | Polygonatum | 56,968 | ||
| Solanum | 2015 | Polygonum | 34,024 | ||
| Solidago | cf | 3393 | Portulaca | 33,510 | |
| Spergula | 7238 | Potentilla | 1593 | ||
| Spiraea | 530 | Potentilla/Dasiphora | 1593 | ||
| Stellaria | cf | 7238 | Primula | 637 | |
| Swida | 17,652 | Quercus | 6648 | ||
| Symphyotrichum | cf | 14,137 | Ranunculus | 8181 | |
| Syringa | 9193 | Rhamnus | 5195 | ||
| Tanacetum | cf | 15,551 | Rhus | 14,380 | |
| Taraxacum | cf | 18,697 | Robinia | 6097 | |
| Tilia | 18,817 | Rosa | 3979 | ||
| Toxicodendron | 5753 | Rubus | 6336 | ||
| Tragopogon | 65,450 | Rudbeckia | cf | 14,137 | |
| Trifolium summed | 4337 | Rumex | 8818 | ||
| Urtica | 4849 | Salvia | 11,494 | ||
| Verbascum | 4817 | Sambucus | 3223 | ||
| Viburnum | 8181 | Sedum | 3054 | ||
| Vitis | 5175 | Solanum | 2145 | ||
| Zea | 248,475 | Solidago | cf | 3393 | |
| Zinnia | cf | 47,713 | Sparganium | 11,494 | |
| Crassulaceae | 2356 | Spiraea | 530 | ||
| Brassicaceae ≤ 20 µm | 3054 | Stellaria/Cerastium | 18,817 | ||
| Swida | 21,069 | ||||
| Syringa | 9193 | ||||
| Symphoricarpus | 33,510 | ||||
| Symphyotrichum | 14,137 | ||||
| Taraxacum | cf | 11,494 | |||
| Taxus | 14,137 | ||||
| Thalictrum | 4189 | ||||
| Toxicodendron | 3817 | ||||
| Trifolium summed | 5429 | ||||
| Urtica | 1767 | ||||
| Verbascum | 4817 | ||||
| Veronica | 8033 | ||||
| Viburnum | 12,464 | ||||
| Viola | 12,315 | ||||
| Vitis | 4817 | ||||
| Weigela | cf | 47,713 | |||
| Wisteria | 5236 | ||||
| Zea | 248,475 | ||||
| Apiaceae—no genus | 1697 | ||||
| Boraginaceae—no genus | 5575 | ||||
| Brassicaceae—no genus 18–20 um | 3054 | ||||
| Brassicaceae—no genus 20 um | 4189 | ||||
| Crassulaceae—no genus | 2356 | ||||
| Poaceae—large pore no genus | 21,167 | ||||
| Poaceae sp. 2 no genus | 19,957 | ||||
| Poaceae—no genus | 4189 | ||||
| Caprifoliaceae—no genus | 4189 | ||||
| Solanaceae | 8181 | ||||
| Annual Plant Sales as Reported by Nurseries at the Beginning of 2015 | Percentages Based on Reported Annual Sales | Additional Cultivated Genera from Visual Surveys, Pesticide Records, or Inventories | ||||
|---|---|---|---|---|---|---|
| Genus | No. Plants Prides Corner Farms | No. Plants Monrovia Nursery | % Prides Corner Farms | % Monrovia Nursery | Prides Corner Farms | Monrovia Nursery |
| Abelia | X | |||||
| ACER | 33,511 | 3080 | 1.4% | 0.2% | ||
| Achillea | X | X | ||||
| Agastache | X | X | ||||
| Ajuga | X | X | ||||
| Allium | X | |||||
| Anemone | X | X | ||||
| Aquilegia | X | X | ||||
| Armeria | X | |||||
| Asclepias | X | X | ||||
| Astilbe | X | X | ||||
| BERBERIS | 17,001 | 33,693 | 0.7% | 2.2% | ||
| BUDDLEIA | 33,688 | 14,634 | 1.4% | 0.9% | ||
| BUXUS | 129,842 | 49,348 | 5.3% | 3.2% | ||
| Calluna | X | |||||
| Campanula | X | X | ||||
| Caragana | X | |||||
| Caryopteris | X | X | ||||
| Catalpa | X | |||||
| Centaurea | X | |||||
| Cephalanthus | X | X | ||||
| Cercis | X | X | ||||
| Chaenomeles | X | X | ||||
| CLEMATIS | 22,327 | 17,975 | 0.9% | 1.2% | ||
| CLETHRA | 17,583 | 9380 | 0.7% | 0.6% | ||
| COREOPSIS | 18,477 | 13,805 | 0.8% | 0.9% | ||
| CORNUS (or Swida) | 45,303 | 7128 | 1.9% | 0.5% | ||
| Cotinus | X | X | ||||
| CYTISUS | 17,892 | 4827 | 0.7% | 0.3% | ||
| Delosperma | X | X | ||||
| Delphinium | X | |||||
| DEUTZIA | 24,336 | 5103 | 1.0% | 0.3% | ||
| DIANTHUS | 14,384 | 2520 | 0.6% | 0.2% | ||
| Dicentra | X | |||||
| Diervilla | X | X | ||||
| ECHINACEA | 52,876 | 25,706 | 2.2% | 1.6% | ||
| Erica | X | |||||
| EUONYMUS | 57,775 | 15,331 | 2.4% | 1.0% | ||
| Eupatorium | X | X | ||||
| FORSYTHIA | 17,852 | 8016 | 0.7% | 0.5% | ||
| Fothergilla | X | X | ||||
| Fragaria | X | X | ||||
| Gaillardia | X | X | ||||
| Gaura | X | X | ||||
| Geranium | X | X | ||||
| Geum | X | X | ||||
| Helenium | X | X | ||||
| Heliopsis | X | X | ||||
| HELLEBORUS | 23,545 | 10,571 | 1.0% | 0.7% | ||
| HEMEROCALLIS | 62,301 | 20,352 | 2.6% | 1.3% | ||
| HEUCHERA | 26,989 | 22,329 | 1.1% | 1.4% | ||
| HIBISCUS | 38,486 | 7057 | 1.6% | 0.5% | ||
| HOSTA | 54,946 | 68,531 | 2.3% | 4.4% | ||
| HYDRANGEA | 247,037 | 185,588 | 10.2% | 11.9% | ||
| Hypericum | X | X | ||||
| Iberis | X | X | ||||
| ILEX | 115,836 | 45,688 | 4.8% | 2.9% | ||
| IRIS | 17,962 | 6820 | 0.7% | 0.4% | ||
| ITEA | 9571 | 3751 | 0.4% | 0.2% | ||
| Lagerstroemia | X | |||||
| KALMIA | 44,291 | 20,804 | 1.8% | 1.3% | ||
| Kniphofia | X | X | ||||
| Lamium | X | |||||
| LAVANDULA | 33,734 | 5532 | 1.4% | 0.4% | ||
| LEUCANTHEMUM | 15,193 | 7628 | 0.6% | 0.5% | ||
| LEUCOTHOE | 24,502 | 1898 | 1.0% | 0.1% | ||
| Ligularia | X | X | ||||
| Lobelia | X | X | ||||
| Lonicera | X | X | ||||
| LUPINUS | 18,224 | 0.7% | ||||
| Lycium | X | X | ||||
| MALUS | 7790 | 6311 | 0.3% | 0.4% | ||
| MONARDA | 14,250 | 2062 | 0.6% | 0.1% | ||
| NEPETA | 18,642 | 0.8% | X | |||
| Oenothera | X | |||||
| PAEONIA | 8680 | 25,522 | 0.4% | 1.6% | ||
| Penstemon | X | x | ||||
| Perovskia | X | X | ||||
| Philadelphus | X | |||||
| PHLOX | 94,604 | 14,683 | 3.9% | 0.9% | ||
| PHYSOCARPUS | 27,417 | 8788 | 1.1% | 0.6% | ||
| PIERIS | 87,271 | 44,340 | 3.6% | 2.8% | ||
| Platycodon | X | X | ||||
| POTENTILLA (or Dasiphora) | 22,249 | 6694 | 0.9% | 0.4% | ||
| PRUNUS | 19,732 | 22,455 | 0.8% | 1.4% | ||
| Pyrus | X | |||||
| RHODODENDRON | 331,757 | 445,573 | 13.6% | 28.6% | ||
| Rhus | X | X | ||||
| ROSA | 104,066 | 162,585 | 4.3% | 10.4% | ||
| ROSMARINUS | 16,460 | 0.7% | ||||
| RUBUS | 17,575 | 9975 | 0.7% | 0.6% | ||
| RUDBECKIA | 20,905 | 3597 | 0.9% | 0.2% | ||
| SALIX | 14,478 | 0.6% | X | |||
| SALVIA | 26,184 | 15,425 | 1.1% | 1.0% | ||
| Sambucus | X | X | ||||
| SEDUM | 16,552 | 24,497 | 0.7% | 1.6% | ||
| Solanum (Eggplant) | X | |||||
| Solidago | X | X | ||||
| Sorbaria | X | X | ||||
| SPIRAEA | 80,128 | 42,086 | 3.3% | 2.7% | ||
| Stachys | X | |||||
| Stokesia | X | X | ||||
| Symphyotrichum (Aster) | X | X | ||||
| Symphoricarpos | X | |||||
| SYRINGA | 105,496 | 33,502 | 4.3% | 2.1% | ||
| Tiarella | X | X | ||||
| VACCINIUM | 49,533 | 20,200 | 2.0% | 1.3% | ||
| Veronica | X | X | ||||
| VIBURNUM | 45,960 | 14,009 | 1.9% | 0.9% | ||
| VINCA | 17,897 | 5506 | 0.7% | 0.4% | ||
| Vitis | X | X | ||||
| WEIGELA | 47,699 | 35,225 | 2.0% | 2.3% | ||
| total | 2,430,789 | 1,560,130 | ||||
References
- Lindtner, P. Garden Plants for Honey Bees; Wicwas Press, LLC.: Kalamazoo, MI, USA, 2014. [Google Scholar]
- Protecting Bees: Research to Help Plant Growers Protect Bees and Manage Pests. Available online: https://protectingbees.njaes.rutgers.edu/ (accessed on 2 June 2022).
- Mach, B.M.; Potter, D.A. Quantifying bee assemblages and attractiveness of flowering woody landscape plants for urban pollinator conservation. PLoS ONE 2018, 13, e0208428. [Google Scholar] [CrossRef] [PubMed]
- Garbuzov, M.; Ratnieks, F.L.W. Listmaina: The strengths and weaknesses of lists of garden plants to help pollinators. Bioscience 2014, 64, 1019–1026. [Google Scholar] [CrossRef]
- Garbuzov, M.; Ratnieks, F.L.W. Using the British National Collection of asters to compare the attractiveness of 228 varieties to flower-visiting insects. Environ. Entomol. 2015, 44, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Corbet, S.A.; Bee, J.; Dasmahapatra, K.; Gale, S.; Gorringe, E.; La Ferla, B.; Moorhouse, T.; Trevail, A.; Van Bergen, Y.; Vorontsova, M. Native or exotic? Double or single? Evaluating plants for pollinator-friendly gardens. Ann. Bot. 2001, 87, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Garbuzov, M.; Ratnieks, F.L.W. Quantifying variation among garden plants in attractiveness to bees and other flower-visiting insects. Funct. Ecol. 2014, 28, 364–374. [Google Scholar] [CrossRef]
- How Neonicotinoids Can Kill Bees. Available online: https://www.xerces.org/publications/scientific-reports/how-neonicotinoids-can-kill-bees (accessed on 16 June 2022).
- Mach, B.M.; Bondarenko, S.; Potter, D.A. Uptake and dissipation of neonicotinoid residues in nectar and foliage of systemically treated woody landscape plants. Environ. Toxicol. Chem. 2018, 37, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Cowles, R.S.; Eitzer, B.D. Residues of neonicotinoid insecticides in pollen and nectar from model plants. J. Environ. Hortic. 2017, 35, 24–34. [Google Scholar] [CrossRef]
- Stoner, K.A.; Cowles, R.S.; Nurse, A.; Eitzer, B.D. Tracking pesticide residues to a plant genus using palynology in pollen trapped from honey bees (Hymenoptera: Apidae) at ornamental plant nurseries. Environ. Entomol. 2019, 48, 351–362. [Google Scholar] [CrossRef]
- Erickson, E.; Patch, H.M.; Grozinger, C.M. Herbaceous perennial ornamental plants can support complex pollinator communities. Sci. Rep. 2021, 11, 17352. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1991. [Google Scholar]
- Couvillon, M.J.; Riddell, F.C.; Accleton, C.; Fensome, K.A.; Quah, S.K.L.; Taylor, E.L.; Ratnieks, F.L.W. Honey bee foraging distance depends on month and forage type. Apidologie 2015, 46, 61–70. [Google Scholar] [CrossRef]
- Wright, G.A.; Nicolson, S.W.; Shafir, S. Nutritional physiology and ecology of honey bees. Annu. Rev. Entomol. 2018, 63, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Lau, P.; Bryant Jr, V.M.; Ellis, J.D.; Huang, Z.Y.; Sullivan, J.; Schmehl, D.R.; Cabrera, A.R.; Rangel, J. Seasonal variation of pollen collecte by honey bees (Apis mellifera) in developed areas across fourregions in the United States. PLoS ONE 2019, 14, e0217294. [Google Scholar] [CrossRef]
- Bell, K.L.; De Vere, N.; Keller, A.; Richardson, R.T.; Gous, A.; Burgess, K.S.; Brosi, B.J. Pollen DNA barcoding: Current applications and future prospects. Genome 2016, 59, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Danner, N.; Grimmer, G.; von der Ankenbrand, M.; Von Der Ohe, K.; Von Der Ohe, W.; Rost, S.; Härtel, S.; Steffan-Dewenter, I. Evaluating multiplexed next-generation sequencing as a method in palynology for mixed pollen samples. Plant Biol. 2015, 17, 558–566. [Google Scholar] [CrossRef]
- Stoner, K.A.; Eitzer, B.D. Using a hazard quotient to evaluate pesticide residues detected in pollen trapped from honey bees (Apis mellifera) in Connecticut. PLoS ONE 2013, 8, e77550. [Google Scholar] [CrossRef]
- Long, E.Y.; Krupke, C.H. Non-cultivated plants present a season-long route of pesticide exposure for honey bees. Nature Commun. 2016, 7, 11629. [Google Scholar] [CrossRef]
- Smart, M.; Cornman, R.S.; Iwanowicz, D.D.; McDermott-Kubeczko, M.; Pettis, J.S.; Spivak, M.S.; Otto, C.R. A comparison of honey bee-collected pollen from working agricultural lands using light microscopy and ITS metabarcoding. Environ. Entomol. 2017, 46, 38–49. [Google Scholar] [CrossRef]
- McArt, S.H.; Fersch, A.A.; Milano, N.J.; Truitt, L.L.; Böröczky, K. High pesticide risk to honey bees despite low focal crop pollen collection during pollination of a mass blooming crop. Sci. Rep. 2017, 7, 46554. [Google Scholar] [CrossRef]
- Danner, N.; Keller, A.; Härtel, S.; Steffan-Dewenter, I. Honey bee foraging ecology: Season but not landscape diversity shapes the amount and diversity of collected pollen. PLoS ONE 2017, 12, e0183716. [Google Scholar] [CrossRef]
- Danner, N.; Härtel, S.; Steffan-Dewenter, I. Maize pollen foraging by honey bees in relation to crop area and landscape context. Basic Appl. Ecol. 2014, 15, 677–684. [Google Scholar] [CrossRef]
- Tosi, S.; Costa, C.; Vesco, U.; Quaglia, G.; Guido, G. A 3-year survey of Italian honey bee-collected pollen reveals widespread contamination by agricultural pesticides. Sci. Total Environ. 2018, 615, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Requier, F.; Odoux, J.-F.; Tamic, T.; Moreau, N.; Henry, M.; Decourtye, A.; Bretagnolle, V. Honey bee diet in intensive farmland habitats reveals an unexpectedly high flower richness and a major role of weeds. Ecol. Appl. 2015, 25, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Sponsler, D.B.; Grozinger, C.M.; Richardson, R.T.; Nurse, A.; Brough, D.; Patch, H.M.; Stoner, K.A. A screening-level assessment of the pollinator-attractiveness of ornamental nursery stock using a honey bee foraging assay. Sci. Rep. 2020, 10, 831. [Google Scholar] [CrossRef] [PubMed]
- US Census of Agriculture Highlights: Horticulture. Results from the 2019 Census of Horticultural Specialties. Available online: https://www.nass.usda.gov/Publications/Highlights/2020/census-horticulture.pdf (accessed on 6 June 2022).
- 2017 Census of Agriculture—State Data. Connecticut. Table 39. Floriculture and Bedding Crops, Nursery Crops, Propagative Materials Sold, Food Crops Grown Under Glass or Other Protection, and Mushroom Crops: 2017 and 2012. Available online: https://www.nass.usda.gov/Publications/AgCensus/2017/Full_Report/Volume_1_Chapter_1_State_Level/Connecticut/st09_1_0039_0039.pdf (accessed on 6 June 2022).
- Khachatryan, H.; Wei, X.; Rhih, A. Consumer and Producer Perceptions and Preferences of Pollinator-Friendly Labeling Practices in the US Green Industry; University of Florida, Intstitute of Food and Agricultural Sciences Extension: Gainesville, FL, USA, 2020. [Google Scholar]
- Richardson, R.T.; Lin, C.H.; Sponsler, D.B.; Quijia, J.O.; Goodell, K.; Johnson, R.M. Application of ITS2 metabarcoding to determine the provenance of pollen collected by honey bees in an agroecosystem. Appl. Plant Sci. 2015, 3, 1400066. [Google Scholar] [CrossRef]
- Richardson, R.T.; Eaton, T.D.; Lin, C.H.; Cherry, G.; Johnson, R.M.; Sponsler, D.B. Application of plant metabarcoding to identify diverse honeybee pollen forage along an urban-agricultural gradient. Mol. Ecol. 2021, 30, 310–323. [Google Scholar] [CrossRef]
- Bell, K.L.; Fowler, J.; Burgess, K.S.; Dobbs, E.K.; Gruenewald, D.; Lawley, B.; Morozumi, C.; Brosi, B.J. Applying pollen DNA metabarcoding to the study of plant-pollinator interactions. Appl. Plant Sci. 2017, 5, 1600124. [Google Scholar] [CrossRef]
- Bell, K.L.; Burgess, K.S.; Botsch, J.C.; Dobbs, E.K.; Read, T.D.; Brosi, B.J. Quantitative and qualitative assessment of pollen DNA metabarcoding using constructed species mixtures. Mol. Ecol. 2019, 28, 431–455. [Google Scholar] [CrossRef]
- O’Rourke, M.K.; Buchmann, S.L. Standardized analytical techniques for bee-collected pollen. Environ. Entomol. 1991, 20, 507–513. [Google Scholar] [CrossRef]
- Da Silveira, F. Influence of pollen grain volume on the estimation of the relative importance of its source to bees. Apidologie 1991, 22, 495–502. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H.; Buchmann, S.L. What governs protein content of pollen: Pollinator preferences, pollen-pistil interactions, or phylogeny? Ecol. Monogr. 2000, 70, 617–643. [Google Scholar] [CrossRef]
- Odoux, J.-F.; Feuillet, D.; Aupinel, P.; Loublier, Y.; Tasei, J.-N.; Mateescu, C. Territorial biodiversity and consequences on physico-chemical characteristics of pollen collected by honey bee colonies. Apidologie 2012, 43, 561–575. [Google Scholar] [CrossRef]
- Dewitz, J.; Survey, U.G. National Land Cover Datebase (NLCD) 2019 Products, Ver. 2.0, June 2021; US Geological Survey: Reston, VA, USA, 2021. [CrossRef]
- World Imagery. Available online: https://www.arcgis.com/home/item.html?id=10df2279f9684e4a9f6a7f08febac2a9 (accessed on 6 June 2022).
- Faegri, K.; Kaland, P.E.; Krzywinski, K. Textbook of Pollen Analysis; John Wiley & Sons Ltd.: New York, NY, USA, 1989. [Google Scholar]
- Crompton, C.W.; Wojtas, W.A. Pollen Grains of Canadian Honey Plants; Agriculture Canada and Canada Communication Group-Publishing: Ottawa, ON, Canada, 1993. [Google Scholar]
- McAndrews, J.H.; Berti, A.A.; Norris, G. Key to the Quaternary Pollen and Spores of the Great Lakes Region; Royal Ontario Museum: Toronto, ON, Canada, 2005; p. 65. [Google Scholar]
- Kapp, R.O.; King, J.E.; Davis, O.K. Ronald O. Kapp’s Pollen and Spores; American Association of Stratigraphic Palynologists Foundation Publication: Dallas, TX, USA, 2000. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord; Laboratoire de botanique historique et palynologie: Marseille, France, 1992. [Google Scholar]
- PalDat—A Palynological Database: Descriptions, Illustrations, Identification and Information Retrieval. Available online: https:/www.paldat.org (accessed on 7 July 2021).
- Human Impacts Pollen Database, an Illustrated Key. Available online: https://keyserver.lucidcentral.org/key-server/data/0f030b07-0200-4b0f-8509-0a0808060703/media/Html/Key.html (accessed on 7 July 2021).
- Haines, A. New England Wildflower Society’s Flora Novae Angliae: A Manual For the Identification of Native and Naturalized Higher Vascular Plants of New England; Yale University Press: New Haven, CT, USA; London, UK, 2011; p. 973. [Google Scholar]
- Wickham, H.; Grolemund, G. R for Data Science; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2017; p. 492. [Google Scholar]
- UConn Invasive Plant Factsheet: Multiflora Rose (Rosa multiflora). Available online: https://cipwg.uconn.edu/multiflora-rose/ (accessed on 24 May 2022).
- Sponsler, D.B.; Matcham, E.G.; Lin, C.-H.; Lanterman, J.L.; Johnson, R.M. Spatial and taxonomic patterns of honey bee foraging: A choice test between urban and agricultural landscapes. J. Urban Ecol. 2017, 3, juw008. [Google Scholar] [CrossRef]
- Zhang, G.; St. Clair, A.L.; Dolezal, A.; Toth, A.L.; O’Neal, M. Honey bee (Hymenoptera: Apidea) pollen forage in a highly cultivated agroecosystem: Limited diet diversity and its relationship to virus resistance. J. Econ. Entomol. 2020, 113, 1062–1072. [Google Scholar] [CrossRef]
- Krupke, C.H.; Hunt, G.J.; Eitzer, B.D.; Andino, G.; Given, K. Multiple routes of pesticide exposure for honey bees living near agricultural fields. PLoS ONE 2012, 7, e29268. [Google Scholar] [CrossRef] [PubMed]
- Keller, I.; Fluri, P.; Imdorf, A. Pollen nutrition and colony development in honey bees: Part 1. Bee World 2005, 86, 3–10. [Google Scholar] [CrossRef]
- Höcherl, N.; Siede, R.; Illies, I.; Gätschenberger, H.; Tautz, J. Evaluation of the nutritive value of maize for honey bees. J. Insect Physiol. 2012, 58, 278–285. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef]
- Lovell, J.H. The Flower and the Bee; Charles Scribner’s Sons: New York, NY, USA, 1918; p. 286. [Google Scholar]
- Cane, J.H. A brief review of monolecty in bees and benefits of a broadened definition. Apidologie 2021, 52, 17–22. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoner, K.A.; Nurse, A.; Koethe, R.W.; Hatala, M.S.; Lehmann, D.M. Where Does Honey Bee (Apis mellifera L.) Pollen Come from? A Study of Pollen Collected from Colonies at Ornamental Plant Nurseries. Insects 2022, 13, 744. https://doi.org/10.3390/insects13080744
Stoner KA, Nurse A, Koethe RW, Hatala MS, Lehmann DM. Where Does Honey Bee (Apis mellifera L.) Pollen Come from? A Study of Pollen Collected from Colonies at Ornamental Plant Nurseries. Insects. 2022; 13(8):744. https://doi.org/10.3390/insects13080744
Chicago/Turabian StyleStoner, Kimberly A., Andrea Nurse, Robert W. Koethe, Maxwell S. Hatala, and David M. Lehmann. 2022. "Where Does Honey Bee (Apis mellifera L.) Pollen Come from? A Study of Pollen Collected from Colonies at Ornamental Plant Nurseries" Insects 13, no. 8: 744. https://doi.org/10.3390/insects13080744
APA StyleStoner, K. A., Nurse, A., Koethe, R. W., Hatala, M. S., & Lehmann, D. M. (2022). Where Does Honey Bee (Apis mellifera L.) Pollen Come from? A Study of Pollen Collected from Colonies at Ornamental Plant Nurseries. Insects, 13(8), 744. https://doi.org/10.3390/insects13080744