
Rock Microhabitats Provide Suitable Thermal Conditions for Overwintering Insects: A Case Study of the Spongy Moth (Lymantria dispar L.) Population in the Altai Mountains



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Studies
2.2. Determination of the Viability of SM Eggs
2.3. Determination of the Supercooling Point (SCP)
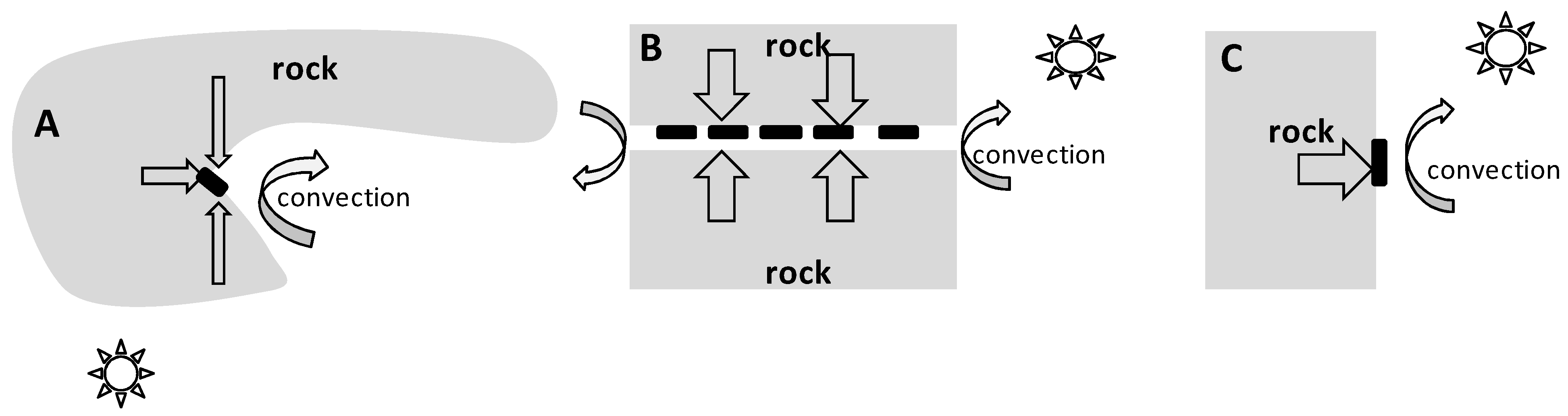
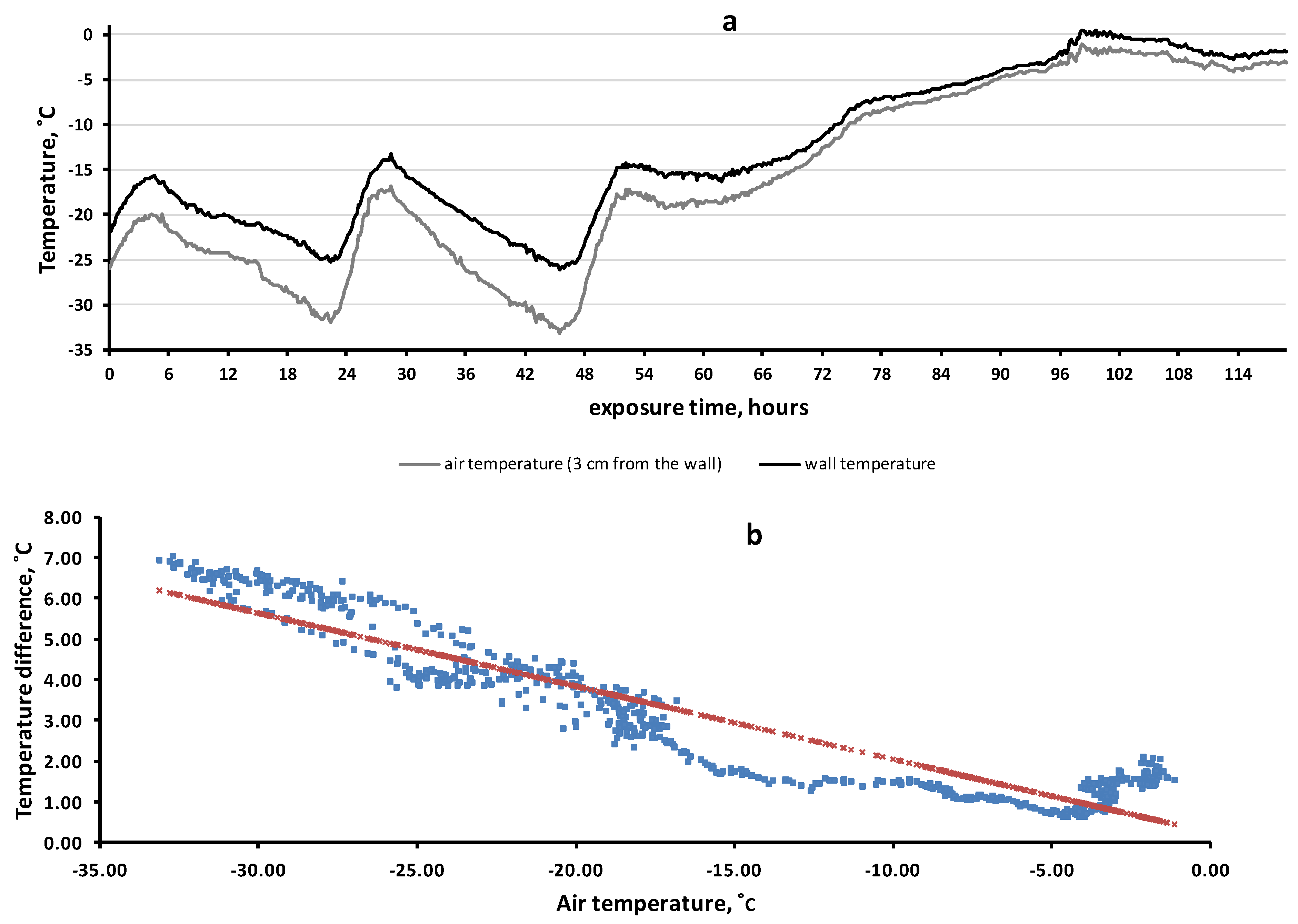
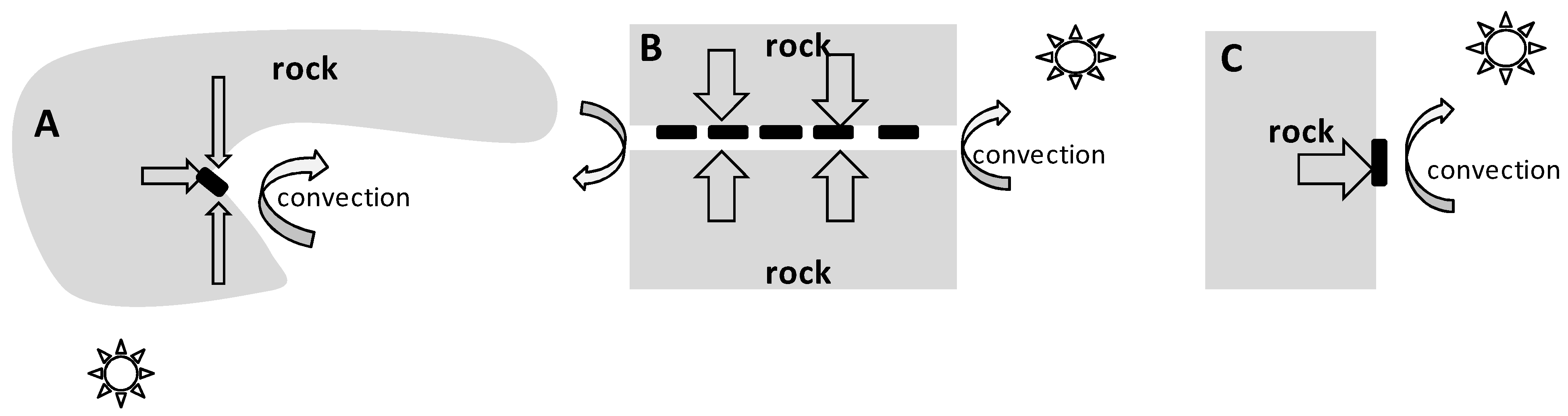
2.4. Rock Microhabitat Simulation Experiment
2.5. Statistical Analysis of the Results
3. Results
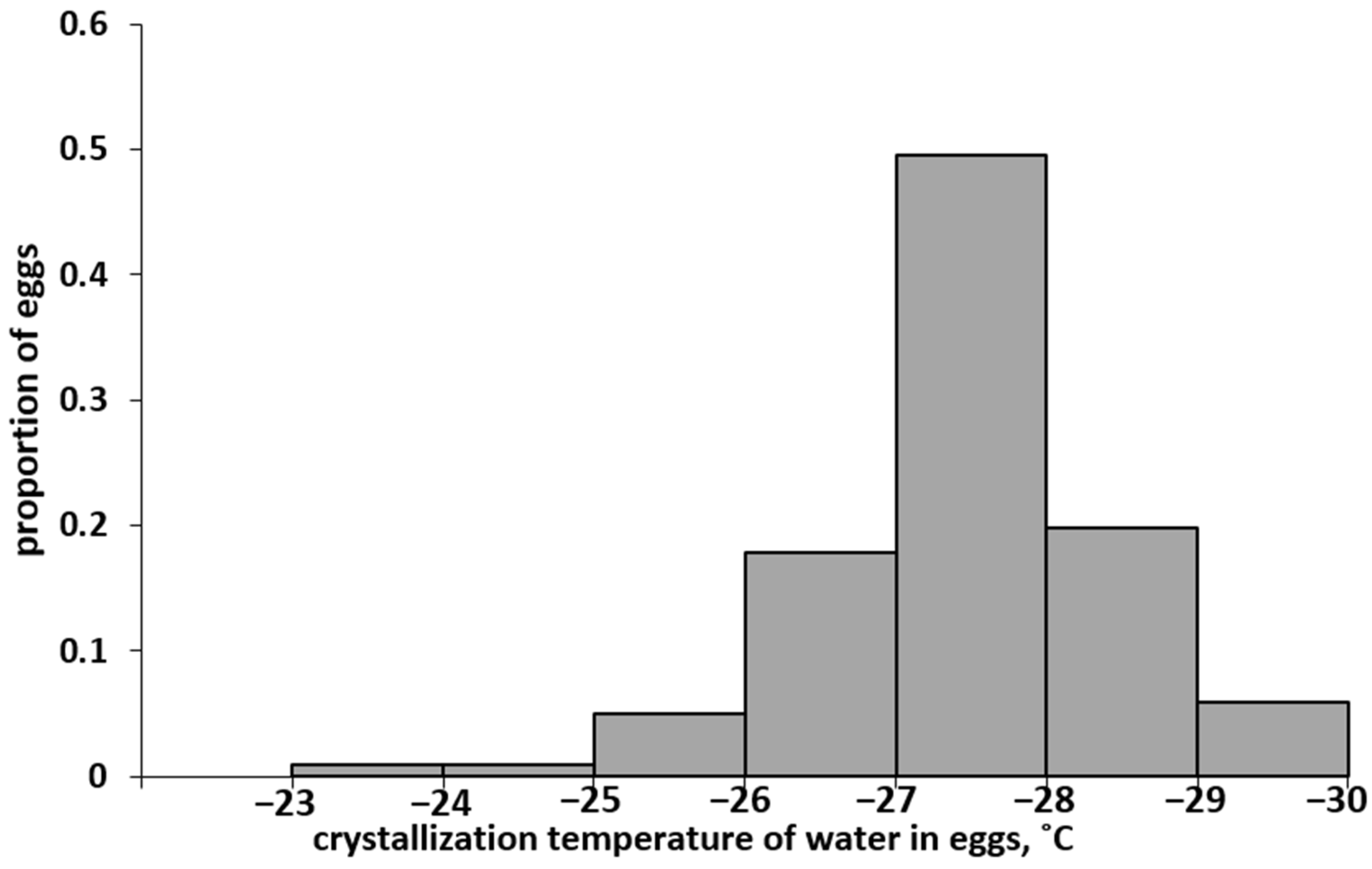
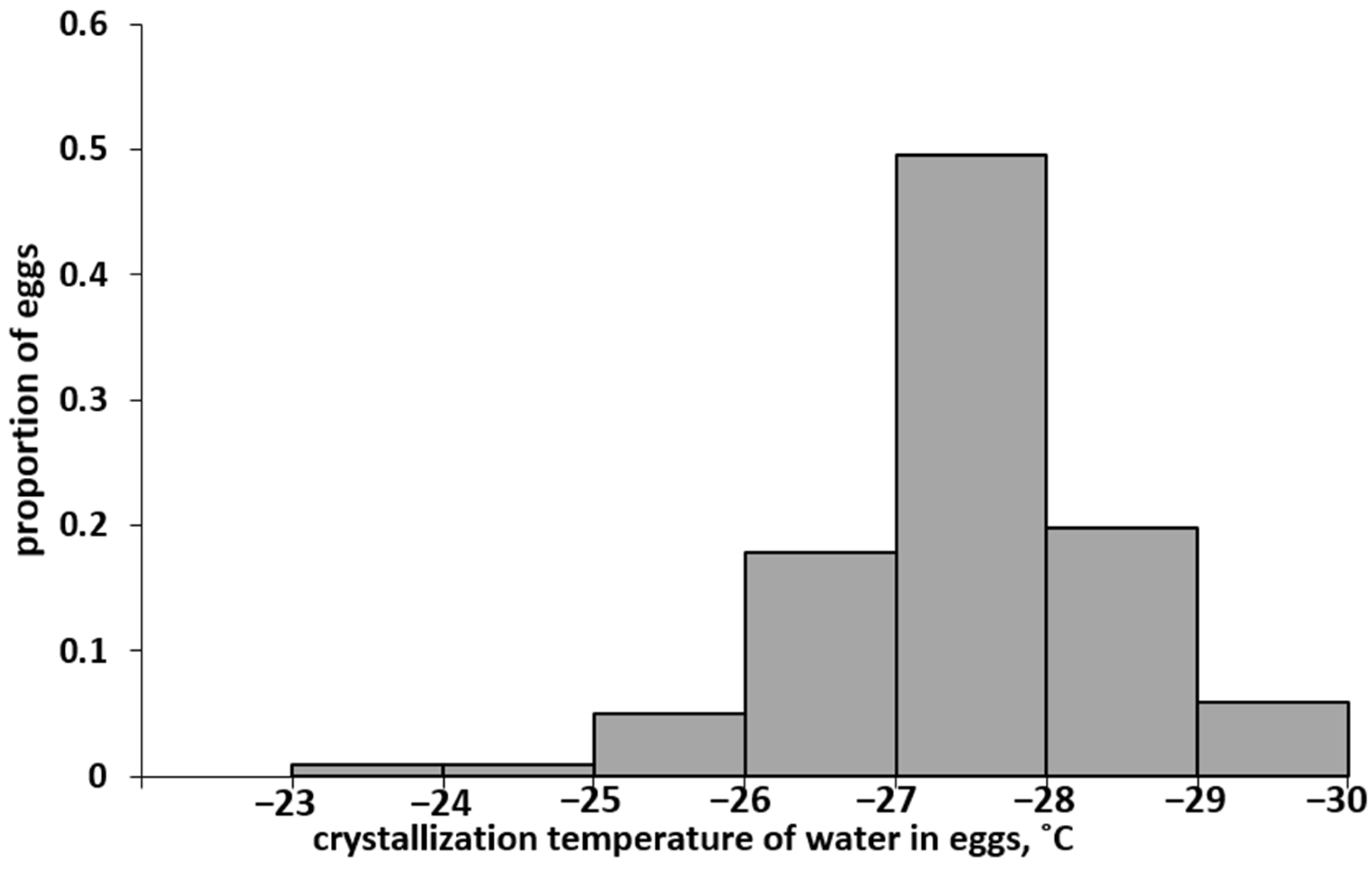
3.1. Distribution Spectrum of Eggs of the Altai Population by SCP
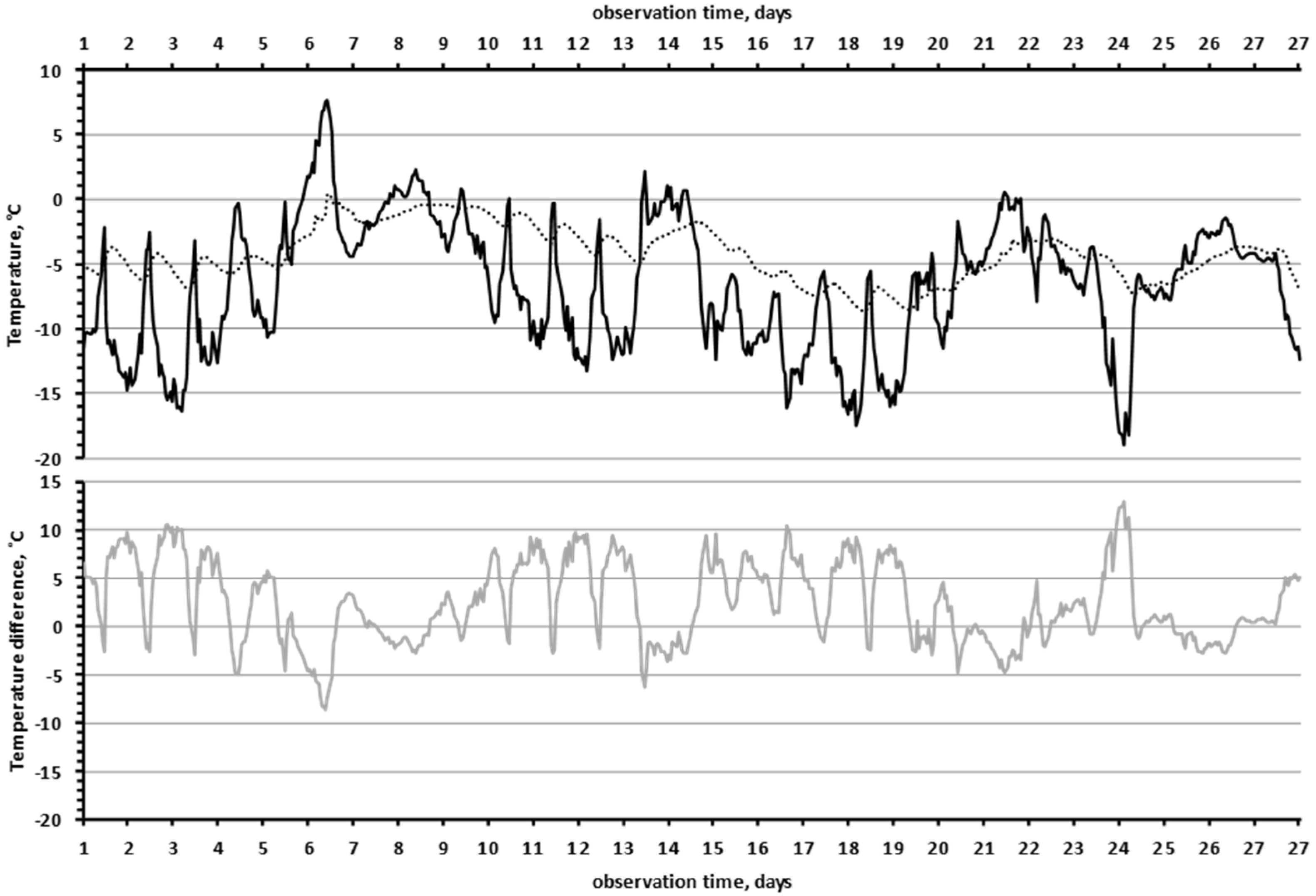
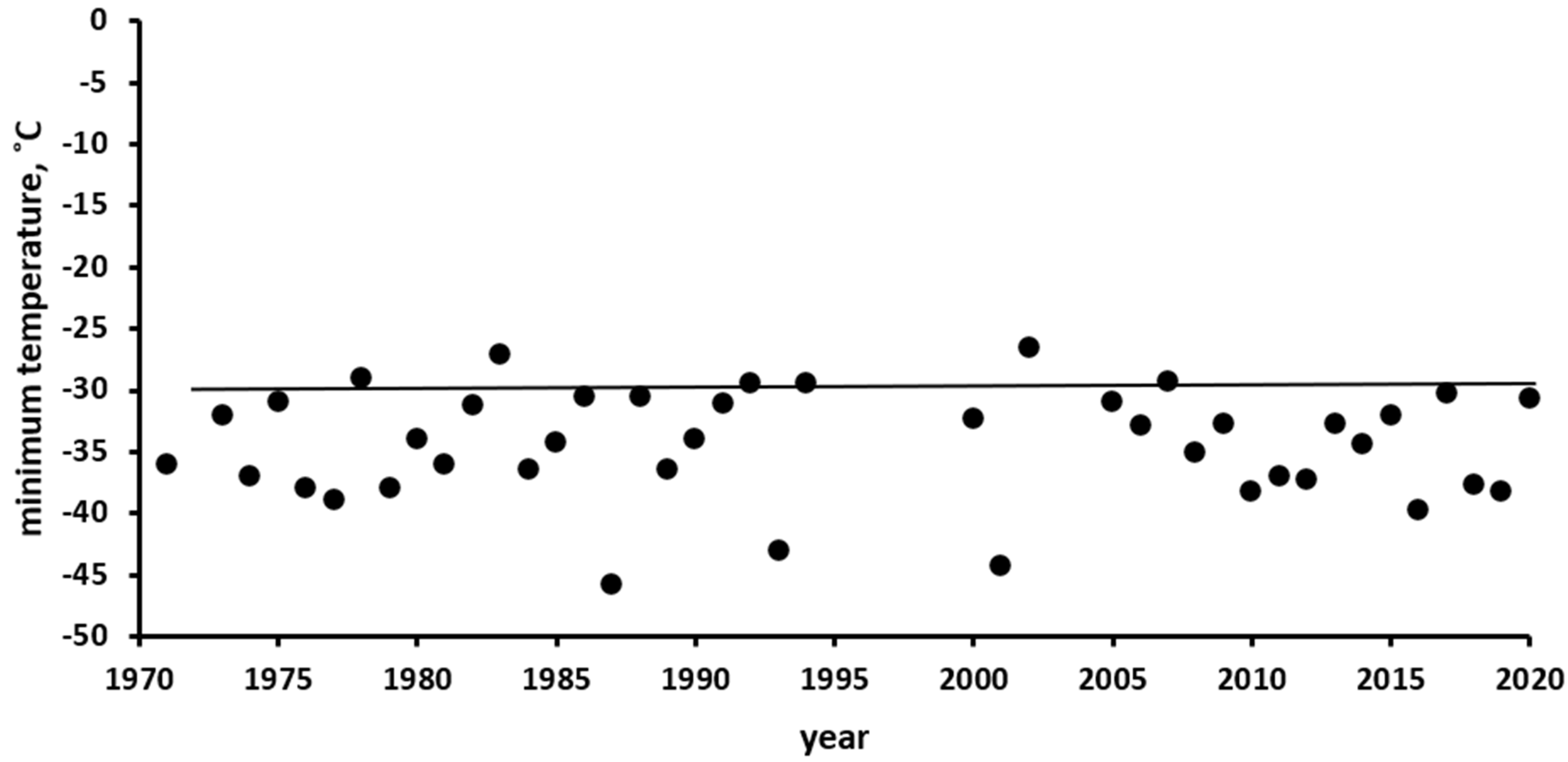
3.2. Characteristics of the Climate in the Observation Area
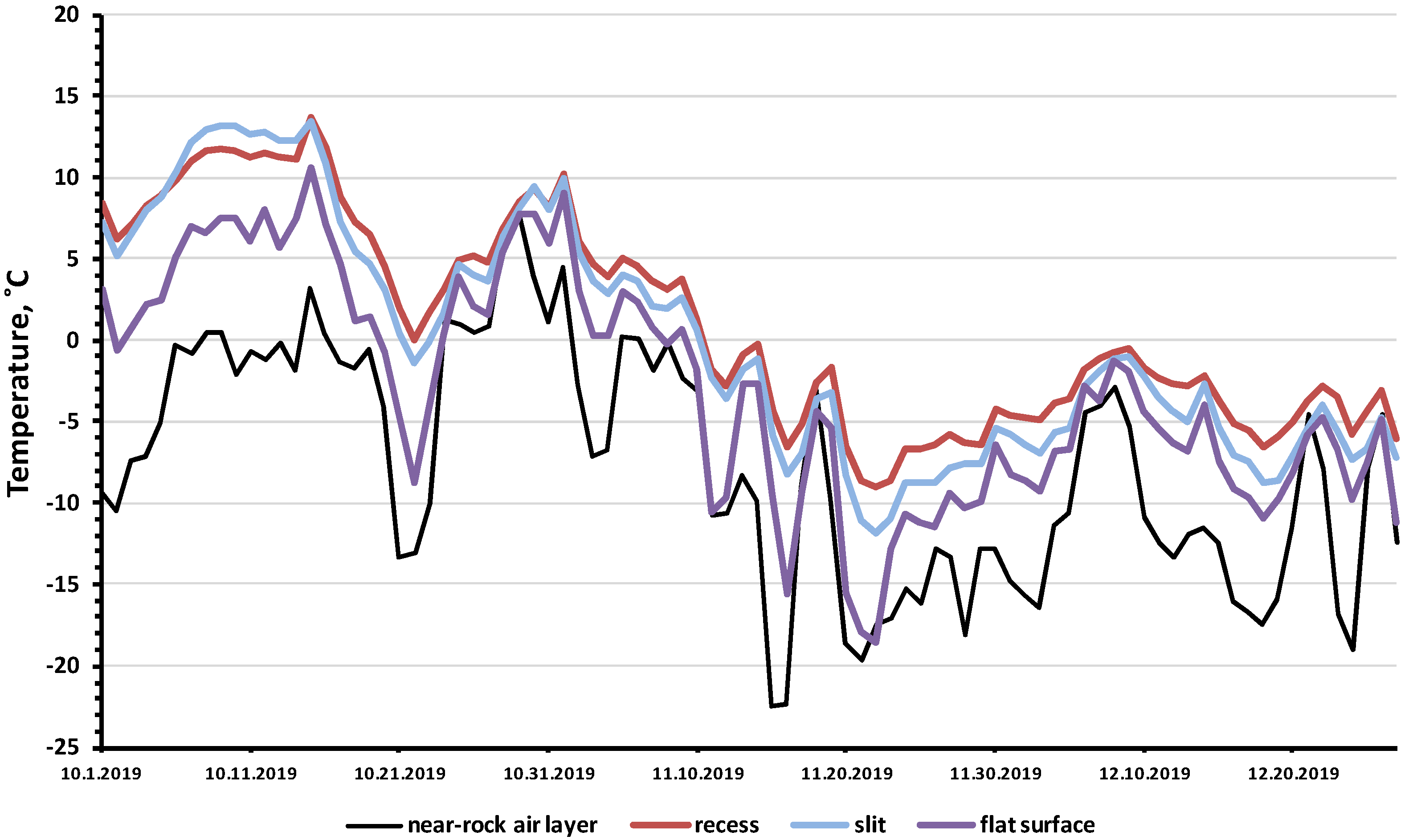
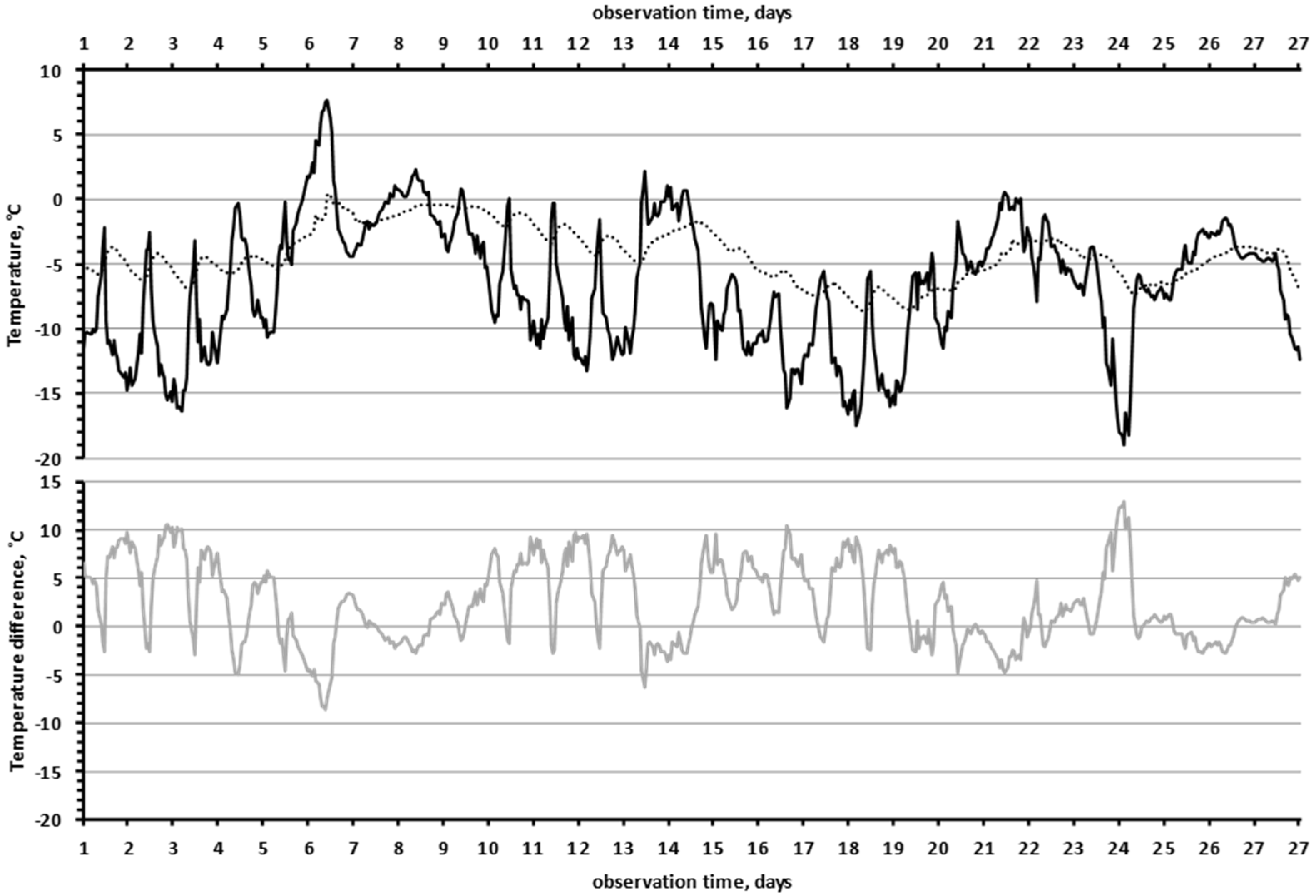
3.3. Comparison of Air Temperature and Temperature in Rock Microhabitats
3.4. Viability of Overwintered SM Eggs
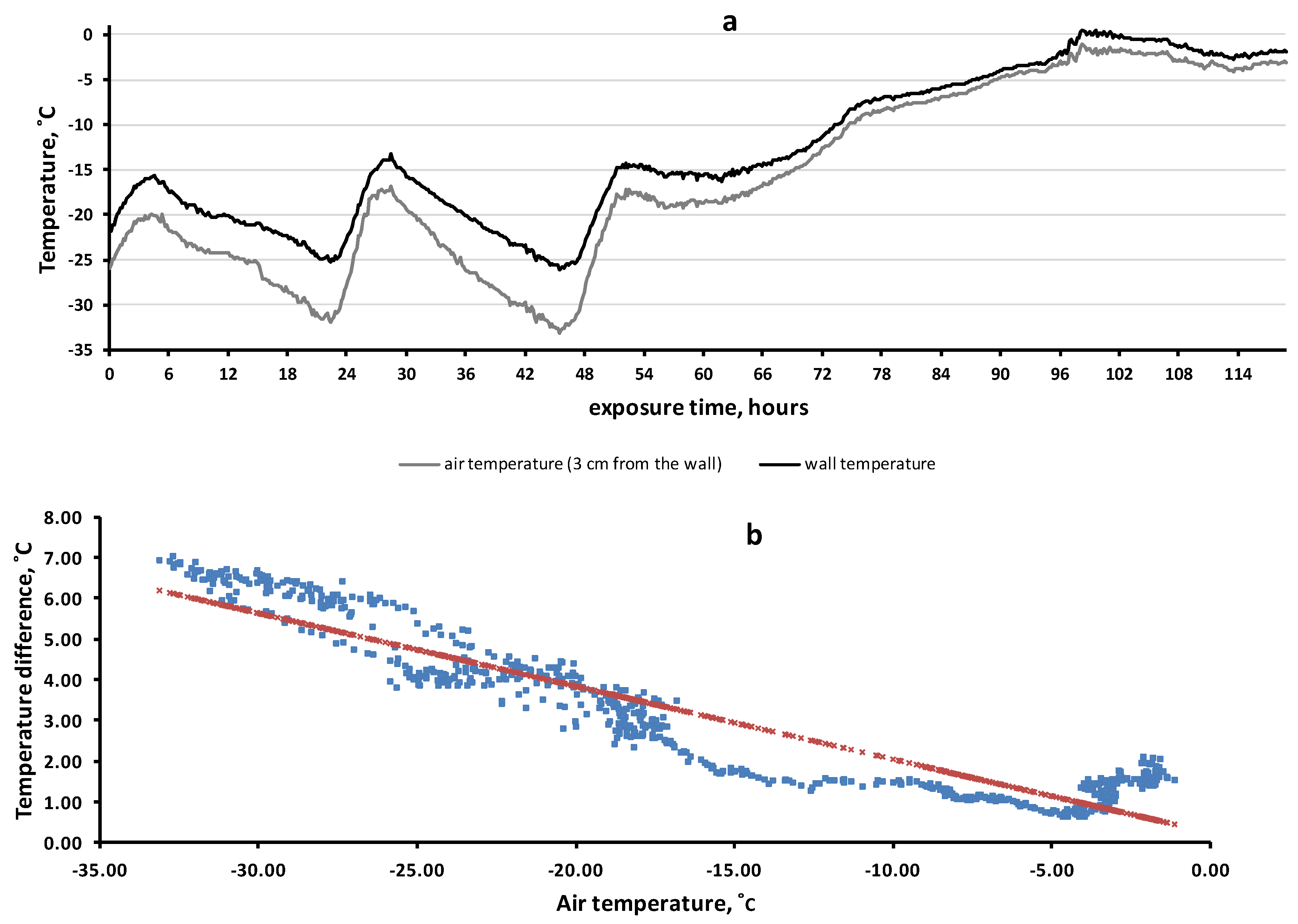
3.5. Rock Microhabitat Simulation Experiment
4. Discussion
4.1. Climate and Microclimate in the Research Area
4.2. The Relationship of Parental Behaviour of the SM with the Climate and Landscape of the Region
4.3. On the Mechanisms of “Warming” of Rock Microhabitats
4.4. Advantages of Rock Microhabitats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, R.E., Jr. Insects at Low Temperature; Lee, R.E., Jr., Denlinger, D.L., Eds.; Chapman and Hall: New York, NY, USA, 1991; pp. 17–46. [Google Scholar] [CrossRef]
- Leather, S.R.; Walters, K.F.A.; Bale, J.S. The Ecology of Insect Overwintering; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar] [CrossRef]
- Williams, C.M.; Henry, H.A.L.; Sinclair, B.J. Cold truths: How winter drives responses of terrestrial organisms to climate change. Biol. Rev. 2015, 90, 214–235. [Google Scholar] [CrossRef] [PubMed]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986; ISBN 0195036352. [Google Scholar]
- Teets, N.M.; Denlinger, D.L. Surviving in a frozen desert: Environmental stress physiology of terrestrial Antarctic arthropods. J. Exp. Biol. 2014, 217, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S. Insects and low temperatures: From molecular biology to distributions and abundance. Philos. Trans. R Soc. Lond. B Biol. Sci. 2002, 357, 849–862. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [PubMed]
- Danks, H.V. Winter habitats and ecological adaptations for winter survival. In Insects at Low Temperature; Lee, R.E., Jr., Denlinger, D.L., Eds.; Chapman and Hall: New York, NY, USA, 1991; pp. 231–259. [Google Scholar] [CrossRef]
- Danks, H.V. Modification of adverse conditions by insects. Oikos 2002, 99, 10–24. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J. Field ecology of freeze tolerance: Interannual variation in cooling rates, freeze-thaw and thermal stress in the microhabitat of the alpine cockroach Celatoblatta quinquemaculata. Oikos 2001, 93, 286–293. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Lord, J.M.; Thompson, C.M. Microhabitat selection and seasonality of alpine Invertebrates. Pedobiologia 2002, 45, 107–120. [Google Scholar] [CrossRef]
- Bennett, V.A.; Lee, R.E.; Nauman, J.S.; Kukal, O. Selection of overwintering microhabitats used by the arctic woolybear caterpillar Gynaephora groenlandica. CryoLetters 2003, 24, 191–200. Available online: https://pubmed.ncbi.nlm.nih.gov/12908029/ (accessed on 1 January 2022).
- Hodkinson, I.D. Adaptations of invertebrates to terrestrial Arctic environments. In The Royal Norwegian Society of Sciences and Letters; Tapir Academic Press: Trondheim, Norway, 2005. [Google Scholar]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess Drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Srivastava, V.; Verena, G.C.; Keena, M.A. Assessing the Potential Distribution of Asian Gypsy Moth in Canada: A Comparison of Two Methodological Approaches. Sci. Rep. 2020, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Elkinton, J.S.; Liebhold, A.M. Population dynamics of gypsy moth in North America. Annu. Rev. Entomol. 1990, 35, 571–596. [Google Scholar] [CrossRef]
- Grayson, K.L.; Johnson, D.M. Novel insights on population and range edge dynamics using an unparalleled spatiotemporal record of species invasion. J. Anim. Ecol. 2018, 87, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Pogue, M.G.; Schaefer, P.W. A Review of Selected Species of Lymantria Hubner [1819] (Lepidoptera: Noctuidae: Lymantriinae) from Subtropical and Temperate Regions of Asia, Including the Descriptions of Three New Species, Some Potentially Invasive to North America; FHTET: Washington, DC, USA, 2007; Available online: http://www.fs.fed.us/foresthealth/technology/ (accessed on 1 January 2022).
- Ponomarev, V.I.; Ilinykh, A.V.; Gninenko, J.I.; Sokolov, G.I.; Andreeva, E.M. Gypsy Moth in Trans-Ural and Western Siberia; Ural Branch of Russian Academy of Science: Yekaterinburg, Russia, 2012; ISBN 978-5-7691-2251-4. (In Russian) [Google Scholar]
- Leonard, D.E. Recent developments in ecology and control of the gypsy moth. Annu. Rev. Entomol. 1974, 19, 197–229. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Lee, R.E.; Yocum, G.D.; Kukal, O. Role of chilling in the acquisition of cold tolerance and the capacitation to express stress proteins in diapausing pharate larvae of the Gypsy Moth, Lymantria dispar. Arch. Insect Biochem. Physiol. 1992, 21, 271–280. [Google Scholar] [CrossRef]
- Sullivan, C.R.; Wallace, D.R. The potential northern dispersal of the gypsy moth, Portheria dispar (Lepidoptera: Lymantriidae). Can. Entomol. 1972, 104, 1349–1355. [Google Scholar] [CrossRef]
- Madrid, F.J.; Stewart, R.K. Ecological significance of cold hardiness and winter mortality of eggs of the gypsy moth Lymantria dispar L. in Quebec. Environ. Entomol. 1981, 10, 586–589. [Google Scholar] [CrossRef]
- Streifel, M.A.; Tobin, P.C.; Kees, A.M.; Aukema, B.H. Range expansion of Lymantria dispar dispar (L.) (Lepidoptera: Erebidae) along its north-western margin in North America despite low predicted climatic suitability. J. Biogeogr. 2019, 46, 58–69. [Google Scholar] [CrossRef]
- Fält-Nardmann, J.J.J.; Ruohomäki, K.; Tikkanen, O.-P.; Neuvonen, S. Cold hardiness of Lymantria monacha and L. dispar (Lepidoptera: Erebidae) eggs to extreme winter temperatures: Implications for predicting climate change impacts. Ecol. Entomol. 2018, 43, 422–430. [Google Scholar] [CrossRef]
- Pantyukohov, G.A. The Effect of negative temperatures upon different populations of Euproctis chrysorrhea L. and Lymantria dispar L. (Lepidoptera, Orgyidae). Entomol. Obozr. 1964, 43, 94–111. (In Russian) [Google Scholar]
- Ananko, G.G.; Kolosov, A.V. Asian gypsy moth (Lymantria dispar L.) populations: Tolerance of eggs to extreme winter temperatures. J. Therm. Biol. 2021, 102, 103123. [Google Scholar] [CrossRef] [PubMed]
- Kurenshchikov, D.K.; Martemyanov, V.V.; Imranova, E.L. Features of the far eastern gypsy moth (Lymantria dispar L.) population outbreak. Contemp. Probl. Ecol. 2020, 13, 172–179. [Google Scholar] [CrossRef]
- Benkevich, V.I. The distribution of gypsy moth (Porthetria dispar L.) egg masses over territory of the mountain Altai. Zool. Xhurnal 1956, 35, 1013–1016. (In Russian) [Google Scholar]
- Hauck, M.; Dulamsuren, C.H.; Heimes, C. Effects of a gypsy moth invasion on the performance of Larix sibirica in a forest-steppe ecotone of northern Mongolia. Environ. Exp. Bot. 2008, 62, 351–356. [Google Scholar] [CrossRef]
- Kondakov, Y.P. The gypsy moth (Ocneria dispar L.) in the forests of the Krasnoyarsk Territory. Prot. Sib. For. Insect Pests 1963, 1, 30–77. (In Russian) [Google Scholar]
- Inoue, M.N.; Suzuki-Ohno, Y.; Haga, Y.; Aarai, H.; Sano, T.; Martemyanov, V.V.; Kunimi, Y. Population dynamics and geographical distribution of the gypsy moth, Lymantria dispar, in Japan. For. Ecol. Manag. 2019, 434, 154–164. [Google Scholar] [CrossRef]
- Coulson, S.J.; Hodkinson, I.D.; Strathdee, A.T.; Block, W.; Webb, N.R.; Bale, J.S.; Worland, M.R. Thermal Environments of Arctic Soil Organisms during Winter. Arct. Alp. Res. 1995, 27, 364–370. [Google Scholar] [CrossRef]
- Smitley, D.; Andresen, J.; Priest, R.; Mech, R.; Mccullough, D. Winter mortality of gypsy moth (Lepidoptera: Lymantriidae) eggs in Michigan. Environ. Entomol. 1998, 27, 700–708. [Google Scholar] [CrossRef]
- Yurchenko, G.I.; Turova, G.I. Distribution of gypsy moth egg masses in deciduous and mixed forests in the Far East. In IX s’yezd Vsesoyuznogo Entomologicheskogo Obshchestva; Naukova Dumka: Kiev, Ukraine, 1984; Tezisy dokladov. (In Russian) [Google Scholar]
- Ciancio, J.J.; Turnbull, K.F.; Gariepy, T.D.; Sinclair, B.J. Cold tolerance, water balance, energetics, gas exchange, and diapause in overwintering brown marmorated stink bugs. J. Insect Physiol. 2021, 128, 104171. [Google Scholar] [CrossRef]
- Conver, J.L.; Yarwood, E.; Hetherington, L.D.; Swann, D.E. Nurse rock microclimates significantly buffer exposure to freezing temperature and moderate summer temperature. J. Arid. Environ. 2020, 177, 104140. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology; Oxford Univ. Press: Oxford, UK, 2005. [Google Scholar]
- Millar, C.I.; Westfall, R.D.; Delany, D.L. Thermal regimes and snowpack relations of periglacial talus slopes, Sierra Nevada, California, USA. Arct. Antarct. Alp. Res. 2014, 46, 483–504. [Google Scholar] [CrossRef]
- Midttømme, K.; Roaldset, E. Thermal conductivity of sedimentary rocks: Uncertainties in measurement and modelling. Geol. Soc. Lond. Spec. Publ. 1999, 158, 45–60. [Google Scholar] [CrossRef]
- Beirão, S.G.S.; Ribeiro, A.P.C.; Lourenço, M.J.V.; Santos, F.J.V.; Nieto de Castro, C.A. Thermal Conductivity of Humid Air. Int. J. Thermophys. 2012, 33, 1686–1703. [Google Scholar] [CrossRef]
- Jury, W.A.; Bellantuoni, B. Heat and water movement under surface rocks in a field soil: I. Thermal effects. Soil Sci. Soc. Am. J. 1976, 40, 505–509. [Google Scholar] [CrossRef]
- Goldsrough, C.L.; Hochuli, D.F.; Shine, R. Fitness benefits of retreat-site selection: Spiders, rocks, and thermal cues. Ecology 2004, 85, 1635–1641. [Google Scholar] [CrossRef]
- Van den Berg, F.T.; Thompson, M.B.; Hochuli, D.F. When hot rocks get hotter: Behavior and acclimatization mitigate exposure to extreme temperatures in a spider. Ecosphere 2015, 6, 88. [Google Scholar] [CrossRef]
- Huey, R.B.; Peterson, C.R.; Arnold, S.J.; Porter, W.P. Hot Rocks and Not-So-Hot Rocks: Retreat-Site Selection by Garter Snakes and Its Thermal Consequences. Ecology 1989, 70, 931–944. [Google Scholar] [CrossRef]
- Hoshikawa, K.; Tsutsui, H.; Honma, K.; Sakagami, S.F. Cold resistance in four species of beetles overwintering in the soil, with notes on the overwintering strategies of some soil insects. Appl. Entomol. Zool. 1988, 23, 273–281. [Google Scholar] [CrossRef]
- Berman, D.I.; Alfimov, A.V.; Zhigulskaya, Z.A.; Leirikh, A.N. Overwintering and Cold-Hardiness of Ants in the Northeast of Asia; Pensoft: Sofia, Bulgaria; Moscow, Russia, 2010; p. 294. ISBN 9789546425492. [Google Scholar]
- Convey, P.; Abbandonato, H.; Bergan, F.; Beumer, L.T.; Biersma, E.M.; Bråthen, V.S.; D’Imperio, L.; Jensen, C.K.; Nilsen, S.; Paquin, K.; et al. Survival of rapidly fluctuating natural low winter temperatures by High Arctic soil invertebrates. J. Therm. Biol. 2015, 54, 111–117. [Google Scholar] [CrossRef]
- Barry-Macaulay, D.; Bouazza, A.; Wang, B.; Singh, R.M. Evaluation of soil thermal conductivity models. Can. Geotech. J. 2015, 52, 1892–1900. [Google Scholar] [CrossRef]
- He, H.; Liu, L.; Dyck, M.; Si, B.; Lv, J. Modelling dry soil thermal conductivity. Int. Commun. Heat Mass Transf. 2020, 116, 104602. [Google Scholar] [CrossRef]
- Srivastava, V.; Keena, M.A.; Maennicke, G.E.; Hamelin, R.C.; Griess, V.C. Potential Differences and Methods of Determining Gypsy Moth Female Flight Capabilities: Implications for the Establishment and Spread in Novel Habitats. Forests 2021, 12, 103. [Google Scholar] [CrossRef]
- Humble, L.M.; Allen, E.A. Forest biosecurity: Alien invasive species and vectored organisms. Can. J. Plant Pathol. 2006, 28, S256–S269. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Months | Parameters | Sites | |||||
|---|---|---|---|---|---|---|---|
| Air—Weather Station | Air—Near-Rock Layer | Rock—“Recess” | Rock—“Slit” | Rock—“Flat Surface” | Rock *** | ||
| October | Mean | 2.1 | 5.3 | 9.4 | 9.2 | 8.4 | 9.0 |
| Difference of averages * | −3.2 | - | 4.1 | 3.9 | 3.1 | 3.7 | |
| Maximum | 21.5 | 29.5 | 15.2 | 17.5 | 37.9 | 23.5 | |
| Minimum | −20.3 | −13.3 | 0.0 | −1.4 | −8.8 | −3.4 | |
| Fluctuation amplitude ** | 41.8 | 42.8 | 15.2 | 18.9 | 46.7 | 26.9 | |
| November | Mean | −8.9 | −5.8 | −0.8 | −1.9 | −3.0 | −1.9 |
| Difference of averages * | −3.1 | - | 5.0 | 3.9 | 2.8 | 3.9 | |
| Maximum | 12.5 | 15.7 | 11.3 | 11.2 | 20.4 | 14.3 | |
| Minimum | −32.2 | −22.5 | −9.0 | −11.8 | −18.5 | −13.1 | |
| Fluctuation amplitude ** | 44.7 | 38.2 | 20.3 | 23.0 | 38.9 | 27.4 | |
| December | Mean | −10.3 | −6.6 | −3.3 | −4.2 | −5.0 | −4.2 |
| Difference of averages * | −3.7 | - | 3.0 | 2.4 | 1.6 | 2.3 | |
| Maximum | 6.6 | 7.6 | 0.4 | 2.3 | 8.4 | 3.7 | |
| Minimum | −22.7 | −19.0 | −8.9 | −8.7 | −13.7 | −10.4 | |
| Fluctuation amplitude ** | 29.3 | 26.6 | 9.3 | 11.0 | 22.1 | 14.1 | |
| Sites | |||||
|---|---|---|---|---|---|
| Air—Near-Rock Layer | Rock—“Recess” | Rock—“Slit” | Rock—“Flat Surface” | Rock * | |
| Average minimal temperature | −10.7 | −2.7 | −4.1 | −6.5 | −4.4 |
| Temperature difference | - | 8.0 | 6.6 | 4.2 | 6.3 |
| p-value ** | U = 14, p = 6.42 × 10−7 | U = 32, p = 2.342 × 10−5 | U = 68, p = 0.0012 | U = 46, p = 0.00036 | |
| Parameters | Population before Wintering | After Wintering—Southern Slope | After Wintering—Eastern Slope | ||
|---|---|---|---|---|---|
| “Flat surface” | “Slit” | “Recess” | “Deep Recess” | ||
| Proportion of viable eggs *, % | 91 ± 2 | 84 ± 7 | 88 ± 3 | 79 ± 6 | 82 ± 5 |
| Time before the beginning of hatching, hour | 163 ± 7 | 14 ± 2 | 27 ± 2 | 22 ± 1 | 91 ± 3 |
| p-value ** | 8.72 × 10−7 | 2.48 × 10−7 | 6.76 × 10−7 | 1.26 × 10−7 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ananko, G.G.; Kolosov, A.V.; Martemyanov, V.V. Rock Microhabitats Provide Suitable Thermal Conditions for Overwintering Insects: A Case Study of the Spongy Moth (Lymantria dispar L.) Population in the Altai Mountains. Insects 2022, 13, 712. https://doi.org/10.3390/insects13080712
Ananko GG, Kolosov AV, Martemyanov VV. Rock Microhabitats Provide Suitable Thermal Conditions for Overwintering Insects: A Case Study of the Spongy Moth (Lymantria dispar L.) Population in the Altai Mountains. Insects. 2022; 13(8):712. https://doi.org/10.3390/insects13080712
Chicago/Turabian StyleAnanko, Grigory G., Aleksei V. Kolosov, and Vyacheslav V. Martemyanov. 2022. "Rock Microhabitats Provide Suitable Thermal Conditions for Overwintering Insects: A Case Study of the Spongy Moth (Lymantria dispar L.) Population in the Altai Mountains" Insects 13, no. 8: 712. https://doi.org/10.3390/insects13080712
APA StyleAnanko, G. G., Kolosov, A. V., & Martemyanov, V. V. (2022). Rock Microhabitats Provide Suitable Thermal Conditions for Overwintering Insects: A Case Study of the Spongy Moth (Lymantria dispar L.) Population in the Altai Mountains. Insects, 13(8), 712. https://doi.org/10.3390/insects13080712