
Back to the Wild: The Parasitoid Community of Lobesia botrana (Lepidoptera: Tortricidae) in a Grapevine-Free Natural Environment




 and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
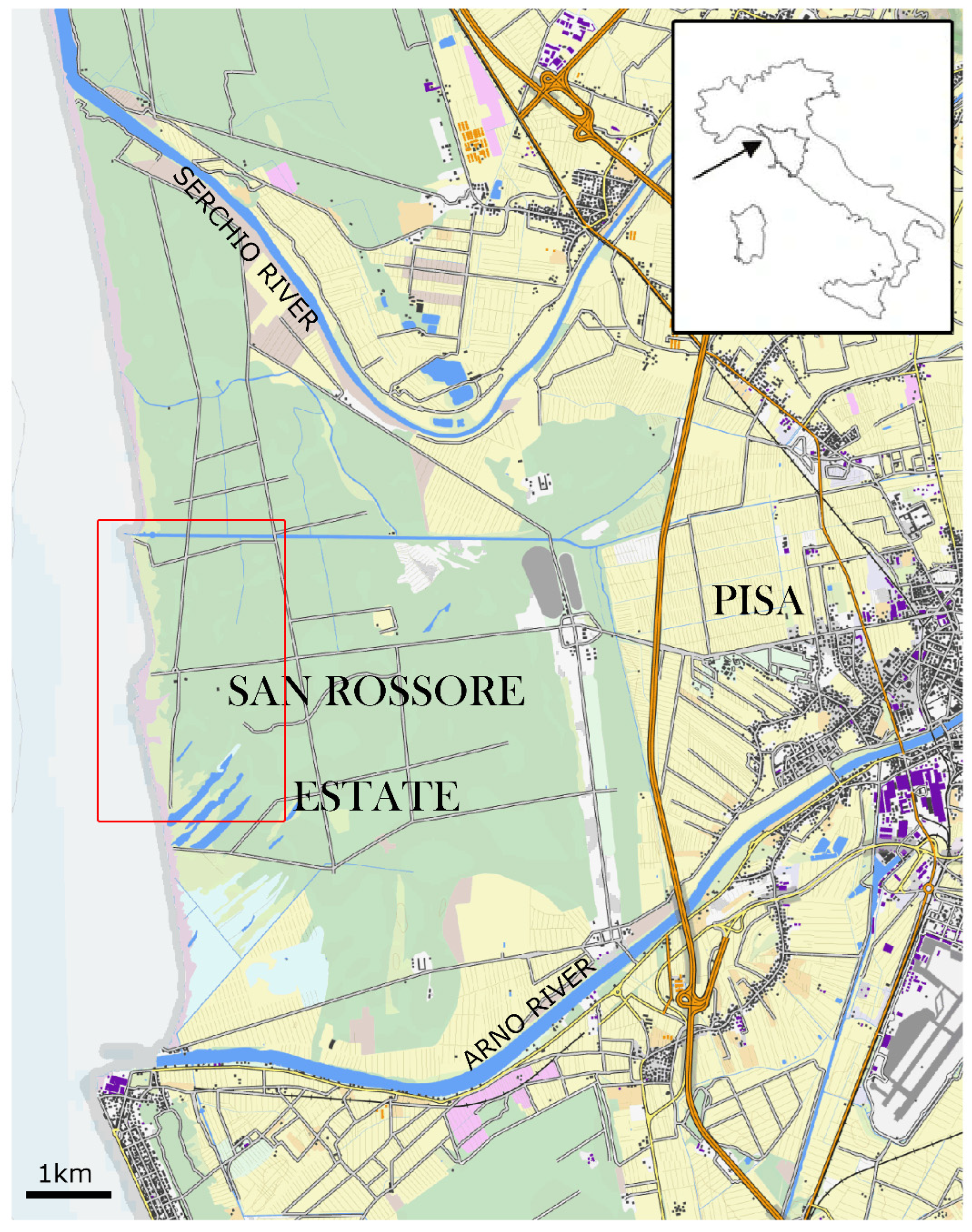
2.1. Study Area
2.2. Sampling Design
2.3. Host Parasitisation Rate and Statistical Analysis
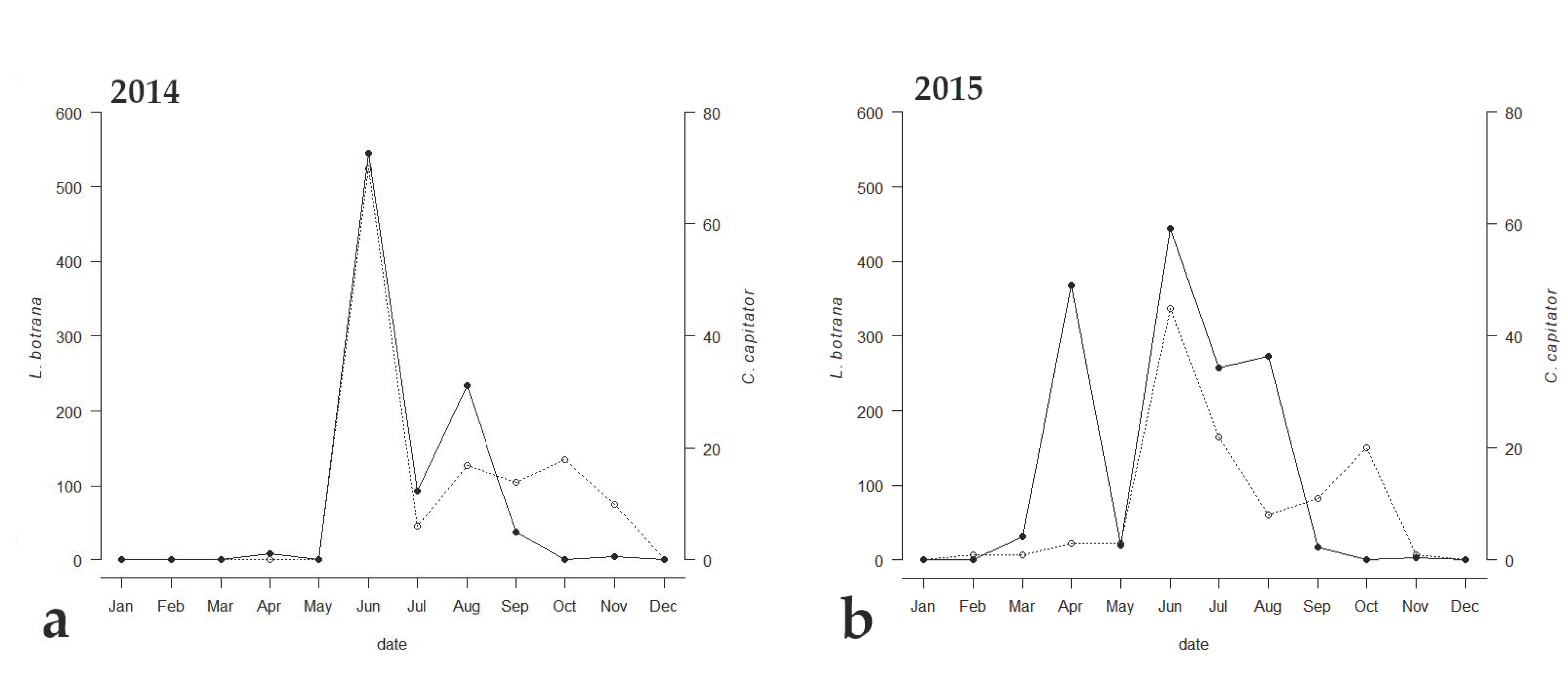
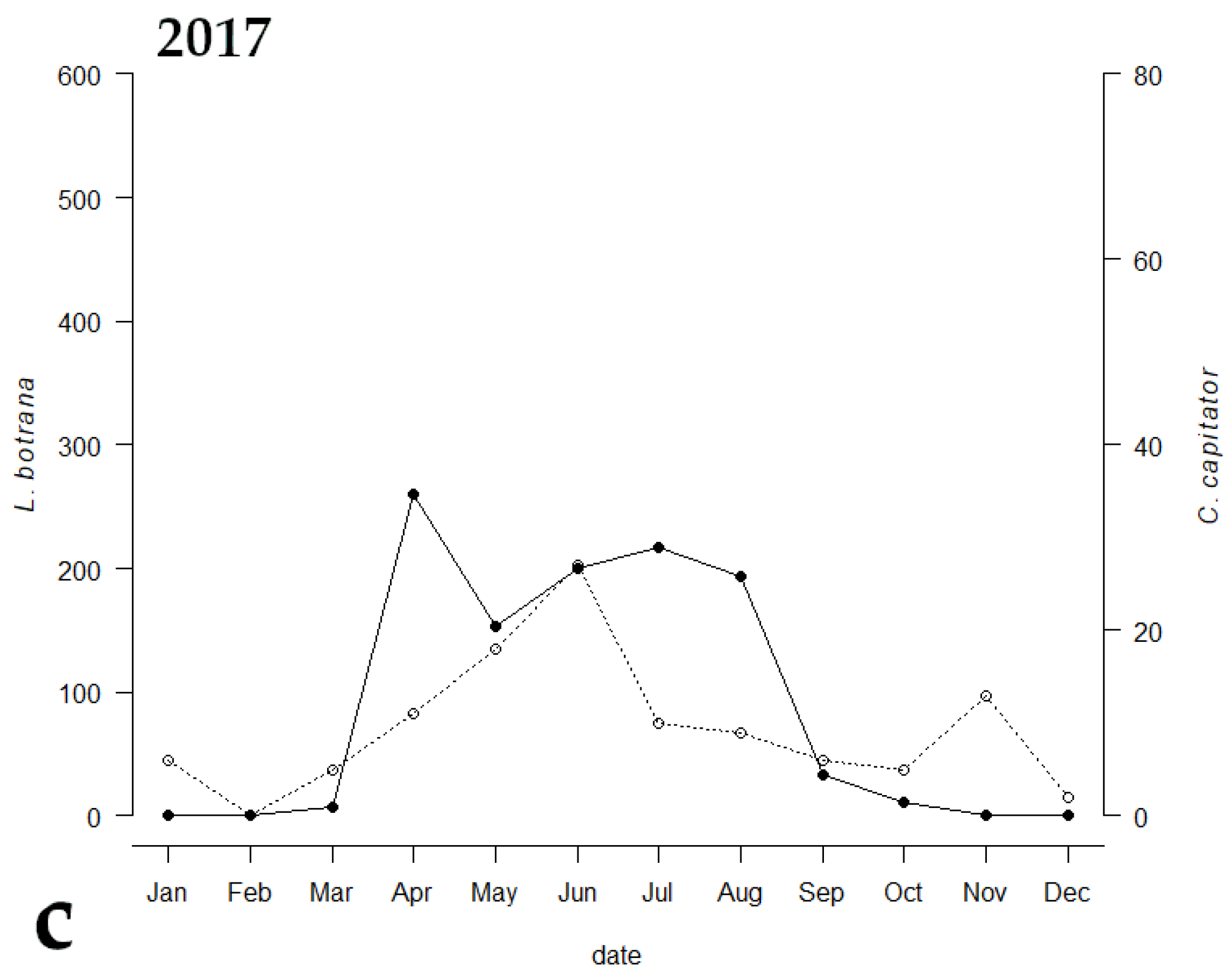
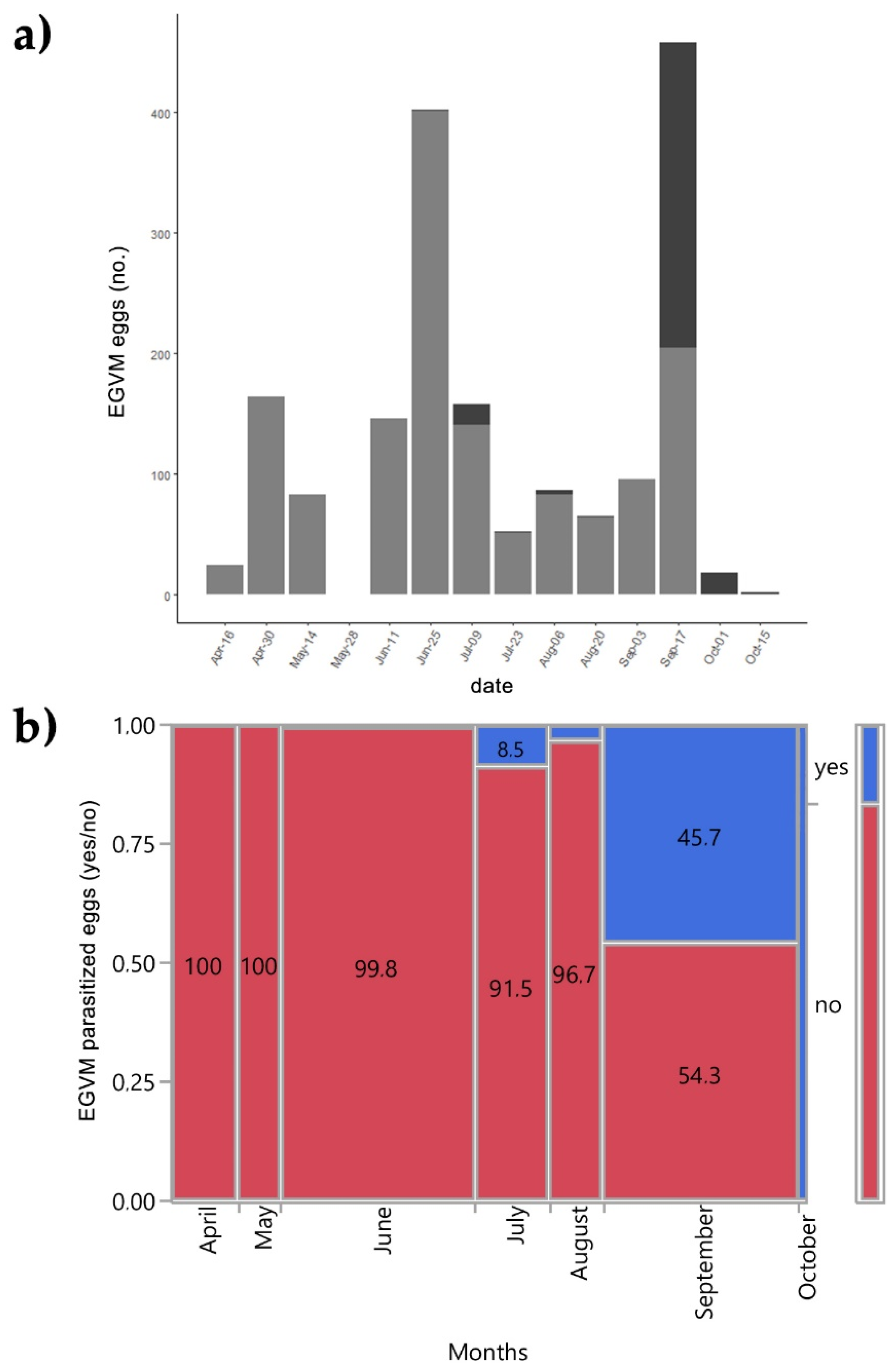
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global food security, biodiversity conservation and the future of agricultural intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Gavin, M.C.; McCarter, J.; Berkes, F.; Mead, A.T.P.; Sterling, E.J.; Tang, R.; Turner, N.J. Effective biodiversity conservation requires dynamic, pluralistic, partnership-based approaches. Sustainability 2018, 10, 1846. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Benelli, G. Towards pesticide-free farming? Sharing needs and knowledge promotes Integrated Pest Management. Environ. Sci. Pollut. Res. 2018, 25, 13439–13445. [Google Scholar] [CrossRef] [PubMed]
- Dent, D.; Binks, R.H. Insect Pest Management, 3rd ed.; CABI: Wallingford, UK, 2020; ISBN 978-1-78924-105-1. [Google Scholar]
- Silva, V.; Mol, H.G.J.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide residues in European agricultural soils—A hidden reality unfolded. Sci. Total Environ. 2019, 653, 1532–1545. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7. [Google Scholar] [CrossRef]
- Mancini, F.; Woodcock, B.A.; Isaac, N.J.B. Agrochemicals in the wild: Identifying links between pesticide use and declines of nontarget organisms. Curr. Opin. Environ. Sci. Health 2019, 11, 53–58. [Google Scholar] [CrossRef]
- Palma-Onetto, V.; Oliva, D.; González-Teuber, M. Lethal and oxidative stress side effects of organic and synthetic pesticides on the insect scale predator Rhyzobius lophanthae. Entomol. Gen. 2021, 345–355. [Google Scholar] [CrossRef]
- European Food Safety Authority; Medina-Pastor, P.; Triacchini, G. The 2018 European Union report on pesticide residues in food. EFSA J. 2020, 18, e06057. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.A.; Lehmler, H.-J.; Kolpin, D.W.; Hladik, M.L.; Vargo, J.D.; Schilling, K.E.; LeFevre, G.H.; Peeples, T.L.; Poch, M.C.; LaDuca, L.E.; et al. A critical review on the potential impacts of neonicotinoid insecticide use: Current knowledge of environmental fate, toxicity, and implications for human health. Environ. Sci. Process. Impacts 2020, 22, 1315–1346. [Google Scholar] [CrossRef] [PubMed]
- Corcos, D.; Inclán, D.J.; Cerretti, P.; Mei, M.; Di Giovanni, F.; Birtele, D.; Rosa, P.; De Biase, A.; Audisio, P.; Marini, L. Environmental heterogeneity effects on predator and parasitoid insects vary across spatial scales and seasons: A multi-taxon approach. Insect Conserv. Divers. 2017, 10, 462–471. [Google Scholar] [CrossRef]
- Rudd, H.; Vala, J.; Schaefer, V. Importance of backyard habitat in a comprehensive biodiversity conservation strategy: Aconnectivity analysis of urban green spaces. Restor. Ecol. 2002, 10, 368–375. [Google Scholar] [CrossRef]
- Tscharntke, T.; Rand, T.A.; Bianchi, F.J. The landscape context of trophic interactions: Insect spillover across the crop—Noncrop interface. Ann. Zool. Fenn. 2005, 42, 421–432. [Google Scholar]
- Weibull, A.-C.; Bengtsson, J.; Nohlgren, E. Diversity of butterflies in the agricultural landscape: The role of farming system and landscape heterogeneity. Ecography 2000, 23, 743–750. [Google Scholar] [CrossRef]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.-L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Miyashita, T.; Chishiki, Y.; Takagi, S.R. Landscape heterogeneity at multiple spatial scales enhances spider species richness in an agricultural landscape. Popul. Ecol. 2012, 54, 573–581. [Google Scholar] [CrossRef]
- Bertrand, C.; Burel, F.; Baudry, J. Spatial and temporal heterogeneity of the crop mosaic influences carabid beetles in agricultural landscapes. Landsc. Ecol. 2016, 31, 451–466. [Google Scholar] [CrossRef]
- Martin, E.A.; Seo, B.; Park, C.-R.; Reineking, B.; Steffan-Dewenter, I. Scale-dependent effects of landscape composition and configuration on natural enemy diversity, crop herbivory, and yields. Ecol. Appl. 2016, 26, 448–462. [Google Scholar] [CrossRef]
- Romano, B.; Zullo, F. Landscape fragmentation in Italy dices implementation to support territorial policies. In Planning Support Tools: Policy Analysis, Implementation and Evaluation; Campagna, M., De Montis, A., Isola, F., Lai, S., Pira, C., Zoppi, C., Eds.; Franco Angeli: Milano, Italy, 2012; pp. 399–414. [Google Scholar]
- Godone, D.; Garbarino, M.; Sibona, E.; Garnero, G.; Godone, F. Progressive fragmentation of a traditional Mediterranean landscape by hazelnut plantations: The impact of CAP over time in the Langhe region (NW Italy). Land Use Policy 2014, 36, 259–266. [Google Scholar] [CrossRef]
- Pili, S.; Serra, P.; Salvati, L. Landscape and the city: Agro-forest systems, land fragmentation and the ecological network in Rome, Italy. Urban. For. Urban. Green. 2019, 41, 230–237. [Google Scholar] [CrossRef]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Thiéry, D.; Louâpre, P.; Muneret, L.; Rusch, A.; Sentenac, G.; Vogelweith, F.; Iltis, C.; Moreau, J. Biological protection against grape berry moths. A review. Agron. Sustain. Dev. 2018, 38, 15. [Google Scholar] [CrossRef]
- Ioriatti, C.; Lucchi, A.; Varela, L.G. Grape berry moths in Western European vineyards and their recent movement into the New World. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 339–359. ISBN 978-94-007-4032-7. [Google Scholar]
- Cooper, M.; Varela, L.; Smith, R.; Whitmer, D.; Simmons, G.; Lucchi, A.; Broadway, R.; Steinhauer, R. Growers, scientists and regulators collaborate on European grapevine moth program. Cal. Ag. 2014, 68, 125–133. [Google Scholar] [CrossRef][Green Version]
- CABI Lobesia botrana (European Grapevine Moth). Available online: https://www.cabi.org/isc/datasheet/42794 (accessed on 1 March 2022).
- Marchal, P. Rapport sur les Travaux Accomplis par la Mission D’étude de la Cochylis et de l’Eudémis PENDANT l’année 1911; Ch. Béranger: Paris et Liège, France, 1912. [Google Scholar]
- Maher, N.; Thiéry, D. Daphne gnidium, a possible native host plant of the European grapevine moth Lobesia botrana, stimulates its oviposition. Is a host shift relevant? Chemoecology 2006, 16, 135–144. [Google Scholar] [CrossRef]
- Lucchi, A.; Santini, L. Life history of Lobesia botrana on Daphne gnidium in a Natural Park of Tuscany. IOBC/WPRS Bull. 2011, 67, 197–202. [Google Scholar]
- Tasin, M.; Lucchi, A.; Ioriatti, C.; Mraihi, M.; De Cristofaro, A.; Boger, Z.; Anfora, G. Oviposition response of the moth Lobesia botrana to sensory cues from a host plant. Chem. Senses 2011, 36, 633–639. [Google Scholar] [CrossRef]
- Laccone, G. Prove di lotta contro Lobesia botrana (Schiff.) (Lepid.—Tortricidae) e determinazione della soglia economica sulle uve da tavola in Puglia. Ann. Della Fac. Di Agraria. Dell’università Di Bari 1978, 30, 717–746. [Google Scholar]
- Moleas, T. Essais de lutte dirigée contre la Lobesia botrana Schiff. dans les Pouilles (Italie). In Proceedings of the International Symposium of IOBC/WPRS on Integrated Control in Agriculture and Forestry, Vienna, Austria, 8–12 October 1979; Russ, K., Berger, H., Eds.; 1979; pp. 542–551. [Google Scholar]
- Delrio, G.; Luciano, P.; Prota, R. Researches on grape-vine moths in Sardinia. In Proceedings of the Meeting of E. C. Experts Group “Integrated Pest Control in Viticulture”, Portoferraio, Italy, 26–28 September 1985; Cavalloro, R., Ed.; 1987; pp. 57–67. [Google Scholar]
- Zangheri, S.; Dalla Montà, L.; Duso, C. Observations on biology and control of grape moths in Venetia. In Proceedings of the Meeting of E. C. Expert Group “Integrated Pest Control in Viticulture”, Portoferraio, Italy, 26–28 September 1985; Cavalloro, R., Ed.; 1987; pp. 27–37. [Google Scholar]
- Pinna, M.; Gremo, F.; Scaramozzino, P.L. A preliminary investigation into the influence of biotic and abiotic environmental factors on the winter populations of Lobesia botrana (Den. & Schiff.) in veniyards in Piedmont, Italy (Lepidoptera—Tortricidae). In Proceedings of the Influence of Environmental Factors on the Control of Grape Pests, Diseases and Weeds, Thessaloniki, Greece, 6–8 October 1987; Cavalloro, R., Ed.; 1989; pp. 77–86. [Google Scholar]
- Forti, D. Resultats preliminaires sur l’activité des parasites du ver de la grappe (Lobesia botrana Schiff.) dans le Trentine. Bull. OILB/SROP 1991, XV, 9. [Google Scholar]
- Marchesini, E.; Dalla Montà, L. Observations on natural enemies of Lobesia botrana (Den. & Schiff.) (Lepidoptera Tortricidae) in Venetian vineyards. Boll. Zool. Agr. Bachic. 1994, 26, 201–230. [Google Scholar]
- Roat, C.; Forti, D. Indagine sulla parassitizzazione della tignoletta della vite (Lobesia botrana Den. & Schiff.) in Trentino. Boll. ISMA 1994, 1, 37–41. [Google Scholar]
- Pérez, M.I.; Sáenz de Cabezón, F.J.; Marco, V. Evaluation of natural parasitism on hibernating pupae of the European grape moth (Lobesia botrana Den. & Schiff.) in vineyard of La Rioja. Boletín De Sanid. Veg. Plagas 2000, 26, 715–721. [Google Scholar]
- Colombera, S.; Alma, A.; Arzone, A. Comparison between the parasitoids of Lobesia botrana and Eupoecilia ambiguella in conventional and integrated vineyards. IOBC/WPRS Bull. 2001, 24, 91–96. [Google Scholar]
- Ribeiro, J.J.A.; Martins, F.M.; Mendonça, T.R.; Lavadinho, A.M.P. Natural parasitism of Lobesia botrana during the hibernation period in the Region of “Vinhos Verdes”. IOBC/WPRS Bull. 2001, 24, 117–120. [Google Scholar]
- Thiéry, D.; Xuéreb, A.; Villemant, C.; Sentenac, G.; Delbac, L.; Kuntzman, P. Les parasites larvaires de tordeuses de vignobles: Aperçu de quelques espèces présentes dans 3 régions viticoles françaises. IOBC/WPRS Bull. 2001, 24, 135–142. [Google Scholar]
- Thiéry, D.; Xuéreb, A. Relative abundance of several larval parasitoids of Lobesia botrana on different varieties of grapes. IOBC/WPRS Bull. 2003, 26, 147–150. [Google Scholar]
- Bagnoli, B.; Lucchi, A. Parasitoids of Lobesia botrana (Den. & Schiff.) in Tuscany. IOBC/WPRS Bulletin 2006, 29, 139–142. [Google Scholar]
- Carlos, C.R.; Costa, J.R.; Tão, C.B.; Alves, F.; Torres, L.M. Parasitismo associado à traça-da-uva, Lobesia botrana (Denis & Schiffermüller) na Região Demarcada do Douro. Bol. San. Veg. Plagas 2006, 32, 355–362. [Google Scholar]
- Thiéry, D.; Yoshida, T.; Guisset, M. Phytomyptera nigrina (Meigen), a parasite of first generation European grapevine moth larvae in several vineyards in the Roussillon area. Tachinid Times 2006, 19, 1–4. [Google Scholar]
- Xuéreb, A.; Thiéry, D. Does natural larval parasitism of Lobesia botrana (Lepidoptera: Tortricidae) vary between years, generation, density of the host and vine cultivar? Bull. Entomol. Res. 2006, 96, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Bărbuceanu, D.; Jenser, G. The parasitoid complex of Lobesia botrana (Denis et Schiffermüller) (Lep.: Tortricidae) in some vineyards of southern Romania. Acta Phytopathol. Et Entomol. Hung. 2009, 44, 177–184. [Google Scholar] [CrossRef]
- Moreau, J.; Villemant, C.; Benrey, B.; Thiéry, D. Species diversity of larval parasitoids of the European grapevine moth (Lobesia botrana, Lepidoptera: Tortricidae): The influence of region and cultivar. Biol. Control. 2010, 54, 300–306. [Google Scholar] [CrossRef]
- Akbarzadeh, S.G. Larval parasitoids of Lobesia botrana (Denis and Schiffermüller, 1775) (Lepidoptera: Tortricidae) in Orumieh vineyards. J. Agr. Sci. Tech. 2012, 14, 267–274. [Google Scholar]
- Lotfalizadeh, H.; Masnadi-Yazdinejad, A.; Saber, M. New records of the grape berry moth hymenopterous parasitoids in Iran. Mun. Ent. Zool. 2012, 7, 284–291. [Google Scholar]
- Carlos, C.; Gonçalves, F.; Sousa, S.; Salvação, J.; Sharma, L.; Soares, R.; Manso, J.; Nóbrega, M.; Lopes, Á.; Soares, S.; et al. Environmentally safe strategies to control the European Grapevine Moth, Lobesia botrana (Den. & Schiff.), in the Douro Demarcated Region. Ciência E Técnica Vitivinícola 2013, 28, 1006–1011. [Google Scholar]
- Ozsemerci, F.; Altindisli, F.O.; Koclu, T.; Karsavuran, Y. Egg parasitoids of Lobesia botrana (Den. & Schiff.) (Lepidoptera: Tortricidae) in the vineyards of Izmir and Manisa Provinces in Turkey. BIO Web Conf. 2016, 7, 1–4. [Google Scholar] [CrossRef]
- Shapira, I.; Keasar, T.; Harari, A.R.; Gavish-Regev, E.; Kishinevsky, M.; Steinitz, H.; Sofer-Arad, C.; Tomer, M. Does mating disruption of Planococcus ficus and Lobesia botrana affect the diversity, abundance and composition of natural enemies in Israeli vineyards? Pest. Manag. Sci. 2018, 74, 1837–1844. [Google Scholar] [CrossRef]
- Carlos, C.; Gonçalves, F.; Villemant, C.; Paredes, D.; Salvação, J.; Torres, L. Parasitoids of Lobesia botrana (Lepidoptera: Tortricidae) in Douro Demarcated Region vineyards and prospects for enhancing conservation biological control. Bull. Entomol. Res. 2022, 1–10. [Google Scholar] [CrossRef]
- Scaramozzino, P.L.; Di Giovanni, F.; Loni, A.; Gisondi, S.; Lucchi, A.; Cerretti, P. Tachinid (Diptera, Tachinidae) parasitoids of Lobesia botrana (Denis & Schiffermüller, 1775) (Lepidoptera, Tortricidae) and other moths. ZooKeys 2020, 934, 111–140. [Google Scholar] [CrossRef]
- Nuzzaci, G.; Triggiani, O. Note sulla biocenosi in Puglia della Lobesia (Polychrosis) botrana (Schifi.) (Lepidoptera: Tortricidae) infeudata a Daphne gnidium L. Entomologica 1982, 17, 47–52. [Google Scholar] [CrossRef]
- Luciano, P.; Delrio, G.; Prota, R. Osservazioni sulle popolazioni di Lobesia botrana (Den. & Schiff.) su Daphne gnidium L. in Sardegna. Atti XV Congr. Naz. Ital. Di Entomol. 1988, 15, 543–548. [Google Scholar]
- Loni, A.; Samartsev, K.G.; Scaramozzino, P.L.; Belokobylskij, S.A.; Lucchi, A. Braconinae parasitoids (Hymenoptera, Braconidae) emerged from larvae of Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Tortricidae) feeding on Daphne gnidium L. ZooKeys 2016, 587, 125–150. [Google Scholar] [CrossRef]
- Lucchi, A.; Scaramozzino, P.L.; Michl, G.; Loni, A.; Hoffmann, C. The first record in Italy of Trichogramma cordubense Vargas & Cabello 1985 (Hymenoptera Trichogrammatidae) emerging from the eggs of Lobesia botrana (Denis & Schiffermüller, 1775) (Lepidoptera Tortricidae). VITIS—J. Grapevine Res. 2016, 55, 161–164. [Google Scholar] [CrossRef]
- Scaramozzino, P.L.; Loni, A.; Gandini, L.; Lucchi, A. Who’s who in the nests molded by Lobesia botrana on Daphne gnidium? IOBC/WPRS Bull. 2017, 128, 130–134. [Google Scholar]
- Federici, P.R. Atlante Tematico Della Provincia di Pisa; Pacini Editore: Pisa, Italy, 2003; p. 80. [Google Scholar]
- Bertacchi, A. Dune habitats of the Migliarino—San Rossore—Massaciuccoli Regional Park (Tuscany—Italy). J. Maps 2017, 13, 322–331. [Google Scholar] [CrossRef]
- Bertacchi, A.; Lombardi, T. Aspetti botanici delle pinete litoranee toscane. In Le Pinete Litoranee Come Patrimonio Culturale; Atti della Accademia dei Georgofili. Suppl.; Edizioni Polistampa: Firenze, Italy, 2019; pp. 59–68. [Google Scholar]
- Logli, F. Le pinete del Parco Migliarino San Rossore Massaciuccoli. In Le Pinete Litoranee Come Patrimonio Culturale; I Atti della Accademia dei Georgofili. Suppl.; Edizioni Polistampa: Firenze, Italy, 2019; pp. 43–52. [Google Scholar]
- Scaramozzino, P.L.; Loni, A.; Lucchi, A. A review of insect parasitoids associated with Lobesia botrana (Denis & Schiffermüller, 1775) in Italy. 1. Diptera Tachinidae and Hymenoptera Braconidae (Lepidoptera, Tortricidae). ZooKeys 2017, 647, 67–100. [Google Scholar] [CrossRef]
- Scaramozzino, P.L.; Di Giovanni, F.; Loni, A.; Ricciardi, R.; Lucchi, A. Updated list of the insect parasitoids (Insecta, Hymenoptera) associated with Lobesia botrana (Denis & Schiffermüller, 1775) (Lepidoptera, Tortricidae) in Italy. 2. Hymenoptera, Ichneumonidae, Anomaloninae and Campopleginae. ZooKeys 2018, 772, 47–95. [Google Scholar] [CrossRef]
- Huddleston, T. The Palaearctic species of Ascogaster (Hymenoptera: Braconidae). Bull. Br. Mus. Nat. Hist., Ent. 1984, 49, 341–392. [Google Scholar]
- Peck, O.; Bouček, Z.; Hoffer, A. Keys to the Chalcidoidea of Czechoslovakia (Insecta: Hymenoptera). Mem. Ent. Soc. Can. 1964, S34, 7–121. [Google Scholar] [CrossRef]
- Cerretti, P. I Tachinidi Della Fauna Italiana (Diptera, Tachinidae), con Chiave Interattiva dei Generi Ovest-Paleartici; Centro Nazionale Biodiversità Forestale, Cierre Edizioni: Verona, Italy, 2010; p. 912. [Google Scholar]
- Aeschlimann, J.P. Revision des espèces ouest-palearctiques du genre Triclistus Förster (Hymenoptera: Ichneumonidae). Mitt. Schweiz. Entomol. Ges. 1973, 46, 219–252. [Google Scholar]
- Kasparyan, D.R. A review of the Palearctic ichneumonids of the tribe Pimplini (Hymenoptera, Ichneumonidae). The genera Itoplectis Foerst. and Apechthis Foerst. Entomol. Obozr. 1973, 52, 665–681. [Google Scholar]
- Kasparyan, D.R. A review of the Palearctic ichneumonids of the tribe Pimplini (Hymenoptera, Ichneumonidae). The genus Pimpla Fabricius. Entomol. Obozr. 1974, 53, 102–117. [Google Scholar]
- Gauld, I.D.; Mitchell, P.A. Handbooks for the Identification of British Insects; Vol. VII. Part 2.; Ichneumonidae Orthopelmatinae & Anomaloninae, Royal Entomological Society of London: London, UK, 1977; p. 32. [Google Scholar]
- Horstmann, K. Revision der mit difformis (Gmelin, 1790) verwandten westpaläarktischen Arten der Gattung Campoplex Gravenhorst, 1829 (Hymenoptera, Ichneumonidae). Entomofauna 1985, 6, 129–163. [Google Scholar]
- Tolkanitz, V.I. Ichneumon flies of the genus Exochus Gravenhorst (Hymenoptera: Ichneumonidae: Metopiinae) of the fauna of Palearctic region. Russ. Entomol. J. 2007, 16, 339–358. [Google Scholar]
- Sumer, F.; Tuncbilek, A.S.; Oztemiz, S.; Pintureau, B.; Rugman-Jones, P.F.; Stouthamer, R. A molecular key to the common species of Trichogramma of the Mediterranean region. BioControl 2009, 54, 617–624. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Elsevier/Academic Press: Burlington, NJ, USA, 2006; p. 324. [Google Scholar]
- Gaston, J.K.; He, F. Species occurrence and occupancy. In Biological Diversity: Frontiers in Measurement and Assessment; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 141–151. [Google Scholar]
- Mills, N.J. Parasitoid guilds: Defining the structure of the parasitoid communities ofendopterygote insect hosts. Environ. Entomol. 1994, 23, 1066–1083. [Google Scholar] [CrossRef]
- Thompson, W.R. A Catalogue of the Parasites and Predators of Insect Pest; Section 1. Parasite Host Catalogue, Part. 8. Parasites of the Lepidoptera (N-P); Commonwealth Institute of Biological Control, Imperial Agricultural Bureaux, Parasite Service: Belleville, ON, Canada, 1946. [Google Scholar]
- Coscollá, R. La polilla del racimo de la vid (Lobesia botrana Den. y Schiff.). In Consejerfa de Agricultura, Pesca y Alimentación; Generalitat Valenciana: Valencia, Spain, 1997; p. 613. [Google Scholar]
- Hoffmann, C.; Michl, G. Parasitoide von Traubenwicklern—Ein Werkzeug der naturlichen Schadlingsregulation? Dtsch. Weinbaujahrbuch 2003, 55, 1–13. [Google Scholar]
- Yu, D.S.; van Achterberg, C.; Horstmann, K. Taxapad 2016. World Ichneumonoidea (2015). Taxonomy, Biology, Morphology and Distribution; Nepean: Ottawa, ON, Canada, 2016. [Google Scholar]
- Di Giovanni, F.; Scaramozzino, P.L.; Loni, A.; Lucchi, A. Taxonomic revision of the Campoplex difformis group (Ichneumonidae, Campopleginae), with particular reference to species of economic importance. Eur. J. Taxon. 2021, 740, 1–35. [Google Scholar] [CrossRef]
- Villemant, C.; Delvare, G.; Martinez, M.; Sentenac, G.; Kuntzman, P. Parasitoïdes de tordeuses. In La Faune Auxiliaire des Vignobles de France; Sentenac, G., Ed.; Editions France Agricole: Paris, France, 2011; p. 470. [Google Scholar]
- Gordh, G. Goniozus gallicola Fouts, parasite of moth larvae with notes on other bethylids (Hymenoptera: Bethylidae; Lepidoptera: Gelechiidae). USDA Tech. Bull. 1976, 1524, 1–27. [Google Scholar]
- Mellini, E. Studi sui ditteri larvevoridi. XII. Nemorilla maculosa Meig. su Depressaria marcella Rebel (Lepidoptera Gelechiidae). Boll. Dell’istituto Di Entomol. Dell’università Di Bologna 1964, 27, 145–169. [Google Scholar]
- Rohdendorf, B.B. Beiträge zur Kenntnis der Tachinen des Wiesenzünslers (Loxostege sticticalis L.). Izv. Akad. Nauk SSSR 1935, 5, 753–780. [Google Scholar]
- Catoni, G. Die Traubenwikler (Polychrosis botrana Schiff. und Conchylis ambiguella Hübn.) und ihre natürlichen Feinde in Südtyrol. Z. Für Angew. Entomol. 1914, 1, 248–259. [Google Scholar] [CrossRef]
- Schwangart, F. Ueber die Traubenwickler (Clysia [Conchylis] Ambiguella Hübn. und Polychrosis Botrana Schiff.) und ihre Bekämpfung, mit Berücksichtigung Natürlicher Bekämpfungsfaktoren. Zweiter Teil; Verlag von Oustav Fischer: Jena, Germany, 1913. [Google Scholar]
- Bordera, S.; Selfa, J. Datos sobre la fauna ibérica del género Bathythrix Foerster, 1868 (Hym., Ichneumonidae). Bolétin De La Asoc. Española De Entomol. 1992, 16, 193–197. [Google Scholar]
- Sawoniewicz, J. Revision of European species of the genus Bathythrix Förster (Hym., Ichneumonidae). Ann. Zool. 1980, 35, 319–365. [Google Scholar]
- Kasparyan, D.R.; Kopelke, J.P. Taxonomic review and key to European ichneumon flies (Hymenoptera, Ichneumonidae), parasitoids of gall-forming sawflies of the genera Pontania Costa, Phyllocolpa Benson, and Euura Newman (Hymenoptera, Tenthredinidae) on willows: Part I. Entomol. Rev. 2009, 89, 933–957. [Google Scholar] [CrossRef]
- Noyes, J.S. Universal Chalcidoidea Database. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 8 June 2022).
- Coscollá, R. Aproximación al estudio del parasitismo natural sobre Lobesia botrana Den. y Schiff. en las comarcas vitícolas Valencianas. Boletín De Sanid. Veg.—Plagas 1980, 6, 5–15. [Google Scholar]
- Genini, M. Antagonistes de la cicadelle verte et des vers de la grappe dans le vignoble valaisan et les milieux environnants. Rev. Suisse De Vitic. Arboric. Et Hortic. 2000, 32, 153–162. [Google Scholar]
- Lucchi, A.; Ricciardi, R.; Loni, A.; Cosci, F.; Alvarez, A.R.; Beeche, M.; Scaramozzino, P.L. Rearing Campoplex capitator Aubert in Italy and in Chile: Preliminary achievements. In Proceedings of the Future IPM 3.0 towards a Sustainable Agriculture, IOBC/WPRS General Assembly, Riva del Garda, Italy, 15–20 October 2017; p. 370. [Google Scholar]
- Ibrahim, R.A.E.M.A. Biological control of grape berry moths Eupoecilia ambiguella Hb. and Lobesia botrana Schiff. (Lepidoptera: Tortricidae) by using egg parasitoids of the genus Trichogramma. Ph.D. Thesis, Institute of Phytopathology and Applied Zoology, Justus Liebig University of Giessen, Giessen, Germany, 2004. [Google Scholar]
- Hommay, G.; Gertz, C.; Kienlen, J.C.; Pizzol, J.; Chavigny, P. Comparison between the control efficacy of Trichogramma evanescens Westwood (Hymenoptera: Trichogrammatidae) and two Trichogramma cacoeciae Marchal strains against grapevine moth (Lobesia botrana Den. & Schiff.), depending on their release density. Biocontrol. Sci. Technol. 2002, 12, 569–581. [Google Scholar] [CrossRef]
- Naeem, S. Species redundancy and ecosystem reliability. Conserv. Biol. 1998, 12, 7. [Google Scholar] [CrossRef]
- Mori, A.S.; Furukawa, T.; Sasaki, T. Response diversity determines the resilience of ecosystems to environmental change. Biol. Rev. 2013, 88, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Flynn, D.F.; Prager, C.M.; Hart, G.M.; DeVan, C.M.; Ahrestani, F.S.; Palmer, M.I.; Bunker, D.E.; Knops, J.M.; Jouseau, C.F.; et al. The importance of rare species: A trait-based assessment of rare species contributions to functional diversity and possible ecosystem function in tall-grass prairies. Ecol. Evol 2014, 4, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, A.; Loni, A.; Scaramozzino, P.L. Pests in the wild: Life history and behaviour of Lobesia botrana on Daphne gnidium in a natural environment (Lepidoptera Tortricidae; Malvales Thymelaeaceae). IOBC/WPRS Bull. Meet. Integr. Prot. Prod. Vitic. 2017, 128, 84–88. [Google Scholar]
- Moreau, J.; Buffenoir, N.; Thiéry, D.; Vogelweith, F. Lobesia botrana as a preferred host of Campoplex capitator, the most occurring larval parasitoid in European vineyards. Entomol. Gen. 2019, 39, 307–312. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
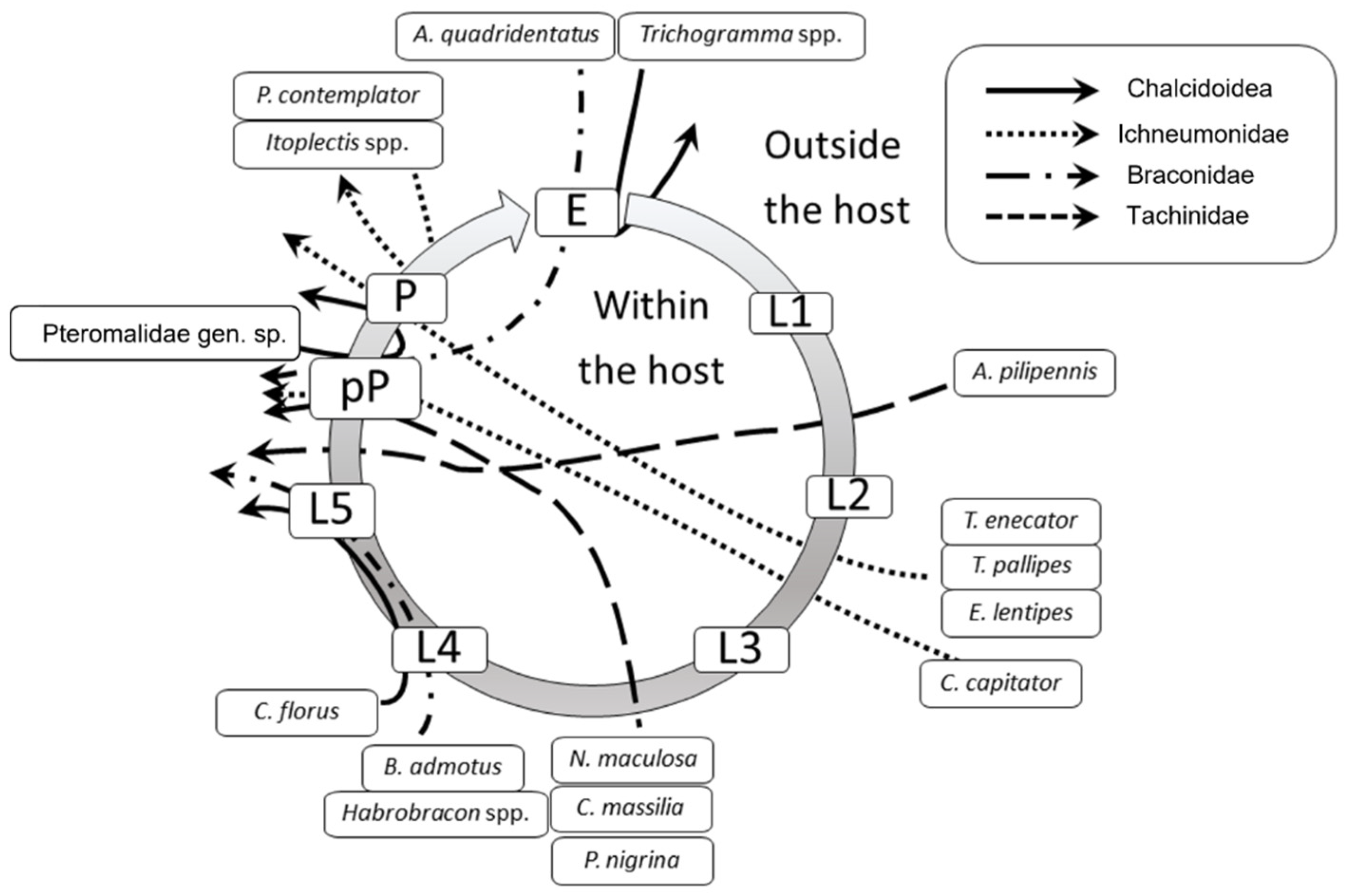
| Family and Species | 2014 | 2015 | 2017 | TOT |
|---|---|---|---|---|
| Braconidae | 30 (2.05) | 2 (0.16) | 5 (0.32) | 37 (0.87) |
| Ascogaster quadridentata Wesmael, 1835 | - | 1 | - | 1 |
| Bracon admotus Papp, 2000 | 2 | 1 | 1 | 4 |
| Habrobracon concolorans (Marshall, 1900) | 9 | - | - | 9 |
| Habrobracon hebetor (Say, 1836) | 3 | - | - | 3 |
| Habrobracon pillerianae Fisher, 1980 | 16 | - | 4 | 20 |
| Eulophidae | 12 (0.82) | 12 (0.97) | 2 (0.13) | 26 (0.61) |
| Colpoclypeus florus (Walker, 1839) | 12 * | 12 | 2 | 26 |
| Eupelmidae | 1 (0.07) | - | - | 1 (0.02) |
| Eupelmus sp. | 1 | - | - | 1 |
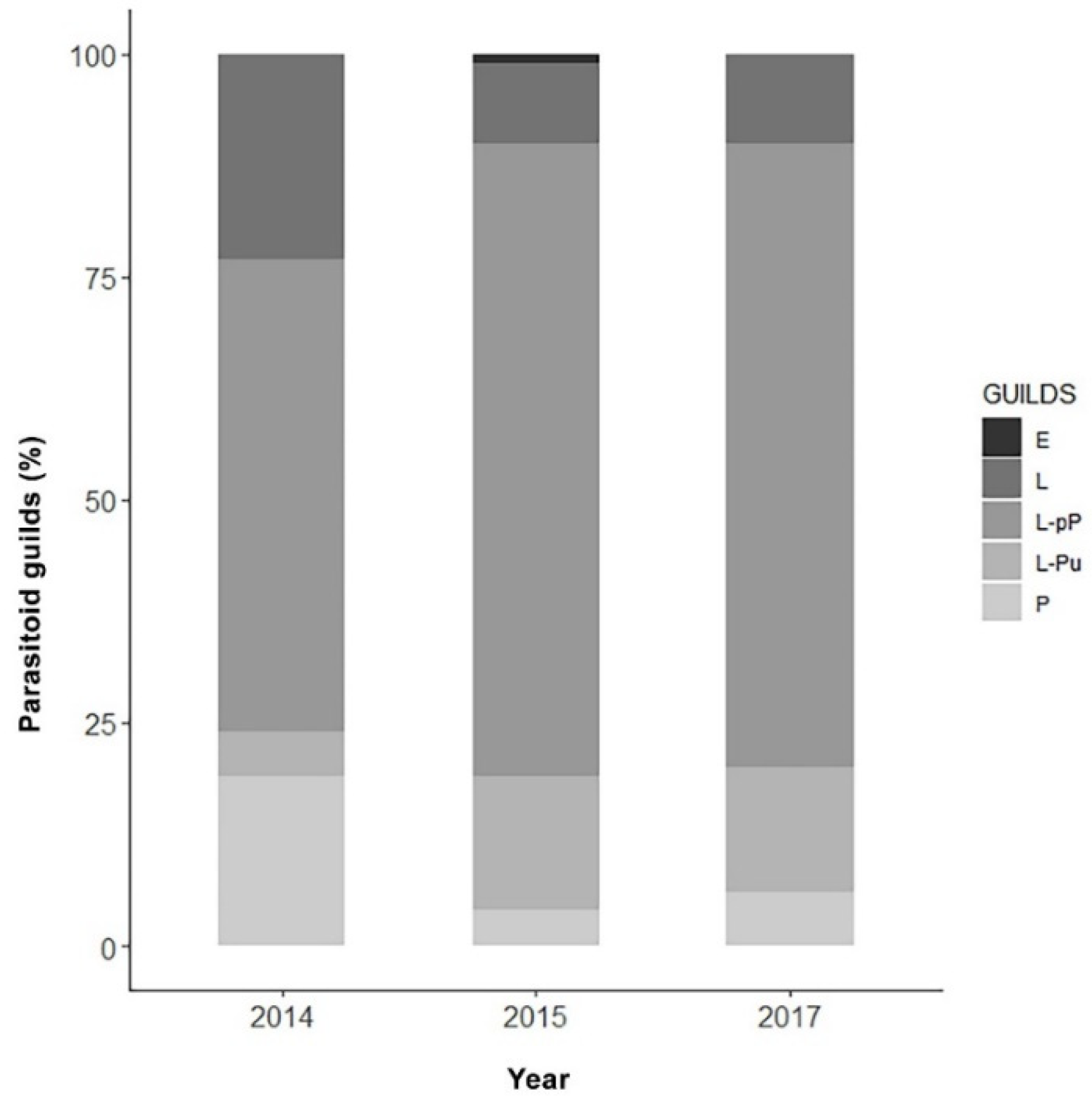
| Ichneumonidae | 181 (12.38) | 143 (11.61) | 145 (9.16) | 469 (10.97) |
| Campoplex capitator Aubert, 1960 | 128 (8.76) | 114 (9.25) | 112 (7.08) | 354 (8.28) |
| Exochus lentipes Gravenhorst, 1829 | 1 | 3 | - | 4 |
| Itoplectis alternans (Gravenhorst, 1829) | 13 ** | 1 | 7 | 21 |
| Itoplectis maculator (Fabricius, 1775) | 3 | - | - | 3 |
| Itoplectis tunetana (Schmiedeknecht, 1914) | 25 | 4 | 3 | 32 |
| Pimpla contemplator (Müller, 1776) | 1 | - | - | 1 |
| Trichomma enecator (Rossi, 1790) | 7 | - | - | 7 |
| Triclistus pallipes Holmgren, 1873 | 3 | 21 | 23 | 47 |
| Pteromalidae | 3 (0.20) | 1 (0.08) | - | 4 (0.09) |
| gen. sp. | 3 | 1 | - | 4 |
| Tachinidae | 12 (0.82) | 4 (0.32) | 11 (0.69) | 27 (0.63) |
| Actia pilipennis (Fallén, 1810) | 11 | 2 | 10 | 23 |
| Phytomyptera nigrina (Meigen, 1824) | 1 | - | - | 1 |
| Clemelis massilia Herting, 1977 | - | 1 | - | 1 |
| Nemorilla maculosa (Meigen, 1824) | - | 1 | 1 | 2 |
| Total | 239 (16.35) | 162 (13.15) | 163 (10.30) | 564 (13.19) |
| Lobesia botrana (Denis and Schiffermüller, 1775) | 1223 | 1070 | 1420 | 3713 |
| Parasitoid Species | Occupancy (%) | ||
|---|---|---|---|
| 2014 | 2015 | 2017 | |
| Actia pilipennis (Fallén, 1810) | 55 | - | 67 |
| Campoplex capitator Aubert, 1960 | 100 | 100 | 100 |
| Colpoclypeus florus (Walker, 1839) | 55 | 44 | - |
| Habrobracon pillerianae Fischer, 1980 | - | - | 44 |
| Itoplectis alternans (Gravenhorst, 1829) | 44 | - | 67 |
| Itoplectis tunetana (Schmiedeknecht, 1914) | 88 | - | - |
| Trichomma enecator (Rossi, 1790) | 44 | - | - |
| Triclistus pallipes Holmgren, 1873 | 44 | 67 | 88 |
| Tuscany | Apulia | Sardinia | ||||||
|---|---|---|---|---|---|---|---|---|
| Host Plant | Lifestyle | V.v. [47,59] | D.g. [32,59,62,63,64] | V.v. [34,35,59] | D.g. [60] | V.v. [36] | D.g. [61] | |
Parasitoid Guild/Species [Ordo-Family] | ||||||||
| Egg endoparasitoid [EN] | ||||||||
| Trichogramma cacaeciae Marchal, 1927 [Hym-Trich] | I | ■ | ||||||
| Trichogramma cordubense Vargas and Cabello, 1985 [Hym-Trich] | I | ■ | ||||||
| Trichogramma evanescens Westwood, 1833 [Hym-Trich] | I | ■ | ■ | |||||
| Trichogramma euproctidis (Girault, 1911) [Hym-Trich] | I | ■ | ||||||
| Egg-prepupal endoparasitoid [EpPN] | ||||||||
| Ascogaster quadridentata Wesmael, 1835 [Hym-Brac] | K | ■ | ■ | |||||
| Chelonus sp. [Hym-Brac] | K | ■ | ||||||
| Early larval endoparasitoid [ELN] | ||||||||
| Phytomyptera nigrina (Meigen, 1824) [Dipt-Tach] | K | ■ | ■ | ■ | ■ | ■ | ||
| Late larval endoparasitoid [LLN] | ||||||||
| Actia pilipennis (Fallén, 1810) [Dipt-Tach] | K | ■ | ||||||
| Neoplectops pomonellae (Schnabl and Mokrzecki, 1903) [Dipt-Tach] | K | ■ | ||||||
| Larval ectoparasitoid [LC] | ||||||||
| Bracon admotus Papp, 2000 [Hym-Brac] | I G? | ■ | ||||||
| Colpoclypeus florus (Walker, 1839) [Hym-Eul] | KG | ■ | ■ | |||||
| Elachertus affinis Masi, 1911 [Hym-Eul] | I G | ■ | ■ | |||||
| Goniozus gallicola (Kieffer, 1905) [Hym-Bet] | I [G (1)] | ■ | ||||||
| Larval ectoparasitoid [LC] | ||||||||
| Habrobracon sp. [Hym-Brac] | I G | ■ | ||||||
| Habrobracon concolorans (Marshall, 1900) [Hym-Brac] | I G | ■ | ||||||
| Habrobracon hebetor (Say, 1936) [Hym-Brac] | I G | ■ | ■ | |||||
| Habrobracon pillerianae Fischer, 1980 [Hym-Brac] * | I G | ■ | ||||||
| Scambus elegans (Woldstedt, 1877) [Hym-Ichn] | I FH | ■ | ||||||
| Larval-prepupal endoparasitoid [LpPN] [a] Early attackers | ||||||||
| Agathis sp. [Hym-Brac] | K | ■ | ||||||
| Agathis malvacearum Latreille, 1805 [Hym-Brac] | K | ■ | ||||||
| Aleiodes sp. [Hym-Brac] | K | ■ | ||||||
| Apanteles sp. [Hym-Brac] | ■ | |||||||
| Apanteles halidayi Marshall, 1872 [Hym-Brac] | K | ■ | ||||||
| Campoplex borealis (Zetterstedt, 1838) [Hym-Ichn] | K | ■ | ||||||
| Campoplex capitator Aubert, 1960 [Hym-Ichn] | K | ■ | ■ | |||||
| Campoplex difformis (Gmelin, 1790) [Hym-Ichn] | K | ■ | ||||||
| Pristomerus vulnerator (Panzer, 1799) [Hym-Ichn] | K | ■ | ||||||
| Therophilus linguarius (Nees, 1812) [Hym-Brac] | K | ■ | ||||||
| Therophilus tumidulus (Nees, 1812) [Hym-Brac] | K | ■ | ||||||
| Larval-prepupal endoparasitoid [LpPN] [b] Late attackers | ||||||||
| Nemorilla maculosa (2) (Meigen, 1824) [Dipt-Tach] | K | ■ | ||||||
| Clemelis massilia (3) Herting, 1977 [Dipt-Tach] | K | ■ | ||||||
| Larval-prepupal ectoparasitoid [LpPC] | ||||||||
| Phytodietus sp. [Hym-Ichn] | K | ■ | ||||||
| Phytodietus polyzonias (Förster, 1771) [Hym-Ichn] | K | ■ | ||||||
| Larval-pupal endoparasitoid [LPN] | ||||||||
| Exochus sp. [Hym-Ichn] | K | ■ | ||||||
| Exochus lentipes Gravenhorst, 1829 [Hym-Ichn] | K | ■ | ||||||
| Trichomma enecator (Rossi, 1790) [Hym-Ichn] | K | ■ | ||||||
| Triclistus sp. [Hym-Ichn] | K | ■ | ■ | |||||
| Triclistus lativentris Thomson, 1887 [Hym-Ichn] | K | ■ | ||||||
| Triclistus pallipes Holmgren, 1873 [Hym-Ichn] | K | ■ | ||||||
| Prepupal-pupal ectoparasitoid [pPPC] | ||||||||
| Agrothereutes pumilus (Kriechbaumer, 1899) [Hym-Ichn] | I FH | ■ | ||||||
| Dibrachys affinis Masi, 1907 [Hym-Pterom] | I FH | ■ | ■ | ■ | ||||
| Dibrachys microgastri (Bouche, 1834) [Hym-Pterom] | I FH | ■ | ||||||
| Ischnus alternator (Gravenhorst, 1829) [Hym-Ichn] | I | ■ | ||||||
| Pupal endoparasitoid [PN] | ||||||||
| Dicaelotus inflexus Thomson, 1891 [Hym-Ichn] | I | ■ | ■ | |||||
| Hockeria sp. [Hym-Chalc] | I | ■ | ||||||
| Itoplectis alternans (Gravenhorst, 1829) [Hym-Ichn] | I FH | ■ | ■ | |||||
| Itoplectis maculator (Fabricius, 1775) [Hym-Ichn] | I FH | ■ | ||||||
| Itoplectis tunetana (Schmiedeknecht, 1914) [Hym-Ichn] | I FH | ■ | ||||||
| Pimpla apricaria Costa, 1885 [Hym-Ichn] | I | ■ | ||||||
| Pimpla spuria Gravenhorst, 1829 [Hym-Ichn] | I | ■ | ||||||
| Pimpla turionellae (Linnaeus, 1758) [Hym-Ichn] | I FH | ■ | ||||||
| Hyperparasitoid | ||||||||
| Bathythrix argentata (4) (Gravenhorst, 1829) [Hym-Ichn] | H | ■ | ||||||
| Theroscopus hemipteron (5) (Riche, 1791) [Hym-Ichn] | FH | ■ | ■ | |||||
| Uncertain placement | ||||||||
| Pachyneuron sp. [Hym-Pterom] | H | ■ | ||||||
| Tetrastichus sp. (6) [Hym-Eul] | ? | ■ | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Giovanni, F.; Ricciardi, R.; Loni, A.; Scaramozzino, P.L.; Benelli, G.; Lucchi, A. Back to the Wild: The Parasitoid Community of Lobesia botrana (Lepidoptera: Tortricidae) in a Grapevine-Free Natural Environment. Insects 2022, 13, 627. https://doi.org/10.3390/insects13070627
Di Giovanni F, Ricciardi R, Loni A, Scaramozzino PL, Benelli G, Lucchi A. Back to the Wild: The Parasitoid Community of Lobesia botrana (Lepidoptera: Tortricidae) in a Grapevine-Free Natural Environment. Insects. 2022; 13(7):627. https://doi.org/10.3390/insects13070627
Chicago/Turabian StyleDi Giovanni, Filippo, Renato Ricciardi, Augusto Loni, Pier Luigi Scaramozzino, Giovanni Benelli, and Andrea Lucchi. 2022. "Back to the Wild: The Parasitoid Community of Lobesia botrana (Lepidoptera: Tortricidae) in a Grapevine-Free Natural Environment" Insects 13, no. 7: 627. https://doi.org/10.3390/insects13070627
APA StyleDi Giovanni, F., Ricciardi, R., Loni, A., Scaramozzino, P. L., Benelli, G., & Lucchi, A. (2022). Back to the Wild: The Parasitoid Community of Lobesia botrana (Lepidoptera: Tortricidae) in a Grapevine-Free Natural Environment. Insects, 13(7), 627. https://doi.org/10.3390/insects13070627