
The Role of Insect Symbiotic Bacteria in Metabolizing Phytochemicals and Agrochemicals

Abstract
:Simple Summary
Abstract
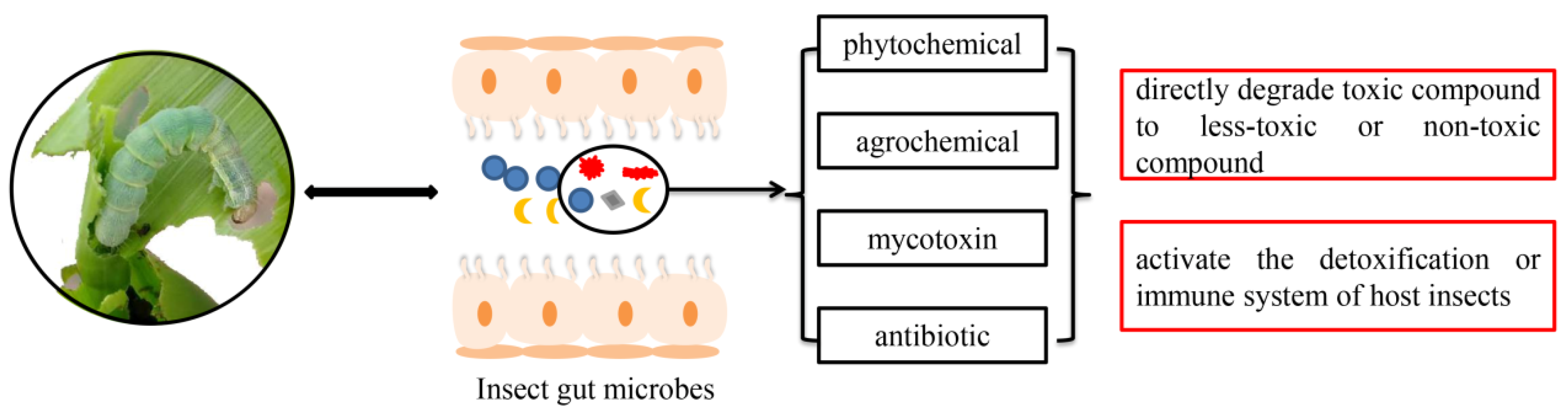
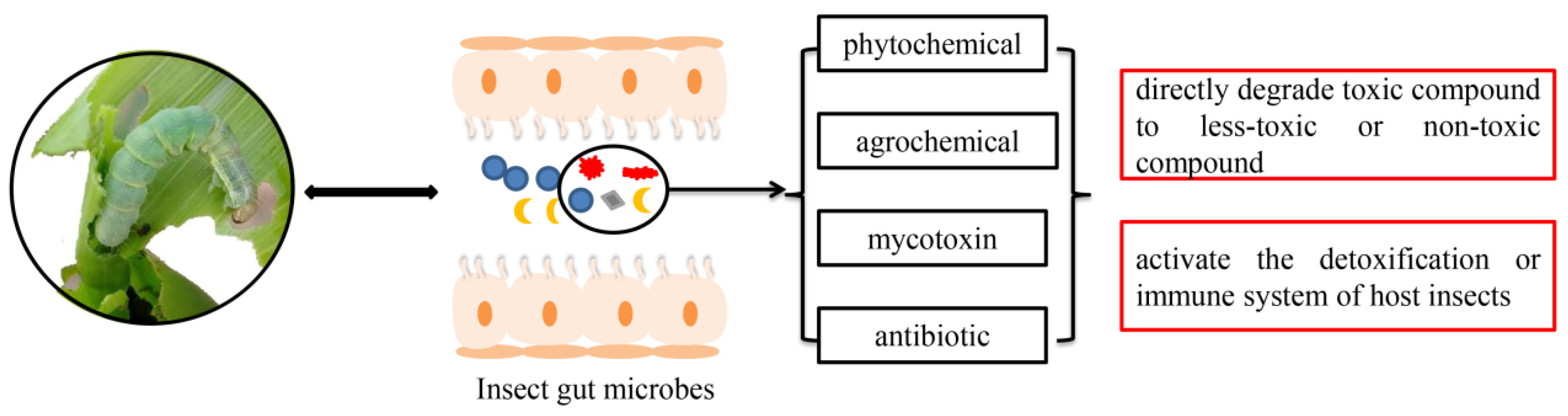
1. Introduction
2. Insect Bacteria Confer Resistance to Phytochemicals
3. Association between Gut Bacteria and Insects’ Adaptation to Agrochemicals
3.1. Symbionts Directly Degrade Agrochemicals
3.2. Indirect Regulation of Insect Resistance by Gut Bacteria

{kind=link}
| Bacteria and Insect Host | Target Agrochemical | Description | Reference |
|---|---|---|---|
| Serratia oryzae and Acinetobacter junii in Aedes albopictus | Deltamethrin | S. oryzae and A. junii had higher abundance in deltamethrin-resistant strain (by 16S rRNA sequencing) | [72] |
| Arthrobacter nicotinovorans in Spodoptera frugiperda | Cleared 54.9% of deltamethrin (by LC-MS) | [75] | |
| Burkholderia strains in Riptortus pedestris and Cletus punctiger | Fenitrothion | Bacteria metabolized fenitrothion into nontoxic substance, and insects infected with fenitrothion-degrading Burkholderia strains had higher survival rate and larger body size (by HPLC). | [76,77] |
| Pseudomonas aeruginosa G1, Stenotrophomonas maltophilia G2, and Acinetobacter lwoffii G5 in Blatta orientalis | α-endosulfan | Degradation rates of P. aeruginosa G1, S. maltophilia G2, and A. lwoffii G5 to α-endosulfan were 88.5%, 85.5%, and 80.2%, respectively (by HPLC) | [78] |
| Sphingobacterium, and Lysinibacillus Streptococcus and Rubrobacter in Anopheles gambiae | Pyrethroid | Sphingobacterium, Lysinibacillus, Streptococcus, and Rubrobacter significantly more abundant in resistant mosquitoes (by 16S rRNA gene sequencing) | [79] |
| Acetobacter tropicalis in Drosophila melanogaster | Atrazine | Atrazine exposure reduced relative abundance of Acetobacter, and restoration of A. tropicalis in germ-free flies reduced atrazine toxicity bacterium contained genes involved in atrazine metabolism (by 16S rRNA gene sequencing) | [80] |
| Serratia marcescens and Pseudomonas protegens in Nasonia vitripennis | Bacterial densities of S. marcescens and P. protegens in atrazine-fed N. vitripennis significantly increased, and degradation rates to atrazine were 20% and 10%, respectively; both contained genes involved in atrazine metabolism (by 16S rRNA gene sequencing, HPLC, whole-genome sequencing) | [24] | |
| Stenotrophomonas maltophilia in Bombyx mori | Chlorpyrifos | Enhanced host resistance to chlorpyrifos by increasing activities of acetylcholinesterase (by 16S rRNA gene sequencing, qRT-PCR, GC-MS) | [85] |
| Aeromonas hydrophila in Culex pipiens | Deltamethrin | Increased the resistance of hosts to deltamethrin by enhancing activities of CYP450s (measurement of activity levels of enzyme) | [86] |
| Enterococcus sp. in Plutella xylostella | Chlorpyrifos | Enhanced insecticide resistance to chlorpyrifos by regulating expression of antimicrobial peptide named gloverin (by using a UV spectrophotometer at 293 nm absorbance and qRT-PCR) | [87] |
| Wolbachia in Nilaparvata lugens | Imidacloprid | Enhanced resistance of hosts to imidacloprid by promoting expression of NlCYP4CE1 (by 16S rRNA gene sequencing, qRT-PCR, measurement of activity levels of enzyme) | [88] |
| gut bacteria in Apis mellifera | Thiacloprid, tau-fluvalinate and flumethrin | E=Enhanced insecticide resistance of hosts by promoting expression of immune-related genes and detoxification-related genes (by 16S rRNA gene sequencing, qRT-PCR, HPLC) | [89,90] |
4. Degradation of Other Detrimental Substances by Insect Bacteria
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Engel, M.S. Insect evolution. Curr. Biol. 2015, 25, 868–872. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Feng, J.C.; Xue, D.Y.; Sang, W.G.; Axmacher, J. Insect diversity: Addressing an important but strongly neglected research topic in China. J. Resour. Ecol. 2011, 4, 380–384. [Google Scholar]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.L.; Keesey, L.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222, 1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizé, A.; Mckay, R.; Lewis, Z. Kin recognition in Drosophila: The importance of ecology and gut microbiota. ISME J. 2014, 8, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farine, J.P.; Habbachi, W.; Cortot, J.; Roche, S.; Ferveur, J.F. Maternally-transmitted microbiota affects odor emission and preference in Drosophila larva. Sci. Rep. 2017, 7, 6062. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xian, X.Q.; Gu, Y.J.; Castane, C.; Arno, J.; Wu, S.; Wan, F.H.; Liu, W.X.; Zhang, G.F.; Zhang, Y.B. Similar bacterial communities among different populations of a newly emerging invasive species, Tuta absoluta (Meyrick). Insects 2022, 13, 252. [Google Scholar] [CrossRef]
- Yang, H.; Huang, Y.P. Insect microbiome: As guardians of insect health and adaptation. Acta Microbiol. Sinica. 2018, 58, 961–962. [Google Scholar]
- Dong, S.Z.; Pang, K.; Bai, X.; Yu, X.P.; Hao, P.Y. Identification of two species of yeast-like symbiotes in the brown planthopper, Nilaparvata lugens. Curr. Microbiol. 2011, 62, 1133–1138. [Google Scholar] [CrossRef]
- Vogel, K.J.; Moran, N.A. Functional and evolutionary analysis of the genome of an obligate fungal symbiont. Genome Biol Evol. 2013, 5, 891–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, K.; Dong, S.Z.; Hao, P.Y.; Chen, T.T.; Wang, X.L.; Yu, X.P.; Lin, H.F. Fungicides reduce the abundance of yeast-like symbionts and survival of white-backed planthopper Sogatelle furcifera (Homoptera: Delphacidae). Insects 2020, 11, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strand, M.R.; Burke, G.R. Polydnavirus-wasp associations: Evolution, genome organization, and function. Curr. Opin. Virol. 2013, 3, 587–594. [Google Scholar] [CrossRef]
- Xu, P.J.; Yang, L.Y.; Yang, X.M.; Li, T.; Graham, R.I.; Wu, K.M.; Wilson, K. Novel partiti-like viruses are conditional mutualistic symbionts in their normal lepidopteran host, African armyworm, but parasitic in a novel host, Fall armyworm. PLoS Pathog. 2020, 16, 1008467. [Google Scholar] [CrossRef] [PubMed]
- Ohkuma, M. Symbioses of flagellates and prokaryotes in the gut of lower termites. Trends Microbiol. 2008, 16, 345–352. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, Z.; Han, S.; Fan, S.; Yang, H. Phylogenetic diversity of Archaea in the intestinal tract of termites from different lineages. J. Basic Microbiol. 2015, 55, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, P.; Moran, N.A. Functional and evolutionary insights into the simple yet specific gut microbiota of the honey bee from metagenomic analysis. Gut Microbes. 2013, 4, 60–65. [Google Scholar] [CrossRef]
- Dantur, K.I.; Enrique, R.; Welin, B.; Castagnaro, A.P. Isolation of cellulolytic bacteria from the intestine of Diatraea saccharalis larvae and evaluation of their capacity to degrade sugarcane biomass. AMB Express. 2015, 5, 15. [Google Scholar] [CrossRef] [Green Version]
- Dillon, R.; Charnley, K. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Shao, Y.Q.; Chen, B.S.; Sun, C.; Ishida, K.; Hertweck, C.; Boland, W. Symbiont-derived antimicrobials contribute to the control of the lepidopteran gut microbiota. Cell. Chem. Biol. 2017, 24, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Powell, J.E.; Steele, M.I.; Dietrich, C.; Moran, N.A. Honeybee gut microbiota promotes host weight gain via bacterial metabolism and hormonal signaling. Proc. Natl. Acad. Sci. USA 2017, 114, 4775–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Q.; Cao, L.J.; Sun, L.N.; Chen, J.C.; Gong, Y.J.; Pu, D.Q.; Huang, Q.; Hoffmann, A.A.; Wei, S.J. Similar gut bacterial microbiota in two fruit-feeding moth pests collected from different host species and locations. Insects 2020, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.H.; Berdy, B.M.; Velasquez, O.; Jovanovic, N.; Alkhalifa, S.; Minbiole, K.P.C.; Brucker, R.M. Changes in microbiome confer multigenerational host resistance after sub-toxic pesticide exposure. Cell Host Microbe. 2020, 27, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Shapira, M. Gut microbiotas and host evolution: Scaling up symbiosis. Trends Ecol. Evol. 2016, 31, 539–549. [Google Scholar] [CrossRef]
- Cao, L.; Ning, K. Metagenomics of the insect gut: The frontier of microbial big data. Acta Microbiol. Sinica. 2018, 58, 964–984. [Google Scholar]
- Zhang, Z.J.; Mu, X.H.; Cao, Q.N.; Shi, Y.; Hu, X.S.; Zheng, H. Honeybee gut Lactobacillus modulates host learning and memory behaviors via regulating tryptophan metalolism. Nat. Commun. 2022, 13, 2037. [Google Scholar] [CrossRef]
- Bai, L.; Wang, L.L.; Vega-Rodriguez, J.; Wang, G.D.; Wang, S.B. A gut symbiotic bacterium Serratia marcescens renders mosquito resistance to Plasmodium infection through activation of mosquito immune responses. Front. Microbiol. 2019, 10, 1580. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstein, S.R.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microbe. 2021, 29, 879–893. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Zhang, C.X.; Chen, R.Z.; He, S.Y. Challenging battles of plants with phloem-feeding insects and prokaryotic pathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 23390–23397. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Auad, A.M.; Mendes, S.M.; Frizzas, M.R. Crop losses and the economic impact of insect pests on Brazilian agriculture. Crop. Prot. 2014, 56, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Alyokhin, A.; Chen, Y.H. Adaptation to toxic hosts as a factor in the evolution of insecticide resistance. Curr. Opin. Insect Sci. 2017, 21, 33–38. [Google Scholar] [CrossRef]
- Hammer, T.J.; Bowers, M.D. Gut microbes may facilitate insect herbivory of chemically defended plants. Oecologia 2015, 179, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Consales, F.; Schweizer, F.; Erb, M.; Gouhier-Darimont, C.; Bodenhausen, N.; Bruessow, F.; Sobhy, I.; Reymond, P. Insect oral secretions suppress wound-induced responses in Arabidopsis. J. Exp. Bot. 2012, 63, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atamian, H.S.; Chaudhary, R.; Cin, V.D.; Bao, E.; Girke, T.; Kaloshian, I. In planta expression or delivery of potato aphid Macrosiphum euphorbiae effectors Me10 and Me23 enhances aphid fecundity. Mol. Plant Microbe Interact. 2013, 26, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.H.; Rosa, C.; Scully, E.D.; Peiffer, M.; Tooker, J.F.; Hoover, K.; Luthe, D.S.; Felton, G.W. Herbivore exploits orally secreted bacteria to suppress plant defenses. Proc. Natl. Acad. Sci. USA 2013, 110, 15728–15733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Garcia, D.; Mestre-Rincon, N.; Zchori-Fein, E.; Morin, S. Inside out: Microbiota dynamics during host-plant adaptation of whiteflies. IMSE J. 2020, 14, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Leite-Mondin, M.; Dilegge, M.J.; Manter, D.K.; Weir, T.L.; Silva-Filho, M.C.; Vivanco, J.M. The gut microbiota composition of Trichoplusia ni is altered by diet and may influence its polyphagous behavior. Sci. Rep. 2021, 11, 5786. [Google Scholar] [CrossRef]
- Vilanova, C.; Baixeras, J.; Latorre, A.; Porcar, M. The generalist inside the specialist: Gut bacterial communities of two insect species feeding on toxic plants are dominated by Enterococcus sp. Front. Microbiol. 2016, 7, 1005. [Google Scholar] [CrossRef] [Green Version]
- Boone, K.C.; Keefover-Ring, K.; Mapes, A.C.; Adams, A.S.; Bohlmann, J.; Raffa, K.F. Bacteria associated with a tree-killing insect reduce concentrations of plant defense compounds. J. Chem. Ecol. 2013, 39, 1003–1006. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.S.; Aylward, F.O.; Adams, S.M.; Erbilgin, N.; Aukema, B.H.; Currie, C.R.; Suen, G.; Raffa, K.F. Mountain pine beetles colonizing historical and naïve host trees are associated with a bacterial community highly enriched in genes contributing to terpene metabolism. Appl. Environ. Microb. 2013, 79, 3468–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.T.; Lu, M.; Sun, J.H. Invasive bark beetle-associated microbes degrade a host defensive monoterpene. Insect Sci. 2016, 23, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Berasategui, A.; Salem, H.; Paetz, C.; Santoro, M.; Gershenzon, J.; Kaltenpoth, M.; Schmidt, A. Gut microbiota of the pine weevil degrades conifer diterpenes and increases insect fitness. Mol. Ecol. 2017, 26, 4099–4110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.K.; Shu, J.P.; Xue, H.J.; Zhang, W.; Zhang, Y.B.; Liu, Y.N.; Fang, L.X.; Wang, Y.D.; Wang, H.J. The gut microbiota in camellia weevils are influenced by plant secondary metabolites and contribute to saponin degradation. Msystems 2020, 5, e00692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, C.S.; Figueiredo, M.B.; Moraes, C.S.; Pereira, S.B.; Dyson, P.; Mello, C.B.; Castro, D.P.; Azambuja, P. Azadirachtin interferes with basal immunity and microbial homeostasis in the Rhodnius prolixus midgut. Dev. Comp. Immunol. 2021, 114, 103864. [Google Scholar] [CrossRef]
- Nuringtyas, T.R.; Verpoorte, R.; Klinkhamer, P.G.L.; van Oers, M.M.; Leiss, K.A. Toxicity of pyrrolizidine alkaloids to Spodoptera exigua using insect cell lines and injection bioassays. J. Chem. Ecol. 2014, 40, 609–616. [Google Scholar] [CrossRef]
- Ceja-Navarro, J.A.; Vega, F.E.; Karaoz, U.; Hao, Z.; Jenkins, S.; Lim, H.C.; Kosina, P.; Infante, F.; Northen, T.R.; Brodie, E.L. Gut microbiota mediate caffeine detoxification in the primary insect pest of coffee. Nat. Commun. 2015, 6, 7618. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic bacteria enable olive fly larvae to overcome host defences. R. Soc. Open Sci. 2015, 2, 150170. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.H.; Wickham, J.D.; Chen, L.; Xu, D.D.; Lu, M.; Sun, J.H. Bacterial microbiota protect an invasive bark beetle from a pine defensive compound. Microbiome 2018, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Mason, C.J.; Lowe-Power, T.M.; Rubert-Nason, K.F.; Lindroth, R.L.; Raffa, K.F. Interactions between bacteria and aspen defense chemicals at the phyllosphere-herbivore interface. J. Chem. Ecol. 2016, 42, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.P.; Beran, F. Gut microbiota degrades toxic isothiocyanates in a flea beetle pest. Mol. Ecol. 2020, 29, 4692–4705. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Hosokawa, T.; Oshima, K.; Hattori, M.; Fukatsu, T. Reductive evolution of bacterial genome in insect gut environment. Genome Biol. Evol. 2011, 3, 702–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al, K.F.; Daisley, B.A.; Chanyi, R.M.; Bjazevic, J.; Razvi, H.; Reid, G.; Burton, J.P. Oxalate-degrading Bacillus subtilis mitigates urolithiasis in a Drosophila melanogaster model. Msphere 2020, 5, e00498. [Google Scholar] [CrossRef]
- Zeng, J.Y.; Wu, D.D.; Shi, Z.B.; Yang, J.; Zhang, G.C.; Zhang, J. Influence of dietary aconitine and nicotine on the gut microbiota of two lepidopteran herbivores. Arch. Insect Biochem. Physiol. 2020, 104, e21676. [Google Scholar] [CrossRef]
- Engl, T.; Kaltenpoth, M. Influence of microbial symbionts on insect pheromones. Nac. Prod. Rep. 2018, 35, 386–397. [Google Scholar] [CrossRef]
- Pizzolante, G.; Cordero, C.; Tredici, S.M.; Vergara, D.; Pontieri, P.; Giudice, L.D.; Capuzzo, A.; Rubiolo, P.; Kanchiswamy, C.N.; Zebelo, S.A.; et al. Cultivable gut bacteria provide a pathway for adaptation of Chrysolina herbacea to Mentha aquatic volatiles. BMC Plant Biol. 2017, 17, 30. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Ma, Y.G.; Xie, M.X.; Lu, Y.Y.; Cheng, D.F. Rectal bacteria produce sex pheromones in the male oriental fruit fly. Curr. Biol. 2021, 31, 2220–2226. [Google Scholar] [CrossRef]
- Xu, L.; Wang, J.H.; Mei, Y.; Li, D.Z. Research progress on the molecular mechanisms of insecticides resistance mediated by detoxification enzymes and transporters. Chin. J. Pestic. Sci. 2020, 22, 1–10. [Google Scholar]
- Mallinger, R.E.; Werts, P.; Gratton, C. Pesticide use within a pollinator-dependent crop has negative effects on the abundance and species richness of sweat bees, Lasioglossum spp., and on bumble bee colony growth. J. Insect Conserv. 2015, 19, 999–1010. [Google Scholar] [CrossRef]
- Obregon, D.; Guerrero, O.R.; Stashenko, E.; Poveda, K. Natural habitat partially mitigates negative pesticide effects on tropical pollinator communities. Glob. Ecol. Conserv. 2021, 28, e01668. [Google Scholar] [CrossRef]
- Mills, N.J.; Beers, E.H.; Shearer, P.W.; Unruh, T.R.; Amarasekare, K.G. Comparative analysis of pesticide effects on natural enemies in western orchards: A synthesis of laboratory bioassay data. Biol. Control. 2016, 102, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Pestana, D.; Teixeira, D.; Faria, A.; Domingues, V.; Monteiro, R.; Calhau, C. Effects of the environmental pesticide DDT and its metabolites on the human breast cancer cell line MCF-7. Toxicol. Lett. 2010, 196, 180. [Google Scholar] [CrossRef]
- Edgerton, M.D.; Fridgen, J.; Anderson, J.R., Jr.; Ahlgrim, J.; Criswell, M.; Dhungana, P.; Gocken, T.; Li, Z.; Mariappan, S.; Pilcher, C.D.; et al. Transgenic insect resistance traits increase corn yield and yield stability. Nat. Biotechnol. 2012, 30, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.D. Detection and mechanisms of resistance evolved in insects to Cry toxins from Bacillus thuringiensis. Adv. Insect Physiol. 2014, 47, 297–342. [Google Scholar]
- Jakka, S.R.K.; Gong, L.; Hasler, J.; Banerjee, R.; Sheets, J.J.; Narva, K.; Blanco, C.A.; Jurat-Fuentes, J.L. Field-evolved mode 1 fall armyworm resistance to Bt corn associated with reduced Cry1Fa toxin binding and midgut alkaline phosphatase expression. Appl. Environ. Microb. 2015, 82, 02871-15. [Google Scholar]
- Guo, L.; Wang, Y.; Zhou, X.G.; Li, Z.Y.; Liu, S.Z.; Pei, L.; Gao, X.W. Functional analysis of a point mutation in the ryanodine receptor of Plutella xylostella (L.) associated with resistance to chlorantraniliprole. Pest Manag. Sci. 2014, 70, 1083–1089. [Google Scholar] [CrossRef]
- Wang, X.L.; Cao, X.W.; Jiang, D.; Yang, Y.H.; Wu, Y.D. CRISPR/Cas9 mediated ryanodine receptor I4790M knockin confers unequal resistance to diamides in Plutella xylostella. Insect Biochem. Mol. Biol. 2020, 125, 103453. [Google Scholar] [CrossRef]
- Li, X.X.; Li, R.; Zhu, B.; Gao, X.W.; Liang, P. Overexpression of cytochrome P450 CYP6BG1 may contribute to chlorantraniliprole resistance in Plutella xylostella (L.). Pest Manag. Sci. 2018, 74, 1386–1393. [Google Scholar] [CrossRef]
- Hou, J.Y.; Yu, J.Z.; Qin, Z.H.; Liu, X.J.; Zhao, X.P.; Hu, X.Q.; Yu, R.X.; Wang, Q.; Yang, J.Y.; Shi, Y.; et al. Guadipyr, a new insecticide, induces microbiota dysbiosis and immune disorders in the midgut of silkworms (Bombyx mori). Environ. Pollut. 2021, 286, 117531. [Google Scholar] [CrossRef]
- Wang, Y.T.; Shen, R.X.; Xing, D.; Zhao, C.P.; Gao, H.T.; Wu, J.H.; Zhang, N.; Zhang, H.D.; Chen, Y.; Zhao, T.Y.; et al. Metagenome sequencing reveals the midgut microbiota makeup of Culex pipiens quinquefasciatus and its possible relationship with insecticide resistance. Front. Microbiol. 2021, 12, 625539. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Zhang, C.X.; Cheng, P.; Wang, Y.; Liu, H.M.; Wang, H.F.; Wang, H.W.; Gong, M.Q. Differences in the intestinal microbiota between insecticide-resistant and -sensitive Aedes albopictus based on full-length 16S rRNA sequencing. MicrobiologyOpen 2021, 10, 1177. [Google Scholar] [CrossRef] [PubMed]
- Arévalo-Cortés, A.; Mejia-Jaramillo, A.M.; Granada, Y.; Coatsworth, H.; Lowenberger, C.; Triana-Chavez, O. The midgut microbiota of colombian Aedes aegypti populations with different levels of resistance to the insecticide lambda-cyhalothrin. Insects 2020, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- Gressel, J. Microbiome facilitated pest resistance: Potential problems and uses. Pest Manag. Sci. 2018, 74, 511–515. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, L.G.; de Moraes, L.A.B.; Trigo, J.R.; Omoto, C.; Consoli, F.L. The gut microbiota of insecticide-resistant insects houses insecticide-degrading bacteria: A potential source for biotechnological exploitation. PLoS ONE 2017, 12, e0174754. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [Green Version]
- Ishigami, K.; Jang, S.; Itoh, H.; Kikuchi, Y. Insecticide resistance governed by gut symbiosis in a rice pest, Cletus punctiger, under laboratory conditions. Biol. Lett. 2021, 17, 20200780. [Google Scholar] [CrossRef]
- Ozdal, M.; Ozdal, O.G.; Alguri, O.F. Isolation and characterization of alpha-endosulfan degrading bacteria from the microflora of cockroaches. Pol. J. Microbiol. 2016, 65, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Omoke, D.; Kipsum, M.; Otieno, S.; Esalimba, E.; Sheth, M.; Lenhart, A.; Njeru, E.M.; Ochomo, E.; Dada, N. Western Kenyan Anopheles gambiae showing intense permethrin resistance harbour distinct microbiota. Malar. J. 2021, 20, 77. [Google Scholar] [CrossRef]
- Brown, J.B.; Langley, S.A.; Snijders, A.M.; Wan, K.H.; Morris, S.N.S.; Booth, B.W.; Fisher, W.W.; Hammonds, A.S.; Park, S.; Weiszmann, R.; et al. An integrated host-microbiome response to atrazine exposure mediates toxicity in Drosophila. Commun. Biol. 2021, 4, 1324. [Google Scholar] [CrossRef]
- Cheng, D.F.; Guo, Z.J.; Riegler, M.; Xi, Z.Y.; Liang, G.W.; Xu, Y.J. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramya, S.L.; Venkatesan, T.; Murthy, K.S.; Jalali, S.K.; Verghese, A. Detection of carboxylesterase and esterase activity in culturable gut bacterial flora isolated from diamondback moth, Plutella xylostella (Linnaeus), from India and its possible role in indoxacarb degradation. Braz. J. Microbiol. 2016, 47, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Wang, W.F.; Lu, Y.J. Symbiotic microbiota and insecticide resistancein insects. Chin. J. Appl. Entomol. 2021, 58, 265–276. [Google Scholar]
- Liu, X.D.; Guo, H.F. Importance of endosymbionts Wolbachia and Rickettsia in insect resistance development. Curr. Opin. Insect Sci. 2019, 33, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.S.; Zhang, N.; Xie, S.; Zhang, X.C.; He, J.T.; Muhammad, A.; Sun, C.; Lu, X.M.; Shao, Y.Q. Gut bacteria of the silkworm Bombyx mori facilitate host resistance against the toxic effects of organophosphate insecticides. Environ. Int. 2020, 143, 105886. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.F.; Liu, Z.H.; Zhang, R.M.; Zhou, D.; Sun, Y.; Ma, L.; Shen, B. Effect of the midgut symbiotic Aeromonas hydrophila on the deltamethrin resistance of Culex pipiens pallens. J. Pathog. Biol. 2021, 16, 661–666. [Google Scholar]
- Xia, X.F.; Sun, B.T.; Gurr, G.M.; Vasseur, L.; Xue, M.Q.; You, M.S. Gut microbiota mediate insecticide resistance in the diamondback moth, Plutella xylostella (L.). Front. Microbiol. 2018, 9, 00025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, T.W.; Zhang, Y.H.; Liu, Y.; Deng, X.Q.; He, S.; Li, J.H.; Wang, H. Wolbachia enhances expression of NICYP4CE1 in Nilaparvata lugens in response to imidacloprid stress. Insect Sci. 2021, 28, 355–362. [Google Scholar] [CrossRef]
- Wu, Y.Q.; Zheng, Y.F.; Chen, Y.N.; Wang, S.; Chen, Y.P.; Hu, F.L.; Zheng, H.Q. Honey bee (Apis mellifera) gut microbiota promotes host endogenous detoxification capability via regulation of P450 gene expression in the digestive tract. Microb. Biotechnol. 2020, 13, 1201–1212. [Google Scholar] [CrossRef]
- Yu, L.T.; Yang, H.Y.; Cheng, F.P.; Wu, Z.H.; Huang, Q.; He, X.J.; Yan, W.Y.; Zhang, L.Z.; Wu, X.B. Honey bee Apis mellifera larvae gut microbial and immune, detoxication responses towards flumethrin stress. Environ. Pollut. 2021, 290, 118107. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Lepp, D.; Shao, S.Q.; Zhou, T. Strategies and methodologies for developing microbial detoxification systems to mitigate mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.W.; Zhou, T.; Young, J.C.; Boland, G.J.; Scott, P.M. Chemical and biological transformations for detoxification of trichothecene mycotoxins in human and animal food chains: A review. Trends Food Sci. Technol. 2010, 21, 67–76. [Google Scholar] [CrossRef]
- Zhu, Y.; Hassan, Y.I.; Watts, C.; Zhou, T. Innovative technologies for the mitigation of mycotoxins in animal feed and ingredients—A review of recent patents. Anim. Feed Sci. Technol. 2016, 216, 19–29. [Google Scholar] [CrossRef]
- He, J.W.; Hassan, Y.I.; Perilla, N.; Li, X.Z.; Boland, G.J.; Zhou, T. Bacterial epimerization as a route for deoxynivalenol detoxification: The influence of growth and environmental conditions. Front. Microbiol. 2016, 7, 572. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, R.R.; He, K.L.; Bai, S.X.; Zhang, T.T.; Cong, B.; Wang, Z.Y. Effects of Conogethes punctiferalis (Lepidopteran: Grambidae) infestation on the occurrence of Fusarium ear rot and the yield loss of spring corn. Acta Entomol. Sinica. 2017, 60, 576–581. [Google Scholar]
- Shen, S.K.; Dowd, P.F. Detoxification spectrum of the cigarette beetle symbiont Symbiotaphrina kochii in culture. Entomol. Exp. Appl. 1991, 60, 51–59. [Google Scholar] [CrossRef]
- Bosch, G.; Fels-Klerx, H.J.; Rijk, T.C.; Oonincx, D.G. Aflatoxin B1 tolerance and accumulation in black soldier fly larvae (Hermetia illucens) and yellow mealworms (Tenebrio molitor). Toxins 2017, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Cong, Y.L. Global health ethical reflection on antibiotics abuse. Chin. Med. Ethics 2017, 30, 412–416. [Google Scholar]
- Liu, C.C.; Yao, H.Y.; Chapman, S.J.; Su, J.Q.; Wang, C.W. Changes in gut bacterial communities and the incidence of antibiotic resistance genes during degradation of antibiotics by black soldier fly larvae. Environ. Int. 2020, 142, 105834. [Google Scholar] [CrossRef]
- Liu, C.C.; Yao, H.Y.; Wang, C.W. Black soldier fly larvae can effectively degrade oxytetracycline bacterial residue by means of the gut bacterial community. Front. Microbiol. 2021, 12, 663972. [Google Scholar] [CrossRef]
- Jiang, C.L.; Li, H.Y.; Wang, X.; Teng, C.Y.; Feng, S.Y.; Lou, L.P.; Zhang, Z.J. Effects of fly maggot gut microbiota on the degradation of residual antibiotics in pig manure and its resistance genes. Acta Microbiol. Sin. 2018, 58, 1103–1115. [Google Scholar]
- Guzman, J.; Vilcinskas, A. Bacteria associated with cockroaches: Health risk or biotechnological opportunity? Appl. Microbiol. Biotechnol. 2020, 104, 10369–10387. [Google Scholar] [CrossRef] [PubMed]
| Plant Allelochemical | Functional Bacteria and Host | Description | Reference | |
|---|---|---|---|---|
| Terpenoid | Monoterpene | Serritia marcescens, Pseudomonas mandelii, and Rahnella aquatilis from Dendroctonus ponderosae | S. marcescens reduced 49–79% of 3-carene and (−)-β-pinene, and P. mandelii decreased concentrations of all monoterpenes by 15–24%, while R. aquatilis decreased (−)-α-pinene (38%) and (+)-α-pinene (46%) by 40% and 45% (by GC-MS), respectively | [41] |
| Pseudomonas, Rahnella, Serratia, and Burkholderia in D. ponderosae | Genera contained most genes involved in terpene degradation (by metagenomics) | [42] | ||
| Serratia sp., Pseudomonas sp., and Rahnella aquatilis in Dendroctonus valens | Degraded 20–50% of α-pinene (by GC-MS) | [43] | ||
| Diterpene | gut microbiota of Hylobius abietis | Gut bacterial community of H. abietis reduced most diterpenes, and metagenomic analysis results showed gut community contained 10 degradation genes (dit) (by metagenome sequencing and GC-MS) | [44] | |
| Saponin | Acinetobacter sp. in Curculio chinensis | Acinetobacter sp. in C. chinensis enriched after treating with saponin, and when incubating bacteria with saponin for 72 h, saponin content significantly decreased from 4.054 to 1.867 mg/mL (by 16S rRNA metagenome sequencing and HPLC) | [45] | |
| Azadirachtin | Serratia marcescens in Rhodnius prolixus | S. marcescens load in R. prolixus increased when fed diet containing azadirachtin at 1 μg/mL (by qRT-PCR) | [46] | |
| Alkaloid | Caffeine | Pseudomonas fulva in Hypothenemus hampei | P. fulva processed gene coding one subunit of caffeine demethylase, and reinstatement of P. fulva in germ-free H. hampei degraded all caffeine consumed (by 16S rRNA gene sequencing and GC-MS) | [48] |
| Aconitine, nicotine | entire gut bacteria of Dendrolimus superans and Lymantria dispar | Abundance of genus Pseudomonas in D. superans larvae increased, but Serratia and Enterobacter decreased, and L. dispar larvae fed on aconitine-treated diet and nicotine-treated diet shared dominant bacteria Enterococcus (by 16S rRNA gene sequencing) | [55] | |
| Phenol | Phenolic glycoside | Erwinia dacicola in Bactrocera olea | Larvae developed in unripe olive harbored more E. dacicola (by 16S rRNA gene sequencing) | [49] |
| Phenolic naringenin | Novosphingobium sp. in D. valens | Novosphingobium sp. possesses putative genes involved in degradation of naringenin, and D. valens supplied with Novosphingobium sp. acquired protection against naringenin (by metagenomic analysis) | [50] | |
| Tannins | Acinetobacter sp. in Lymantria dispar | Condensed tannins improved growth of Acinetobacter sp. by 15% (by measuring the optical density) | [51] | |
| Glucosinolate | Pantoea sp. Pc8 in Psylliodes chrysocephala | Laboratory-reared and field-collected P. chrysocephala all contained three core genera Pantoea, Acinetobacter and Pseudomonas, and reintroduction of Pantoea sp. Pc8 in antibiotic-fed beetles restored isothiocyanate degradation ability in vivo (by 16S rRNA gene sequencing and LC-MS) | [52] | |
| Oxalate | Ishikawaella capsulata in Megacopta punctatissima | Encodes genes of oxalate decarboxylase (by whole-genome shotgun sequencing) | [53] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, M.; Lin, X.; Guo, X. The Role of Insect Symbiotic Bacteria in Metabolizing Phytochemicals and Agrochemicals. Insects 2022, 13, 583. https://doi.org/10.3390/insects13070583
Zhao M, Lin X, Guo X. The Role of Insect Symbiotic Bacteria in Metabolizing Phytochemicals and Agrochemicals. Insects. 2022; 13(7):583. https://doi.org/10.3390/insects13070583
Chicago/Turabian StyleZhao, Man, Xingyu Lin, and Xianru Guo. 2022. "The Role of Insect Symbiotic Bacteria in Metabolizing Phytochemicals and Agrochemicals" Insects 13, no. 7: 583. https://doi.org/10.3390/insects13070583
APA StyleZhao, M., Lin, X., & Guo, X. (2022). The Role of Insect Symbiotic Bacteria in Metabolizing Phytochemicals and Agrochemicals. Insects, 13(7), 583. https://doi.org/10.3390/insects13070583