Simple Summary
Acibenzolar-S-methyl is an analogue of salicylic acid, and it is known as a plant elicitor able to induce plant endogenous defences against plant pathogens. Recently, it has been shown to also affect phloem-feeder pests, even though the mechanism is still unclear. Pear psylla (Cacopsylla pyri) is a serious threat for pear production in Europe, and its control is usually based on the use of chemical insecticides. The development of novel innovative control approaches is becoming more and more important, especially in integrated pest management. The present work investigated the possible indirect influence of acibenzolar-S-methyl, through the expression of pear Pathogenesis-Related protein (PR) coding genes, on the probing behaviour and on the survival of C. pyri nymphs and adults feeding on pear potted plants. The minor effects observed on the pest would suggest that acibenzolar-S-methyl cannot be used against psyllas, but it might be recommended on pear orchards in the primary control of other targets such as fire blight disease.
Abstract
European pear psylla, Cacopsylla pyri, is one of the worst pests of pear in Europe. We investigated whether acibenzolar-S-methyl (ASM) application on pear plants might affect the behaviour in C. pyri. The elicitor was applied on pear potted plants, and after 48 h, we confirmed the ASM-mediated induction of several Pathogenesis-Related protein (PR) coding genes. At the same time, an in-depth analysis was performed on the probing behaviour of adults and nymphs of C. pyri on ASM-treated pear plants by the EPG-DC system, as well as the assessment of young nymphs’ survival 7 days after the ASM application. The elicitor application weakly interfered with C. pyri nymphs probing behaviour and survival, while it did not affect adult stages. These data confirm previous observations obtained on C. pyricola and suggest that the elicitor does not represent a viable tool in the control of pear psylla species, especially if used alone, but it might be used in integrated management strategies focused on other plant pathogens such as Erwinia amylovora.
1. Introduction
The pear psylla Cacopsylla pyri L. (Hemiptera: Psyllidae) is one of the most important pests of the common pear (Pyrus communis L.) across Europe [1]. The damage is mainly due to the honeydew excreted by nymphs, which harms leaf tissues and creates fruits russeting. Honeydew also acts as an optimal growth medium for black sooty moulds, whose presence on fruits drastically reduces their market value [2]. In addition, C. pyri is a vector for “Candidatus Phytoplasma pyri” responsible for the Pear Decline disease (PD) [3], a severe condition that reduces tree vigour and may even be lethal for trees.
During the last two decades the control programs based on integrated pest management (IPM) were able to obtain great results in containing C. pyri damage in European pear orchards. This IPM strategy was based on the initial use of conventional insecticides (abamectin and spirotetramat) followed by the natural control exerted by enemies such as the predator Anthocoris nemoralis, unaffected by the chemicals used. The chemical management of the psyllid proves to be very effective because of very low pest resistance toward the active ingredients [4,5,6].
In the last few years, the excessive application of nonselective insecticides to control the massive invasion of orchards by the alien marmorated stink bug, Halyomorpha halys, has induced a decrease in C. pyri natural predators thus triggering delayed near-harvest outbreaks that have often caused significant damage to pear production [7]. On the other hand, abamectin is soon to be banned in the European Union, further limiting the number of products that can be used in the orchard. Therefore, new tools need to be developed in the near future.
A promising strategy to control C. pyri involves the host plant’s innate resistance mechanisms [8]. Nin and colleagues maintain that all main P. communis cultivars are still susceptible to all pear psylla species and despite the long-term worldwide breeding programs, no commercially resistant cultivar has been released.
Conversely, some researchers have suggested the use of plant resistance elicitors, such as harpins, chitosans, and acibenzolar-S-methyl (ASM), to control C. pyricola [9,10,11]. Among these compounds, the ASM, a synthetic analogue of salicylic acid (SA), has no direct effect against pests but causes the activation and accumulation of several pest-resistant proteins and molecules in the treated plants [12,13,14]. Among these, several PR (pathogenesis-related) proteins, upregulated by ASM treatment, show different chemical and physical properties such as chitinase (PR3 and PR8), glucanase (PR2), and antimicrobial activities (antifungal PR1 and osmotin/thaumatin-like PR5). [15,16].
ASM has been successfully applied against pear diseases including fire blight, caused by Erwinia amylovora, due to its ability to induce Systemic Acquired Resistance (SAR). Therefore, it has been registered for foliar application to control fire blight and other pear pathogens [17,18,19,20]. The possibility of using ASM to activate plant defences against phytophagous insects has proved uncertain [21,22]. On one hand, ASM was shown to induce defence responses against the phloem-feeding green peach aphid, Myzus persicae, on tomato [23,24,25]. On the other hand, Cooper and Horton [9,10] have shown that ASM application might affect congeneric C. pyricola nymphs’ infestations in the northeastern USA, although they did not provide evidence of the plant responses activated by the elicitor and did not suggest a possible mechanism of action that might explain how this elicitor affected the pests. Furthermore, Orpet and colleagues [11] reported that treating pear plants with harpin or ASM did not cause a long-term induction of the PR1 gene or a reduction in the C. pyricola population.
Our study aims to investigate whether short-term ASM treatment (as verified by induction of several defence genes) can interfere with C. pyri probing behaviour and nymphs’ survival in pear plants.
2. Materials and Methods
2.1. Source of Insects and Plant Material
All C. pyri adult winter forms were field collected and directly used for EPG-DC recording. Otherwise, reared C. pyri nymph and adult summer forms were maintained on 1-year old potted pear plants (30 cm height) of the susceptible cv. Abbé Fétel and kept in a climate-controlled chamber at 23 ± 1 °C under a 16:8 LD photoperiod.
2.2. ASM Applications
A commercial solution of ASM (Bion® 50 WG; Syngenta Crop Protection, Milan, Italy) was dissolved in distilled water. The concentration applied was 125 mg L−1. One year old potted plants of susceptible cultivar Abbé Fétel were randomly assigned to treatment and removed from the greenhouse prior to solution application in an open field by hand atomizers. Plants were sprayed until runoff of leaves, left to dry for 1 h, and then returned to the greenhouse for 48 h before EPG-DC recording, mortality bioassays, and RNA extraction.
2.3. Gene Expression Analysis after ASM Treatment
Pear leaf samples were collected from treated and control plants and immediately frozen at −80 °C. The total RNA was extracted using the Spectrum Plant Total RNA Prep kit (SIGMA Life Science, St. Louis, MO, USA) from 100 mg of tissue. Upon DNAse treatment (ThermoFisher Scientific, Waltham, MA, USA), 1 µg of RNA was retrotranscribed with a OneScript Plus cDNA Synthesis kit (Abm, Vancouver, BC, Canada). Real-time PCR reactions were performed in a CFX Connect Real-Time (Bio-Rad, Hercules, CA, USA) containing 1.6 µL of cDNA, 6 µL of SsoAdvanced Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, USA), and 0.4 µL of each primer (10 μM) in a 12 µL final volume. The amplification conditions were 95° for 2 min, 40 cycles at 95 °C/10 s, and 60 °C/30 s. A melting curve analysis from 55 to 95 °C was applied after the amplification protocol. Gene expression was normalized using the EF1α reference gene and analysed by the Livak method [26] followed by the ANOVA nonparametric Mann-Whitney test. The primers used are shown in Table 1 (PCP sequences were obtained from the Genome Database for Rosaceae, GDR, [27]).

Table 1.
Primers used in this study. PR: Pathogenesis-Related proteins; EF1α: Elongation Factor 1 alpha.
2.4. EPG-DC Recording
The EPG-DC recording was performed in the laboratory at 21 ± 1 °C under fluorescent light (4000 Lux, HF tubes). The EPG-DC recording involved 3rd instar nymphs as well as winter and summer forms of C. pyri adults (25 individuals for each stage) singularly placed on pear plants previously (48 h) treated with ASM or water (negative control). The EPG-DC system was composed of a 100-Hz Giga 4 amplifier (Wageningen University, Wageningen, The Netherlands) with 1-GΩ resistance input [28]. A thin gold electrode wire (diameter: 20 μm; length: 2 cm) was glued to the dorsum of each C. pyri individual using silver glue, as previously described [29]. The gold electrode was connected to the amplifier by a long and thick copper wire. A second electrode, made of copper, was deeply inserted in the wet soil of the potted pear plant.
During each registration, the plants, the insects, and the amplifier were placed inside a Faraday cage to avoid background electric noise. Each EPG-DC signal was recorded for 8 h and converted from analogic to digital (A–D) through an interface device (KPCI-3102, Keithley Instruments, Cleveland, OH, USA). If one insect fell from the leaf within the first hour of recording, the insect was repositioned once. Later falls were removed from the experiment. Falls were nevertheless quite rare (below 10% on average) At the end of the recording, dead insects were noted and excluded from further analyses. All waveforms were analysed and scored by the software Stylet+ (Wageningen University, Wageningen, The Netherlands) using the following characterization: Np: non probing; PA-PB: stylet penetration (sheath salivation); PC1: parenchyma penetration (sheath salivation); PC2: vascular parenchyma (unknown activity); PD: transition to phloem sieve elements; PE1: sieve element penetration (phloem salivation activity); PE2: sieve element penetration (phloem ingestion); PG: xylem vessel penetration (xylem ingestion) [29,30,31] (Supplementary Materials Figure S1). Once marked, all the recordings were individually selected for subsequent analysis. In particular, all recordings with electrical noises or bad electric connections were discarded, as suggested by Ripamonti and colleagues [32]. All waveforms recorded for both adults and nymphs were analysed by the ANOVA nonparametric Mann-Whitney test with the software package STATISTICA for Windows version 12 (StatSoft, Tulsa, OK, USA), in accordance with the nonsequential parameters [33].
2.5. Nymphs Mortality Bioassay
The foliar spray bioassays were performed as described [4], with slight modifications, in the laboratory at 23 ± 1 °C, 65% relative humidity (r.h.), and a 16:8 h LD photoperiod. Nine pear potted plants (cv. Abbé Fétel) were sprayed by a hand atomizer with 125 mg L−1 ASM and returned to the growth chamber for 48 h. Twenty 2nd instar reared C. pyri nymphs were gently placed on five leaves (4 nymphs per leaf) of each pear potted plant for a total of 9 replicates with 180 nymphs. A batch of nine pear plants sprayed only with distilled water was used as control. The surviving nymphs were scored 10 days after the placement. Nymphs were considered alive when they could walk and produce honeydew. The number of dead nymphs was calculated by subtracting the number of surviving nymphs from the total number of nymphs placed on the plants. Mortality data were analysed by the ANOVA nonparametric Mann-Whitney test with the software package STATISTICA for Windows version 12. The ASM efficacy (corrected mortality) was obtained by the Schneider-Orelli formula [34].
3. Results
3.1. Effect of ASM Treatment on Pear Pathogenesis-Related (PR) Proteins
Pear plants, 48 h after ASM application, were firstly tested at the molecular level to confirm the elicitor effect. RT-qPCR analyses performed on RNA extracted from ASM- or water-treated pear leaves confirmed that several PR genes (PR1, PR2, PR3, PR5, and PR8, known from the literature to be induced by ASM-application in pear and other plant species [12,13,14]) were strongly upregulated by ASM, as expected, when normalized to the EF1α reference gene (Figure 1).
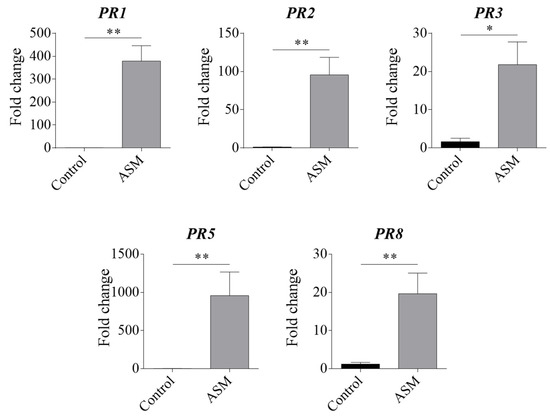
Figure 1.
Fold-change values for PR genes upregulated 48 h after ASM application, normalized with the EF1α reference gene, and analysed by the Livak method. * p < 0.05, ** p< 0.01 vs. control according to the Mann-Whitney test.
3.2. EPG-DC Recording
The EPG-DC recordings were performed to assess the effect of ASM on C. pyri probing behaviour. Probing was registered for 8 h on insects positioned on leaves 48 h after ASM or water applications. Table 2 shows that, for nymphs, the events of stylet penetration in epidermal and parenchymal tissues (along with sheath salivation) (PA, PB, and PC1 [29]) were more frequent (18.82 events/insect vs. 12.55 events/insect, p = 0.04) and shorter on ASM-treated plants as compared to the control (10.07 min/insect vs. 15.60 min/insect, p = 0.007). Once the nymph’s stylet reached the phloem, ASM treatment induced a statistically significant difference in the total duration of phloem salivation events (PE1 [29]) (41.62 min/insect vs. 16.20 min/insect, p = 0.01) and in the number of phloem salivation events (PE1, 10.09 events/insect vs. 4.86 events/insect, p = 0.003), while the duration of each phloem salivation event did not vary between the treated and control plants. Similar differences were also detected in the events related to transition to phloem (PD [29]). Other events, such as the phloem ingestion (PE2 [29]), were apparently not affected by the ASM application during the 8 h of EPG-DC recording with nymphs. On the other hand, no significant differences were observed in all parameters measured when C. pyri adults (summer and winter forms) were used in the feeding test on ASM-treated or control plants (Table 3 and Table 4).
Table 2.
Comparison of EPG-DC waveforms (mean time in minutes ± standard error of the mean, SEM, or mean number of events ± SEM) (8 h of recording) of C. pyri nymphs (n = 22 on ASM, n = 22 control) feeding on ASM- or water- (control) treated pear plants. Waveforms were classified according to [29,30,31]. * p < 0.05 vs. control according to the Mann-Whitney test.
Table 3.
Comparison of EPG-DC waveforms (mean time in minutes ± standard error of the mean, SEM, or mean number of events ± SEM) (8 h of recording) of C. pyri summer adults (n = 16 on ASM, n = 14 control) feeding on ASM- or water- (control) treated pear plants. Waveforms were classified according to [29,30,31].
Table 4.
Comparison of EPG-DC waveforms (mean time in minutes ± standard error of the mean, SEM, or mean number of events ± SEM) (8 h of recording) of C. pyri winter adults (n = 20 on ASM, n = 22 control) feeding on ASM- or water- (control) treated pear plants. Waveforms were classified according to [29,30,31].
3.3. Nymph Mortality Bioassay
The percentage of mortality of C. pyri 2nd instar nymphs feeding on ASM- or water- (control) treated pear plants is reported in Table 5. The mortality on ASM-treated plants was statistically different from the water-treated control (27.97 vs. 16.30; p = 0.009), but the ASM efficacy, expressed as corrected mortality, was nevertheless very weak (13.94%).
Table 5.
Mortality data of C. pyri nymphs on ASM-sprayed pear plants (N = 9) compared to control plants (N = 9) and percentage of ASM efficacy (corrected mortality) calculated according to [34]. ** p < 0.01 vs. control according to the Mann-Whitney test.
4. Discussion
RT-qPCR analysis confirmed that ASM was able to activate plant defences in pear plantlets. In fact, all the PR protein encoding genes tested were strongly upregulated 48 h after treatment. These data are in agreement with the ASM-induced gene upregulation described in several works in many species [13], including apple [14] and pear [12].
Nevertheless, the EPG results indicated that ASM application only weakly interfered with C. pyri nymph probing activities. In particular, ASM caused a significant increase in the mean number of stylet penetration events in nonvascular tissues and in salivation events into the phloem, once the stylet reached this tissue. Furthermore, these effects were only visible in nymphs, which were known to have a probing behaviour mainly limited to phloem [1,29,30]. Winter or summer adults’ behaviour, which is known to be less frequent in phloem [30], was unaffected by the ASM application. A previous study [25] revealed that tomato plants were less acceptable to M. persicae upon ASM application, inducing a sharp decrease in the total duration and in the number of phloem ingestions and a significant reduction in aphid fecundity [23]. These results on tomato after ASM application, as suggested by [24], might be linked to the induction of the defensive pathways, which could also protect susceptible tomato cultivars against aphid infestation.
The modest modifications of C. pyri nymphs feeding on pear plants after ASM application can explain and confirm the data obtained on C. pyricola [9,11]. ASM was able to activate a systemic defence response in pear trees causing a reduction in C. pyricola oviposition and in nymph survival, which led to a general but weak reduction in psylla population. These effects were observed in different conditions, from the laboratory to the managed or unmanaged orchards, after one or multiple ASM applications. The modest activity of ASM on C. pyri feeding as well as on C. pyricola nymphs’ survival do not warrant the use of ASM alone for the control of psyllids. However, elicitors are often used to manage pear fire blight, therefore their use might also contribute to control pear psylla populations within integrated pest management approaches. Furthermore, one cannot exclude that ASM might have an effect on the pathogen transmitted by pysllas such as the pear decline “Candidatus Phytoplasma pyri”; unfortunately, no specific work has been conducted. On the other hand, the effect of plant elicitors, such as ASM, on the resistance to a wide range of pathogens including other phytoplasms is well known [35,36,37,38]. More studies will be required to evaluate the true role of ASM in overall pear protection management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects13060525/s1, Figure S1: Schematical representation of typical C. pyri probing waveforms for winter- and summer-adults and nymphs.
Author Contributions
Conceptualization, S.C. and L.F.; data curation, S.C., D.M., L.B., L.F. and M.P.; formal analysis, S.C., D.M., L.B., L.F. and M.P.; funding acquisition, G.B.; project administration, S.C.; resources, S.C., D.M. and G.B.; supervision, S.C. and G.B.; writing—original draft, S.C. and G.B.; writing—review and editing, S.C., L.F., M.P. and G.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated and analysed during the current study are available within the article, as well as from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Civolani, S. The past and present of pear protection against the pear psylla, Cacopsylla pyri L. In Insecticides—Pest Engineering; Perveen, F., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.A.; Ortin-Angulo, M.C. Abundance and population dynamics of Cacopsylla pyri (Hemiptera: Psyllidae) and its potential natural enemies in pear orchards in Southern Spain. Crop Prot. 2012, 32, 24–29. [Google Scholar] [CrossRef]
- Seemüller, E.; Schneider, B. ‘Candidatus Phytoplasma mali’, ‘Candidatus Phytoplasma pyri’ and ‘Candidatus Phytoplasma prunorum’, the causal agents of apple proliferation, pear decline and European stone fruit yellows, respectively. Int. J. Syst. Evol. Microbiol. 2004, 54, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Civolani, S.; Peretto, R.; Caroli, L.; Pasqualini, E.; Chicca, M.; Leis, M. Preliminary resistance screening on abamectin in pear psylla (Hemiptera: Psyllidae) in Northern Italy. J. Econ. Entomol. 2007, 100, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Civolani, S.; Cassanelli, S.; Rivi, M.; Manicardi, G.C.; Peretto, R.; Chicca, M.; Pasqualini, E.; Leis, M. Survey of susceptibility to abamectin of pear psylla (Hemiptera: Psyllidae) in Northern Italy. J. Econ. Entomol. 2010, 103, 816–822. [Google Scholar] [CrossRef]
- Civolani, S.; Boselli, M.; Butturini, A.; Chicca, M.; Cassanelli, S.; Tommasini, M.G.; Aschonitis, V.; Fano, E.A. Testing spirotetramat as an alternative solution to abamectin for Cacopsylla pyri (Hemiptera: Psyllidae) control: Laboratory and field tests. J. Econ. Entomol. 2015, 108, 2737–2742. [Google Scholar] [CrossRef]
- Daugherty, M.P.; Briggs, C.J.; Welter, S.C. Bottom-up and top-down control of pear psylla (Cacopsylla pyricola): Fertilization, plant quality, and the efficacy of the predator Anthocoris nemoralis. Biol. Control 2007, 43, 257–264. [Google Scholar] [CrossRef]
- Nin, S.; Ferri, A.; Sacchetti, P.; Giordani, E. Pear resistance to psilla (Cacopsylla pyri L.): A review. Adv. Hortic. Sci. 2012, 26, 59–74. [Google Scholar]
- Cooper, R.W.; Horton, D.R. Effects of elicitors of host plant defenses on pear psylla, Cacopsylla pyricola. Entomol. Exp. Appl. 2015, 177, 300–306. [Google Scholar] [CrossRef]
- Cooper, R.W.; Horton, D.R. Elicitors of host plant defenses partially suppress Cacopsylla pyricola (Hemiptera: Psyllidae) populations under field conditions. J. Insect Sci. 2017, 17, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orpet, R.J.; Cooper, W.R.; Beers, E.H.; Nottingham, L.B. Test of plant defense elicitors for arthropod pest suppression and PR-1 gene induction in pear orchards. Entomol. Exp. Appl. 2021, 169, 1137–1146. [Google Scholar] [CrossRef]
- Johnson, K.B.; Temple, T.N. Comparison of Methods of Acibenzolar-S-Methyl application for post-infection fire blight suppression in Pear and Apple. Plant Dis. 2016, 100, 1125–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzo, F.; Faoro, F. Systemic acquired resistance (50 years after discovery): Moving from the lab to the field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Marolleau, B.; Gaucher, M.; Heintz, C.; Degrave, A.; Warneys, R.; Orain, G.; Lemarquand, A.; Brisset, M.N. When a plant resistance inducer leaves the lab for the field: Integrating ASM into routine apple protection practices. Front. Plant Sci. 2017, 8, 1938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuzun, S.; Somanchi, A. The possible role of PR proteins in multigenic and induced systemic resistance. In Multigenic and Induced Systemic Resistance in Plants; Tuzun, S., Bent, E., Eds.; Springer: New York, NY, USA, 2006; Chapter 6; pp. 112–142. [Google Scholar]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Ann. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Maxson-Stein, K.; He, S.-Y.; Hammerschmidt, R.; Jones, A.L. Effect of treating apple trees with acibenzolar-S-methyl on fire blight and expression of pathogenesis-related protein genes. Plant Dis. 2002, 86, 785–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norelli, J.L.; Jones, A.L.; Aldwinckle, H.S. Fire blight management in the twenty-first century: Using new technologies that enhance host resistance in apple. Plant Dis. 2003, 87, 756–765. [Google Scholar] [CrossRef] [Green Version]
- Sparla, F.; Rotino, L.; Valgimigli, M.C.; Pupillo, P.; Trost, P. Systemic resistance induced by benzothiadiazole in pear inoculated with the agent of fire blight (Erwinia amylovora). Sci. Hortic. 2004, 101, 269–279. [Google Scholar] [CrossRef]
- Bazzi, C.; Biondi, E.; Berardi, R.; Brunelli, A. Efficacy of bioagents and chemicals against pear shoot blight. Acta Hortic. 2006, 704, 283–288. [Google Scholar] [CrossRef]
- Inbar, M.; Doostdar, H.; Gerling, D.; Mayer, R.T. Induction of systemic acquired resistance in cotton by BTH has a negligible effect on phytophagous insects. Entomol. Exp. Appl. 2001, 99, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Bi, J.L.; Murphy, J.B.; Felton, G.W. Does salicylic acid act as a signal in cotton for induced resistance to Helicoverpa zea? J. Chem. Ecol. 1997, 23, 1805–1818. [Google Scholar] [CrossRef]
- Boughton, A.J.; Hoover, K.; Felton, G.W. Impact of chemical elicitor applications on greenhouse tomato plants and population growth of the green peach aphid, Myzus persicae. Entomol. Exp. Appl. 2006, 120, 175–188. [Google Scholar] [CrossRef]
- Cooper, W.C.; Jia, L.; Goggin, F.L. Acquired and R-gene-mediated resistance against the potato aphid in tomato. J. Chem. Ecol. 2004, 30, 2527–2542. [Google Scholar] [CrossRef] [PubMed]
- Civolani, S.; Marchetti, E.; Chicca, M.; Castaldelli, G.; Rossi, R.; Pasqualini, E.; Dindo, M.L.; Baronio, P.; Leis, M. Probing behaviour of Myzus persicae on tomato plants containing Mi gene or BTH-treated evaluated by electrical penetration graph. Bull. Insectology 2010, 63, 265–271. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jung, S.; Lee, T.; Cheng, C.H.; Buble, K.; Zheng, P.; Yu, J.; Humann, J.; Ficklin, S.P.; Gasic, K.; Scott, K.; et al. 15 years of GDR: New data and functionality in the Genome Database for Rosaceae. Nucleic Acids Res. 2018, 47, D1137–D1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjallingii, W.F. Electrical nature of recorded signals during stylet penetration by aphids. Entomol. Exp. Appl. 1985, 38, 177–186. [Google Scholar] [CrossRef]
- Civolani, S.; Leis, M.; Grandi, G.; González, E.; Pasqualini, E.; Chicca, M.; Tjallingii, W.F. Plant penetration by Cacopsylla pyri. An electrical penetration graph (EPG) study. J. Insect Physiol. 2011, 57, 1407–1419. [Google Scholar] [CrossRef]
- Civolani, S.; Grandi, G.; Chicca, M.; Pasqualini, E.; Fano, E.A.; Musacchi, S. Probing behaviour of Cacopsylla pyri on a resistant pear selection. J. Appl. Entomol. 2013, 137, 365–375. [Google Scholar] [CrossRef]
- Bonani, J.P.; Fereres, A.; Garzo, E.; Miranda, M.P.; Appezzato-Da-Gloria, B.; Lopes, J.R.S. Characterization of electrical penetration graphs of the Asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomol. Exp. Appl. 2010, 134, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Ripamonti, M.; Maron, F.; Cornara, D.; Marzachì, C.; Fereres, A.; Bosco, D. Leafhopper feeding behaviour on three grapevine cultivars with different susceptibilities to Flavescence dorée. J. Insect Physiol. 2022, 137, 104366. [Google Scholar] [CrossRef]
- Backus, E.A.; Cline, A.R.; Ellerseick, M.R.; Serrano, M.A.S. Lygus hesperus (Hemiptera: Miridae) Feeding on Cotton: New Methods and Parameters for Analysis of Nonsequential Electrical Penetration Graph Data. Ann. Entomol. Soc. Am. 2007, 100, 296–310. [Google Scholar] [CrossRef] [Green Version]
- Schneider-Orelli, O. Computation of the level of effectiveness. In Entomologisches Parktikum; 2 Auflage; Sauerländer: Aarau, Switzerland, 1947; p. 237. [Google Scholar]
- Bressan, A.; Purcell, A.H. Effect of benzothiadiazole on transmission of X-disease phytoplasma by the vector Colladonus montanus to Arabidopsis thaliana, a new experimental host plant. Plant Dis. 2005, 89, 1121–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amelio, R.; Marzachi, C.; Bosco, D. Activity of benzothiadiazole on chrysanthemum yellows phytoplasma (‘Candidatus Phytoplasma asteris’) infection in daisy plants. Crop Prot. 2010, 29, 1094–1099. [Google Scholar] [CrossRef]
- Ratchaseema, M.T.N.; Kladsuwan, L.; Soulard, L.; Swangmaneecharern, P.; Punpee, P.; Klomsa-ard, P.; Sriroth, K.; Keawsompong, S. The role of salicylic acid and benzothiadiazole in decreasing phytoplasma titer of sugarcane white leaf disease. Sci. Rep. 2021, 11, 15211. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, G.; Murolo, S.; Feliziani, E. Effects of an innovative strategy to contain grapevine bois noir: Field treatment with resistance inducers. Phytopathology 2013, 103, 785–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).