
Feeding Behavior of Riptortus pedestris (Fabricius) on Soybean: Electrical Penetration Graph Analysis and Histological Investigations



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Soybean Plants
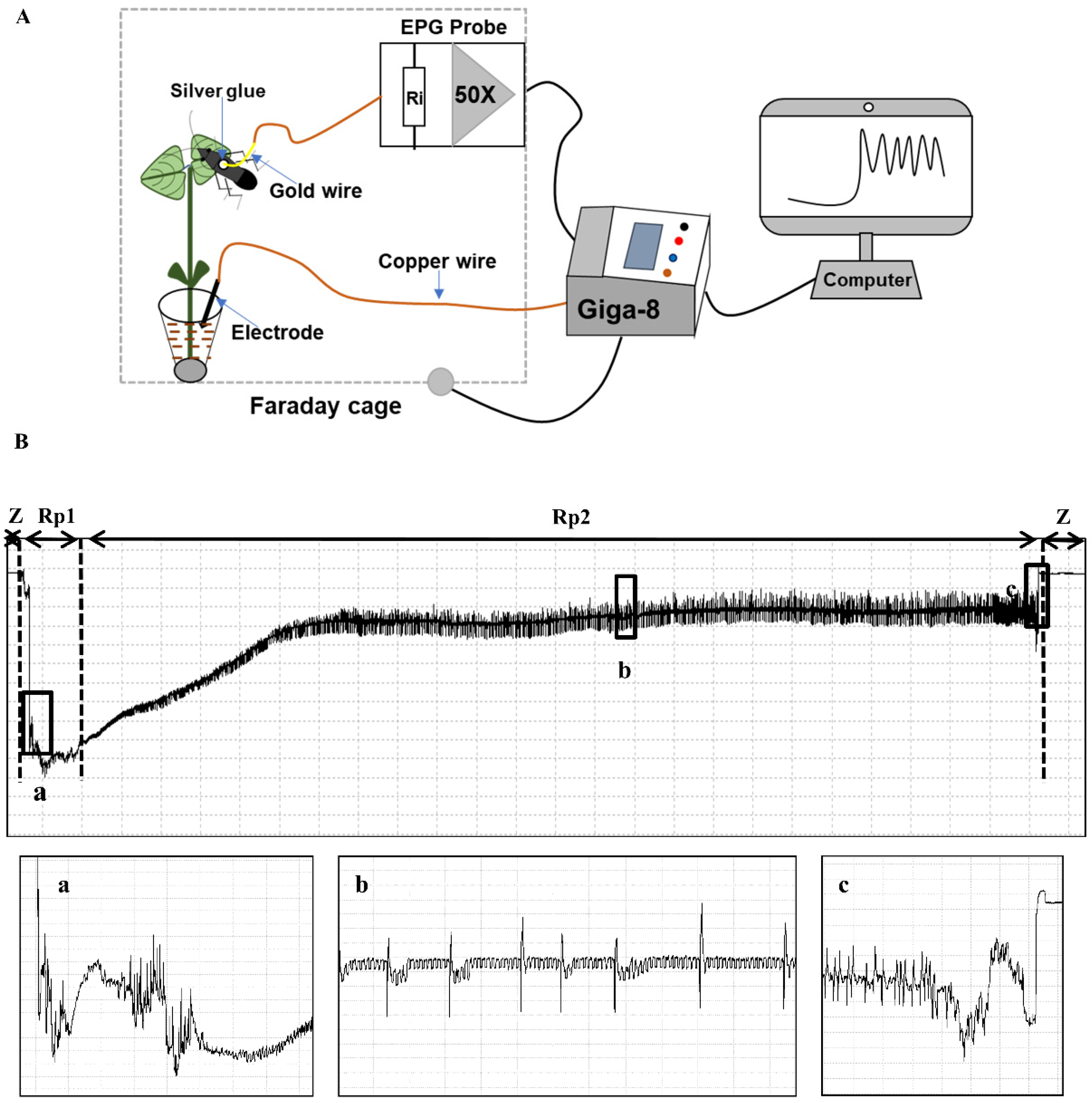
2.2. EPG Recordings
2.3. Plant Tissue Histology
3. Results
3.1. Characterization of EPG Waveforms of R. Pedestris on Soybean Plants
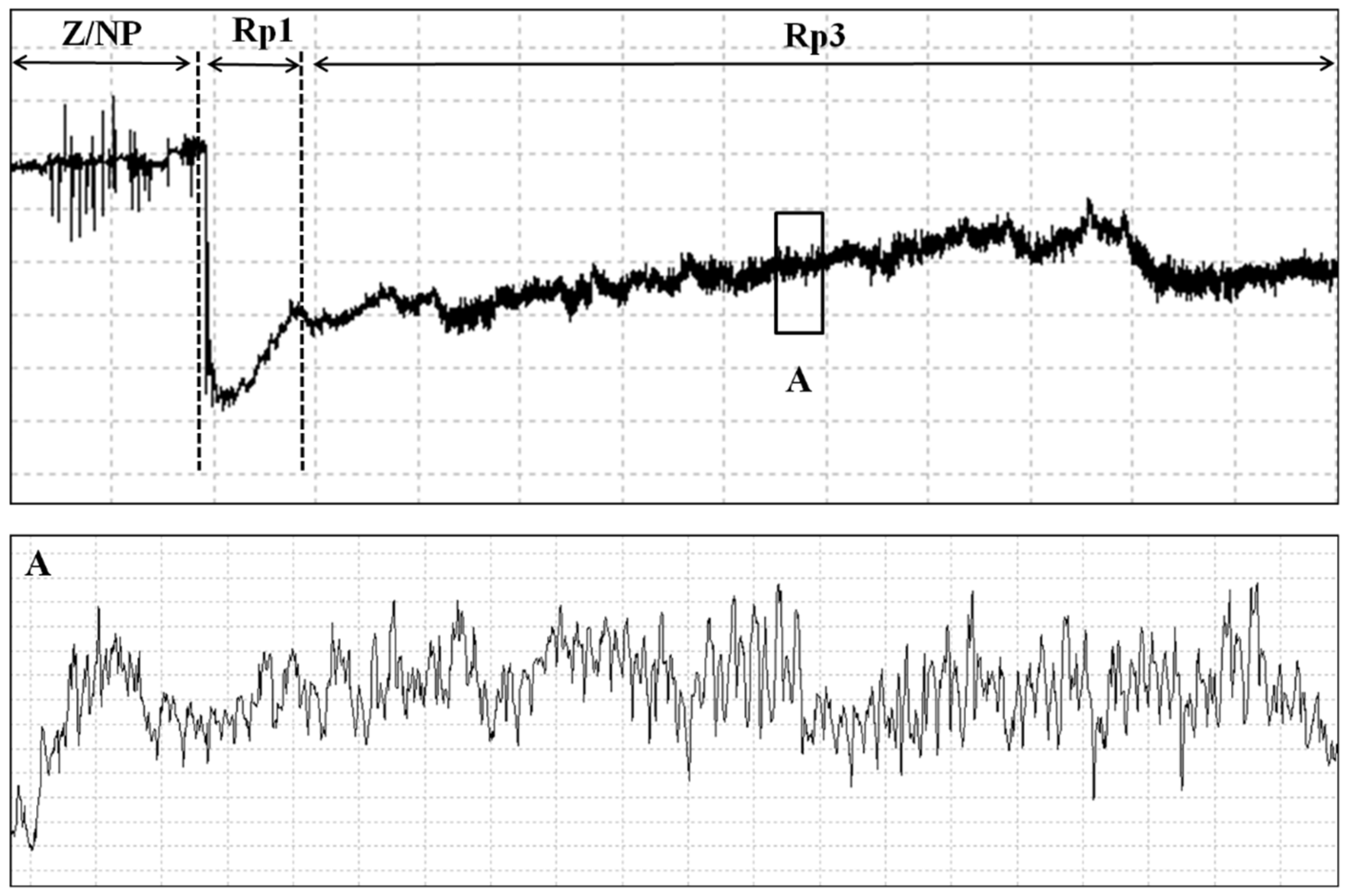
3.1.1. Noneprobing Waveforms (Z and Np)
3.1.2. Probing Waveforms: Pathway Phase–P Family (Rp1)
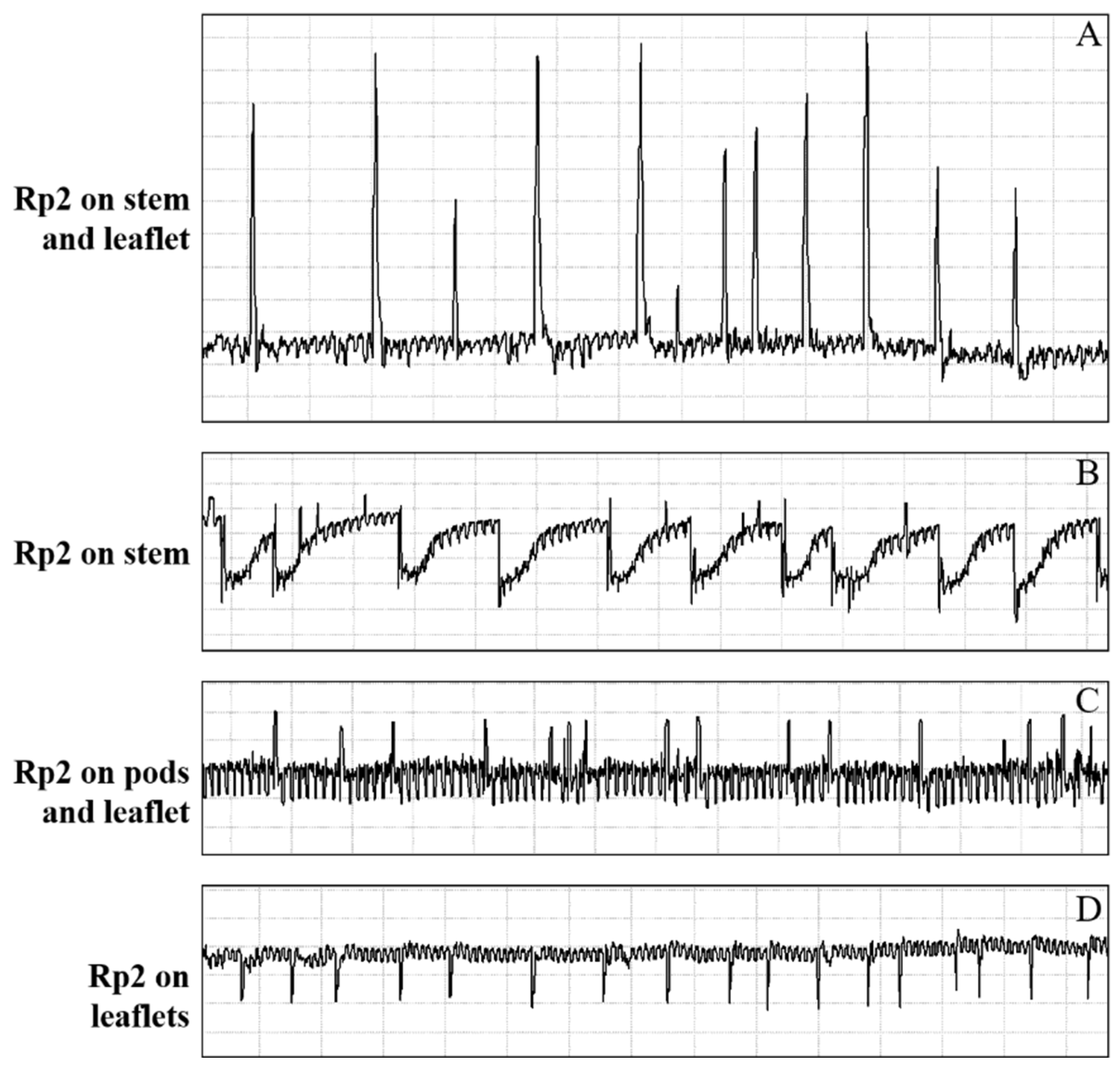
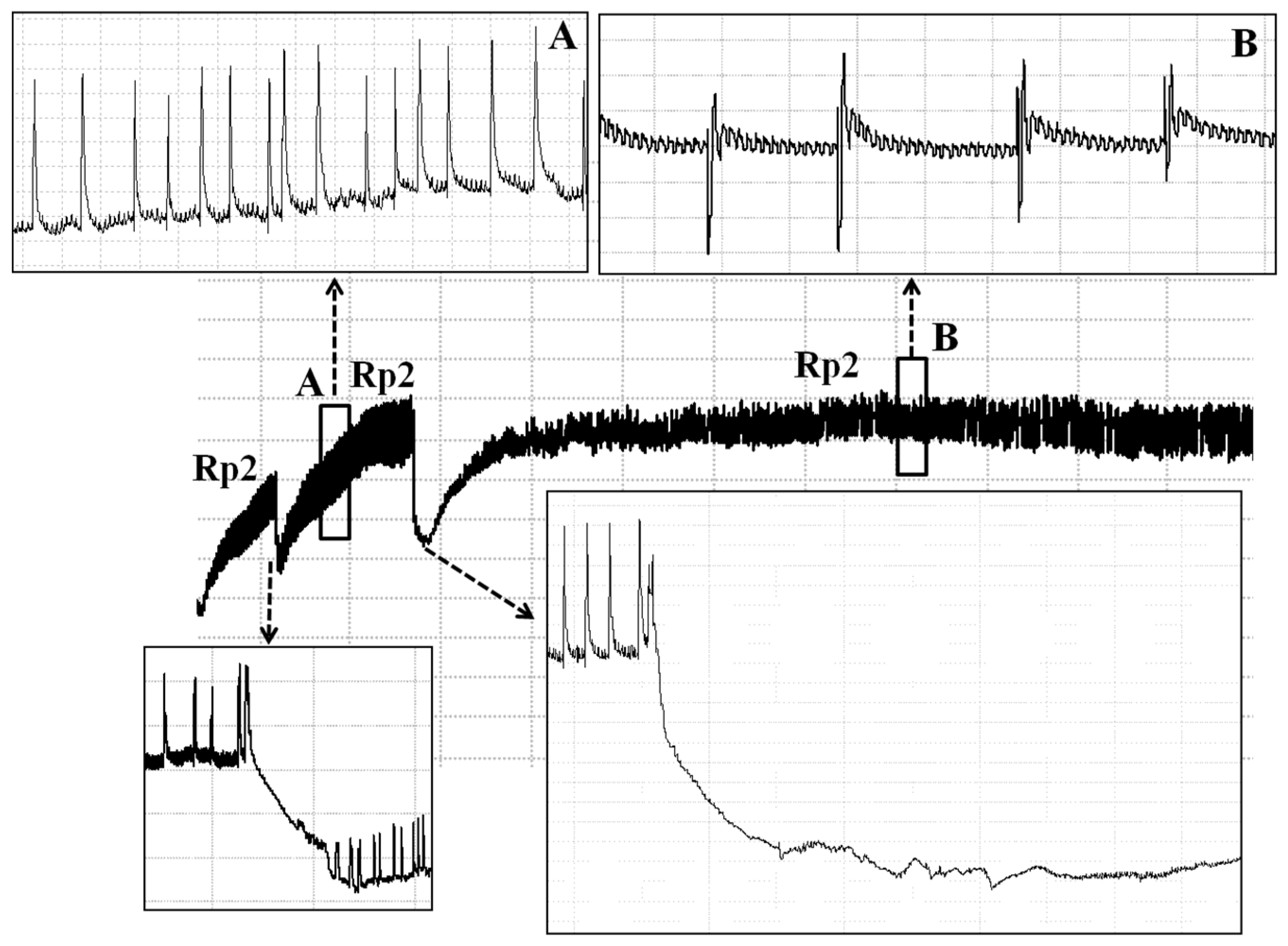
3.1.3. Xylem Sap Ingestion (Rp2)
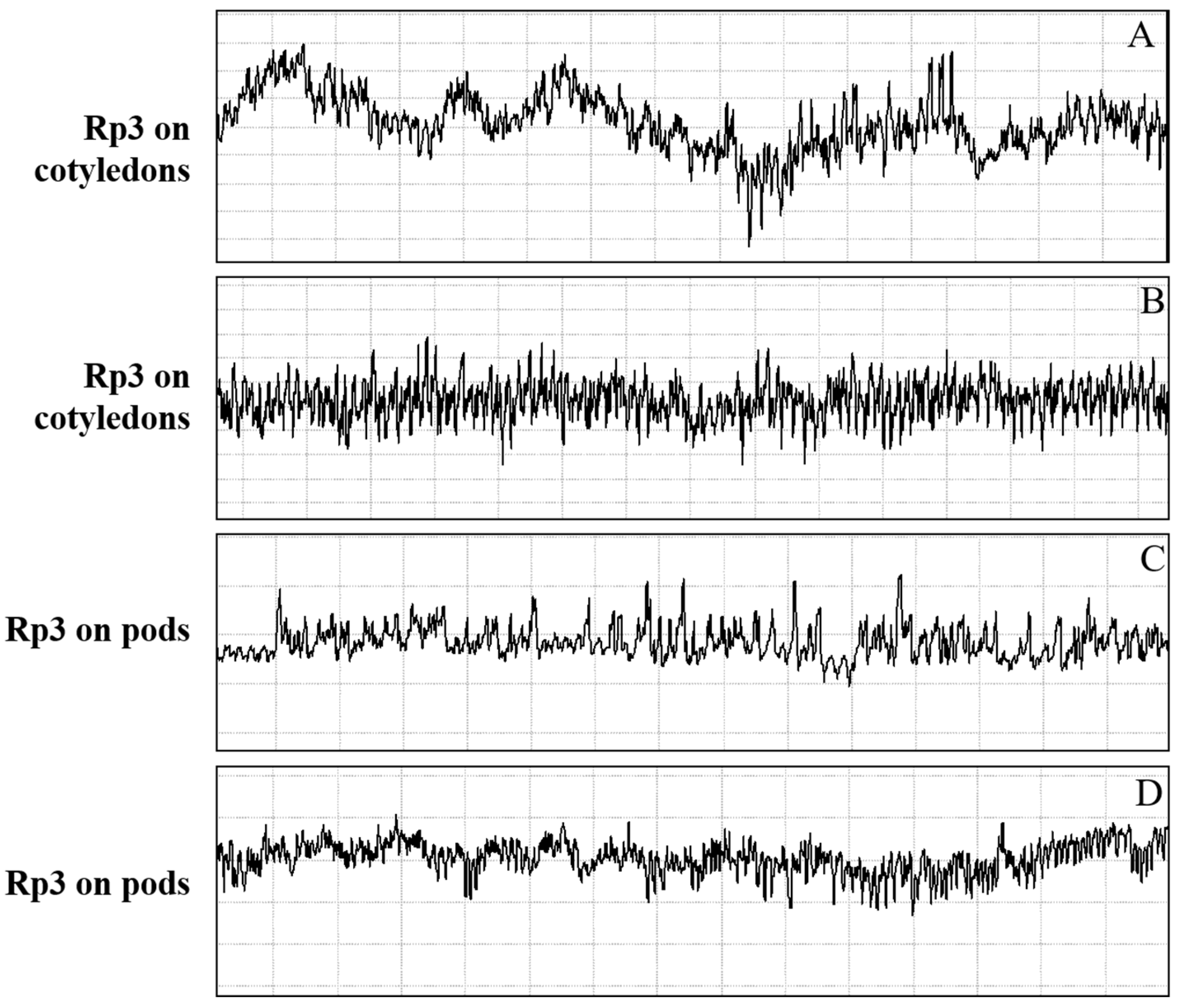
3.1.4. Salivation and Ingestion (Rp3)
3.1.5. Interruption Phase: Saliva Secretion (Rp4)
3.2. Correlations between Waveforms and Stylet Tips Position in Plant Tissues
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, M.; Lee, D.H. Characterization of Overwintering Behaviors and Sites of Bean Bug, Riptortus pedestris (Hemiptera: Alydidae), Under Laboratory and Field Conditions. Environ. Entomol. 2018, 47, 1280–1286. [Google Scholar] [CrossRef] [PubMed]
- Mainali, B.P.; Kim, H.J.; Yoon, Y.N.; Oh, I.S.; Bae, S.D. Evaluation of different leguminous seeds as food sources for the bean bug Riptortus pedestris. J. Asia-Pac. Entomol. 2014, 17, 115–117. [Google Scholar] [CrossRef]
- Lim, K. Fruits of apple and sweet persimmon are not essential food sources for Riptortus pedestris (Hemiptera: Alydidae) which causes fruit-spotting. J. Asia-Pac. Entomol. 2012, 15, 203–206. [Google Scholar]
- Mainali, B.P.; Lim, U.T. Annual pattern of occurrence of Riptortus pedestris (Hemiptera: Alydidae) and its egg parasitoids Ooencyrtus nezarae Ishii and Gryon japonicum (Ashmead) in Andong, Korea. Crop Prot. 2012, 36, 37–42. [Google Scholar] [CrossRef]
- Gao, Y.; Shi, S. The Relationship between Staygreen Syndrome in Soybean and Stink Bugs and Preventive Strategy. Soybean Sci. 2019, 38, 650–655. [Google Scholar]
- Li, K.; Zhang, X.X.; Guo, J.Q.; Penn, H.; Wu, T.T.; Li, L. Feeding of Riptortus pedestris on soybean plants, the primary cause of soybean staygreen syndrome in the Huang-Huai-Hai river basin. Crop J. 2019, 7, 360–367. [Google Scholar] [CrossRef]
- Yan, X.; An, J.; Gao, X. Relationship between population density of Riptortus pedestris and spring sowing soybean yiled using cumulative insect-days method. Plant Prot. 2021, 47, 96–100. [Google Scholar]
- Mahbubur, R.M.; Taek, L.U.; Lightfoot, D.A. Evaluation of mature soybean pods as a food source for two pod-sucking bugs, Riptortus pedestris (Hemiptera: Alydidae) and Halyomorpha halys (Hemiptera: Pentatomidae). PLoS ONE 2017, 12, e0176187. [Google Scholar]
- Hobbs, H.A.; Hill, C.B.; Grau, C.R.; Koval, N.C.; Wang, Y.; Pedersen, W.L.; Domier, L.L.; Hartman, G.L. Green Stem Disorder of Soybean. Plant Dis. 2006, 90, 513–518. [Google Scholar] [CrossRef][Green Version]
- Gao, Y.; Chen, J.; Shi, S. Research progress on soybean stink bug (Riptortus pedestris). Chin. J. Oil Crop Sci. 2019, 41, 804–815. [Google Scholar]
- Bae, D.S.; Kim, H.J.; Mainali, B.P. Infestation of Riptortus pedestris (Fabricius) decreases the nutritional quality and germination potential of soybean seeds. J. Asia-Pac. Entomol. 2014, 17, 477–481. [Google Scholar] [CrossRef]
- Kim, S.; Lim, U.T. New soybean variety, Agakong, as a host of Riptortus pedestris (Fabricius): Study on field occurrence and biological attributes in the laboratory. J. Asia-Pac. Entomol. 2010, 13, 261–265. [Google Scholar] [CrossRef]
- Backus, E.A.; Serrano, M.S.; Ranger, C.M. Mechanisms of hopperburn: An Overview of Insect Taxonomy, Behavior, and Physiology. Annu. Rev. Entomol. 2005, 50, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Lucini, T.; Panizzi, A.R. Electropenetrography (EPG): A Breakthrough Tool Unveiling Stink Bug (Pentatomidae) Feeding on Plants. Neotrop. Entomol. 2018, 47, 6–18. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Kanissery, R.; Ammar, E.-D.; Cabral, I.; Markle, L.T.; Patt, J.M.; Stelinski, L.L. Feeding Behavior of Asian Citrus Psyllid [Diaphorina citri (Hemiptera: Liviidae)] Nymphs and Adults on Common Weeds Occurring in Cultivated Citrus Described Using Electrical Penetration Graph Recordings. Insects 2020, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yang, J.; Liu, B.; Cui, H.; Zhang, Y.; Jiao, X. Feeding behavior explains the different effects of cabbage on MEAM1 and MED cryptic species of Bemisia tabaci. Insect Sci. 2019, 27, 1276–1284. [Google Scholar] [CrossRef]
- Dancewicz, K.; Gabryś, B.; Morkunas, I.; Samardakiewicz, S. Probing behavior of Adelges laricis Vallot (Hemiptera: Adelgidae) on Larix decidua Mill: Description and analysis of EPG waveforms. PLoS ONE 2021, 16, e0251663. [Google Scholar] [CrossRef]
- Almeida, A.C.D.S.; De Jesus, F.G.; Barrigossi, J.A.F. Unveiling the Feeding Behavior of Tibraca limbativentris (Hemiptera: Pentatomidae) on Rice Using an Electropenetrography Waveform Library. J. Insect Sci. 2020, 20. [Google Scholar] [CrossRef]
- Miles, P.W. The Saliva of Hemiptera. In Advances in Insect Physiology; Elsevier: London, UK, 1972; Volume 9, pp. 183–255. [Google Scholar] [CrossRef]
- Zhao, R.; He, Y.; Lu, Z.; Chen, W.; Zhou, C.; Wang, X.; Li, T. An analysis of the feeding behavior of three stages of Toxoptera citricida by DC electrical penetration graph waveforms. Entomol. Exp. Appl. 2019, 167, 370–376. [Google Scholar] [CrossRef]
- Bo, Y.S.; Kwon, Y.H.; Jin, K.J.; Kim, G.H. Electrical penetration graphic waveforms in relation to the actual positions of the stylet tips of Nilaparvata lugens in rice tissue. J. Asia-Pac. Entomol. 2009, 12, 89–95. [Google Scholar]
- Tuelher, E.S.; Backus, E.A.; Cervantes, F.; Oliveira, E.E. Quantifying Lygus lineolaris stylet probing behavior and associated damage to cotton leaf terminals. J. Pest Sci. 2020, 93, 663–677. [Google Scholar] [CrossRef]
- Rech, C.; Lucini, T.; Panizzi, A.R.; Nardi, C. Feeding Behavior of Collaria scenica (Stål) (Hemiptera: Miridae) on Wheat Plants: An EPG Waveform Characterization. Neotrop. Entomol. 2021, 50, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Lucini, T.; Panizzi, A.R. Electropenetrography Monitoring of the Neotropical Brown-Stink Bug (Hemiptera: Pentatomidae) on Soybean Pods: An Electrical Penetration Graph-Histology Analysis. J. Insect Sci. 2018, 18, 1–14. [Google Scholar] [CrossRef]
- Cook, C.A.; Neal, J.J. Feeding Behavior of Larvae of Anasa tristis (Heteroptera: Coreidae) on Pumpkin and Cucumber. Environ. Entomol. 1999, 2, 173–177. [Google Scholar] [CrossRef]
- Bonjour, E.L.; Fargo, W.S.; Webster, J.A.; Richardson, P.E.; Brusewitz, G.H. Probing Behavior Comparisons of Squash Bugs (Heteroptera: Coreidae) on Cucurbit Hosts. Environ. Entomol. 1991, 1, 143–149. [Google Scholar] [CrossRef]
- Shen, D.; Tang, Z.; Wang, C.; Wang, J.; Dong, Y.; Chen, Y.; Wei, Y.; Cheng, B.; Zhang, M.; Grenville-Briggs, L.J.; et al. Infection mechanisms and putative effector repertoire of the mosquito pathogenic oomycete Pythium guiyangense uncovered by genomic analysis. PLoS Genet. 2019, 15, e1008116. [Google Scholar] [CrossRef]
- Serteyn, L.; Ponnet, L.; Backus, E.A.; Francis, F. Characterization of electropenetrography waveforms for the invasive heteropteran pest, Halyomorpha halys, on Vicia faba leaves. Arthropod-Plant Interact. 2019, 14, 113–126. [Google Scholar] [CrossRef]
- Lucini, T.; Panizzi, A.R.; Backus, E.A. Characterization of an EPG Waveform Library for Redbanded Stink Bug, Piezodorus guildinii (Hemiptera: Pentatomidae), on Soybean Plants. Ann. Entomol. Soc. Am. 2016, 109, 198–210. [Google Scholar] [CrossRef]
- Lucini, T.; Panizzi, A.R. Electropenetrographic Comparison of Feeding Behavior of Dichelops furcatus (Hemiptera: Heteroptera: Pentatomidae) on Soybean and Spring Cereals. J. Econ. Entomol. 2020, 113, 1796–1803. [Google Scholar] [CrossRef]
- Lucini, T.; Panizzi, A.R. Feeding Behavior of the Stink Bug Dichelops melacanthus (Heteroptera: Pentatomidae) on Maize Seedlings: An EPG Analysis at Multiple Input Impedances and Histology Correlation. Ann. Entomol Soc. Am. 2017, 110, 160–171. [Google Scholar]
- Mitchell, P.L.; Cooke, S.B.; Smaniotto, L.F. Probing Behavior of Nezara viridula on Soybean: Characterization and Comparison of Electrical Penetration Graph (EPG) Waveforms on Vegetative and Reproductive Plant Structures. J. Agr. Urban Entomol. 2018, 34, 19–43. [Google Scholar] [CrossRef]
- Cervantes, F.A.; Backus, E.A.; Godfrey, L.; Akbar, W.; Clark, T.L. Characterization of an EPG Waveform Library for Adult Lygus lineolaris and Lygus hesperus (Hemiptera: Miridae) Feeding on Cotton Squares. Ann. Entomol. Soc. Am. 2016, 109, 684–697. [Google Scholar] [CrossRef]
- Tian, X.; Hu, Y.; Wang, Z.; Li, W.; Sun, Y.; Gao, Y. Effects of Soybean Organs in Vitro Feeding on the Growth and Development of Riptortus Pedestris Nymphs. Soybean Sci. 2021, 40, 398–402. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phase | Family | Type or Subtype | Plant Structure | Relative Amplitude (%) | Frequency (Hz) | Biological Meaning |
|---|---|---|---|---|---|---|
| Nonprobing | Z | Leaflet/stem/pod/cotyledon | Very low | Irregular | Stand still on plant surface | |
| NP | Leaflet/stem/pod/cotyledon | Low/high | Irregular | Various body movements (Walking, antennating or grooming) on the plant surface | ||
| Pathway | P | Rp1 | Leaflet/stem/pod/cotyledon | 83 (67–100) | Irregular | Stylet penetration with salivary sheath secretion |
| Ingestion | I | Rp2 | Leaflet/stem/pod | 45 (15–83) | 3.75 Hz (3.0–4.5) | Xylem sap ingestion |
| Salivation and ingestion | I | Rp3 | Pod/cotyledon | 70 (30–77) | Irregular | Watery saliva secretion, cell laceration and enzymatic maceration, and ingestion of lacerated/macerated cell tissues |
| Interruption | N | Rp4 | leaflet | Irregular | Brief interludes of salivation during Xylem sap ingestion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, Y.; Zhang, W.; Dong, Y.; Xia, A. Feeding Behavior of Riptortus pedestris (Fabricius) on Soybean: Electrical Penetration Graph Analysis and Histological Investigations. Insects 2022, 13, 511. https://doi.org/10.3390/insects13060511
Jin Y, Zhang W, Dong Y, Xia A. Feeding Behavior of Riptortus pedestris (Fabricius) on Soybean: Electrical Penetration Graph Analysis and Histological Investigations. Insects. 2022; 13(6):511. https://doi.org/10.3390/insects13060511
Chicago/Turabian StyleJin, Yan, Wendan Zhang, Yumei Dong, and Ai Xia. 2022. "Feeding Behavior of Riptortus pedestris (Fabricius) on Soybean: Electrical Penetration Graph Analysis and Histological Investigations" Insects 13, no. 6: 511. https://doi.org/10.3390/insects13060511
APA StyleJin, Y., Zhang, W., Dong, Y., & Xia, A. (2022). Feeding Behavior of Riptortus pedestris (Fabricius) on Soybean: Electrical Penetration Graph Analysis and Histological Investigations. Insects, 13(6), 511. https://doi.org/10.3390/insects13060511