Simple Summary
Japanese giant silkworm (JGS), Caligula japonica Moore, is an emerging defoliator pest of forest and fruit trees in East Asia, causing severe economic losses. In this study, we used eggs of the Chinese oak silkworm (COS), Antheraea pernyi Guérin-Méneville, as an alternative host to rear the dominant JGS egg parasitoid, Anastatus japonicus Ashmead. We showed that A. japonicus could be efficiently reared on COS eggs. Furthermore, A. japonicus reared from COS eggs had a high biological control potential against JGS. This study provides useful information on the demography, parasitism rate, and rearing methods of A. japonicus on COS eggs and the basis for developing a cost-effective biological control program against JGS using A. japonicus.
Abstract
Japanese giant silkworm (JGS), Caligula japonica Moore, is an emerging defoliator pest of forest and fruit trees in East Asia, causing severe economic losses. To develop a cost-effective biological control program against JGS, we used eggs of the Chinese oak silkworm (COS) Antheraea pernyi Guérin-Méneville as an alternative host to rear the most dominant JGS egg parasitoid Anastatus japonicus Ashmead. We compared the demographic parameters and total parasitism (killing) rates of A. japonicus parasitizing JGS and COS eggs using an age-stage, two-sex life table method. The results showed that A. japonicus performed differently on these two different hosts. Anastatus japonicus reared from COS eggs had a higher fecundity (369.7 eggs per female) and a longer oviposition period (35.9 days) on the COS than JGS eggs (180.9 eggs; 24.0 days). Consequently, A. japonicus parasitizing COS eggs had a higher intrinsic rate of increase (r = 0.1466 d−1), finite rate of increase (λ = 1.1579 d−1) and net reproductive rate (R0 = 284.9 offspring) than those parasitizing JGS eggs (r = 0.1419 d−1, λ = 1.1525 d−1, R0 = 150.0 offspring). The total net parasitism rate (the number of parasitized hosts in which the parasitoids successfully developed) of A. japonicus parasitizing COS eggs was 284.9, significantly higher than that of A. japonicus parasitizing JGS eggs (150.0), while the net non-effective parasitism rate (the number of parasitized hosts in which the parasitoids failed to develop) of the former (0.0) was significantly lower than that of the latter (9.6). These results suggest that A. japonicus can be efficiently reared on the alternative (or factitious) COS eggs, and the reared parasitoids have a high biological control potential against the target JGS.
1. Introduction
Native to East Asia, the Japanese giant silkworm (JGS) Caligula japonica Moore is widely distributed in China, Japan, North Korea, and The Russian Far East [1]. In China, 38 plant species belonging to 30 genera in 20 families have been reported as hosts of this pest; the main hosts are walnut (Juglans regia L.) [2,3], ginkgo (Ginkgo biloba L.) [4], and chestnut (Castanea mollissima Blume) [5]. Since it was first found unexpectedly in 2006 in Kangxian and Huixian Counties in Longnan City, Gansu Province, China, JGS has rapidly spread and outbroke in an area of 240 hectares, becoming an emerging pest in this region [5]. Current management strategies for this pest rely on repeated applications of chemical pesticides [6,7], which have led to resistance of this pest to some chemical pesticides [8]. Extensive use of insecticides also poses a serious threat to the environment, natural enemies, and ecological balance [9]. Therefore, it is crucial to develop effective and environmentally friendly alternatives for the sustainable management of this emerging pest. Biological control, especially by means of self-perpetuating and effective parasitoids, could be a viable option for the sustainable and eco-friendly management of insect pests [10,11].
Egg parasitoids have been used as major biological control agents for the control of some invasive agricultural and forestry pests. For example, Anastatus bifasciatus Geoffroy has been reported as a predominant egg parasitoid specie against the invasive brown marmorated stink bug Halyomorpha halys Stål in Europe [12,13,14,15]. Anastatus acherontiae Narayanan, Subba Rao and Ramachandra (later identified as Anastatus echidna Motschulsky), and Anastatus bangalorensis Mani and Kurian were two important parasitoids of the litchi stink bug Tessaratoma javanica Thunberg in India [16,17]. Anastatus fulloi Sheng and Wang was also a notable example of successful biological control of Tessaratoma papillosa Drury in litchi orchards in China [18,19]. Previously, Chen et al. [20] reported that A. japonicus was the most dominant JGS egg parasitoid during field surveys in Gansu Province, China. They found that this egg parasitoid was able to parasitize different ages of JGS eggs [20]. The parasitoid could also be successfully reared on eggs of the Chinese oak silkworm (COS) Antheraea pernyi Guérin-Méneville, and the COS egg seemed to be an ideal factitious host for mass rearing of this parasitoid because of its simple operation method, low production cost, and convenient transportation [21,22]. However, it is still unclear how A. japonicus reared from an alternative host (COS egg) would perform on the target host (JGS egg), i.e., the potential of mass-reared A. japonicus from this non-natural host for the biological control of JGS. Therefore, this study aimed to comparatively evaluate the reproductive performance of A. japonicus reared from COS eggs on these two different hosts.
A life table is an indispensable tool for population ecology and pest management to quantify population demographic parameters such as survival, development, longevity, and fecundity [23,24]. Compared to the traditional female age-specific life table, a two-sex life table method integrates both age-stage and two-sex components to accurately estimate important life-history traits such as age-stage or age-specific survival for both sexes and age-specific fecundity for females and generates important demographic parameters [23,24]. This approach is, therefore, a valuable tool for comparative analyses of a parasitoid’s life-history strategies under different conditions and has been widely used in the analysis of life table of insects such as Altica cyanea Weber, Trichogramma achaeae Nagaraja, and Nagarkatti, Aphelinus asychis Walker, and Encarsia formosa Gahan [25,26,27,28,29,30,31,32,33]. Here, we compared and quantified population demographic parameters and total parasitism rates of A. japonicus parasitizing COS and JGS eggs using the computer programs of TWOSEX-MSChart [34] and CONSUME-MSChart [35] based on the age-stage, two-sex life table method [23,24,36]. The study provides the basis for mass rearing of A. japonicus on COS eggs for an effective biological control program of JGS.
2. Materials and Methods
2.1. Host Insects
Studies were conducted at the Institute of Biological Control, Jinlin Agricultural University, China. Both the Japanese giant silkworm (JGS), C. japonica, and the Chinese oak silkworm (COS) A. pernyi were collected from fields. COS cocoons were collected during late fall every year from fields in Yongji City, Jilin Province, China, and held at 4 °C in a refrigerator (dark) until late February, when the cocoons were transferred to an emergence room (25 ± 1 °C, 60% ± 10% RH and natural photoperiod) for adult emergence. Unfertilized COS eggs were extracted from mature and virgin female moths by squeezing the abdomen of the moths, washed with distilled water, and then air-dried at room temperature prior to their use for the rearing or experiments [37].
JGS cocoons were collected in walnut orchards during the middle of June in Kangxian, Gansu Province, China. Approximately 300 cocoons were collected every year and placed in an insect cage (30 × 30 × 30 cm) in an open insectary so that they were exposed to similar field conditions and expected to emerge as field populations did from August to October. Newly emerged adults were paired and kept in insect rearing cages (50 × 50 × 50 cm). The adults were monitored daily for oviposition, and freshly laid eggs (<2 days) were collected for the experiments.
2.2. Parasitoid
A laboratory colony of A. japonicus was established with adult wasps that emerged from field-collected and parasitized JGS egg masses in walnut orchards in 2017 in Kangxian, Gansu Province (105–106° E, 32.9–33.7° N), China [1,38]. The parasitoid species was identified based on the morphological descriptions by Dr. Gary Gibson (Agriculture and Agri-Food Canada, Canadian National Collection of Insects) and was further confirmed by COI sequence (GenBank accession number: MK604240) [1,39]. The parasitoid was maintained on COS eggs (unfertilized, extracted and washed eggs, see Section 2.1) under controlled conditions (25 ± 1 °C, 70% ± 5% RH, and 14 L:10 D photoperiod) [37] and had been reared for six generations on COS eggs before they were used in this study. Briefly, newly emerged wasps (<6 h) were collected into cylindrical and transparent plastic containers (14.0 × 9.0 cm, height × diameter) covered with fine mesh, with 30% honey water provided as food. Ten pairs of 3-day-old wasps were provided with ≈300 eggs in each container. After a 24 h exposure, the wasps were removed, and the exposed hosts were monitored until the emergence of wasps.
2.3. Performance of A. japonicus Reared from COS Eggs on COS and JGS Eggs
All experiments were conducted under laboratory conditions (25 ± 1 °C, 70% ± 5% RH, and 14 L:10 D photoperiod) and used the parasitoids reared from COS eggs as described above. To determine the age-specific survival and fecundity of A. japonicus on COS eggs, a pair of newly emerged (<6 h) female and male wasps was first mated in a glass tube (12 × 3 cm, length × diameter) and then provided with 40 COS eggs (unfertilized, extracted, and washed eggs, see Section 2.1) and 30% honey water as food in the tube. The host eggs were glued on a strip egg card (6 × 1.5 cm, length × width). New COS eggs and honey water were replaced daily until the death of the female wasp. If the male wasp died before the female, it was replaced with another male. For each replicate, the exposed egg card from the previous day was transferred to a new glass tube (7 × 2 cm, length × diameter). The sex and date of each emerged wasp were recorded daily until the wasp emergence had ceased for 20 consecutive days. Finally, all host eggs without emergence holes were dissected to determine the presence or absence of recognizable parasitoid cadavers under a stereomicroscope (Leica DFC 450, M165C, Singapore). There were 20 valid replicates (5 replicates were excluded because the female wasps either escaped during the replacement of fresh egg cards or were accidentally killed). Male and female A. japonicus adults that emerged on the same day were paired and used to initiate the cohort in this study, which might not reflect the natural emergence pattern of the parasitoid population. Therefore, a bootstrap-match technique based on the life table method as described by Amir-Maafi et al. was used in this study [40].
Similarly, for the test of JGS eggs, a pair of newly emerged (<6 h) wasps reared from COS eggs was mated in a glass tube (12 × 3 cm, length × diameter), with 40 newly JGS eggs and 30% honey water provided in the tube. New JGS eggs and honey water were also replaced daily, and the survival of both female and male wasps was monitored until the death of the female. Any male who died before the female was also replaced with another male. There were 24 valid replicates (1 replicate was excluded because the female wasp escaped during the replacement of fresh egg cards). The rest of the experimental procedures were similar to those described above for the test of COS eggs.
2.4. Data Analysis
Data were analyzed according to the two-sex life table method [23,24,36]. The lifetime longevity, fecundity (i.e., the number of A. japonicus offspring successfully emerged from parasitized hosts), and non-effective (reproductive) parasitism rate (i.e., the number of A. japonicus offspring failed to develop or emerge from parasitized hosts) and other life table parameters were estimated for each population parasitizing COS or JGS eggs.
The age-stage survival rate (sxj) describes the probability that a newly laid egg will survive to age x and developmental stage j [23] and is calculated as:
where n01 is the number of newborns at the beginning of the population and nxj is the number of individuals among n01 surviving to age x and stage j. Variation in developmental rates among individuals may result in an overlap of survival at different stages.
The age-specific survival rate (lx) represents the survival rate from egg to age x, which ignores the stage differentiation and is the sum of sxj of all stages at age x, while the age-specific fecundity (mx) represents the average number of eggs laid for the individuals survived to age x. The lx and mx are calculated as follows:
where β is the number of stages and fxj refers to the effective parasitism rate (i.e., the number of parasitoids successfully emerged from parasitized hosts) [41]. The age-specific net reproductive rate of population (lxmx) is the product of lx multiplying mx, which represents the average number of successfully parasitized eggs by all surviving individuals in the population at age x.
The life expectancy (exj) represents the remaining survival time of individuals at age x and stage j and can be calculated according to Chi and Su [42] as:
where is the probability that an individual of nxj will survive to age i and stage y.
The net reproductive rate (R0) defines the average number of emerged offspring by a parasitoid during its lifetime and is calculated as:
The intrinsic rate of increase (r) refers to the average daily growth rate of the population when it reaches the stable age-stage distribution, which is calculated by the iterative bisection method and the Euler-Lotka equation with the age indexed from 0 [43]:
The finite rate of increase (λ) refers to the average daily growth rate of the population when it reaches the stable age-stage distribution and is calculated as follows:
The mean generation time (T) refers to the time required to achieve R0 when the population reaches a steady growth rate (λ and r) and is calculated as:
The age-stage reproductive value (vxj) refers to the average contribution of individuals at age x and stage j to future population growth [44,45] and is calculated as:
The net non-effective parasitism rate G0 refers to the total number of host eggs that are killed due to parasitism without the successful emergence of parasitoid adults. It is calculated as:
The age-stage total parasitism rate pxj is the sum of the effective parasitism rate (fxj) and the non-effective parasitism rate (gxj). Therefore, the net parasitism rate P0 is the sum of the net reproductive rate (R0) and is calculated as:
The transformation rate (Qp) is defined as the total number of hosts killed to produce a single parasitoid offspring and is calculated as:
The TWOSEX-MSChart program was used to calculate these demographic parameters [34], while the CONSUME-MSChart computer program was used to analyze the non-effective parasitism rate [35]. The bootstrap technique with 100,000 resamplings was used to estimate the standard errors of these population parameters [34]. Paired bootstrap test in the TWOSEX-MSChart program was used to test the significance of differences in the parameters (p ≤ 0.05) [46]. All figures were created using Sigma Plot 14.0.
3. Results
3.1. Longevity and Lifetime Fecundity of A. japonicus Parasitizing COS and JGS Eggs
As shown in Table 1, the initial cohort sizes of A. japonicus were 20:20 (female:male) on COS eggs and 24:24 (female:male) on JGS eggs, and the matched cohort sizes of the parasitoid were 259:77 (female:male) on COS eggs and 175:36 (female:male) on JGS eggs. The female adult longevity, female total longevity, male adult longevity, and male total longevity of A. japonicus parasitizing COS eggs were significantly longer than those parasitizing JGS eggs. However, no significant differences were observed in the adult preoviposition period and total preoviposition period between the parasitoids parasitizing COS and JGS eggs. The oviposition period and number of eggs laid per oviposition day by A. japonicus parasitizing COS eggs were higher than those parasitizing JGS eggs. The lifetime fecundity of the parasitoid was higher on the COS than JGS eggs. The age at lx ≤ 0.5 by the parasitoid parasitizing COS eggs was significantly delayed when compared to those parasitizing JGS eggs.

Table 1.
Population cohort, adult longevity, total longevity, adult preoviposition period, total preoviposition period, oviposition days, eggs laid per oviposition day, fecundity, and age at 0.5 lx of A. japonicus parasitizing COS and JGS eggs.
3.2. Age-Stage and Age-Specific Survival Rate and Fecundity of A. japonicus Parasitizing COS and JGS Eggs
The age-stage survival rate (sxj) revealed significant differences between A. japonicus parasitizing COS and JGS eggs (Figure 1). Adult females parasitizing COS eggs survived 79 days, which was longer than those parasitizing JGS eggs (52 days). However, the peak of the sxj at the adult stage of adult female parasitoids was higher on JGS eggs (82.94%) at the age of 37 days than on COS eggs (76.49%) at the age of 33 or 34 days. On the COS eggs, the sxj started to decrease at the age of 34 days but remained relatively stable from 45 to 52 days of age. On the JGS eggs, however, the sxj remained stable from 32 to 37 days of age and then decreased sharply after the age of 37 days. The peak of the age-stage survival rate of adult male A. japonica was also higher on the COS (22.92%) than JGS eggs (16.11%) at the age of 25 days. Overall, adult females had much longer longevity than adult males (Figure 1).
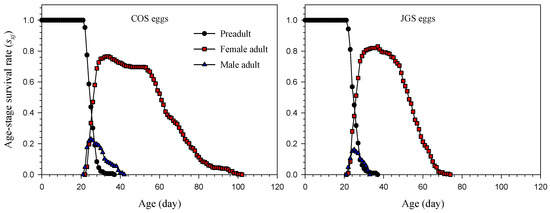
Figure 1.
Age-stage survival rate (sxj) of Anastatus japonicus parasitizing COS and JGS eggs.
The age-specific survival rate (lx), age-specific fecundity (fx2), age-specific fecundity of population (mx), and age-specific net reproductive rate (lxmx) of A. japonicus parasitizing COS eggs and JGS eggs were illustrated on Figure 2. The lx was higher on the JGS than COS eggs from 36 to 47 days of age. The highest mx was 12.80 at the age of 42 days on the COS eggs or 10.54 at the age of 35 days on the JGS eggs. Both the mx and lxmx decreased gradually on COS and JGS eggs (after 42 and 35 days old, respectively), except that the mx increased again from 79 to 82 days of age and then decreased again on the COS eggs. The ages at which adult females reached 90% cumulative fecundity on COS eggs (256.45) and JGS eggs (135.03) were 61 and 47 days, respectively.
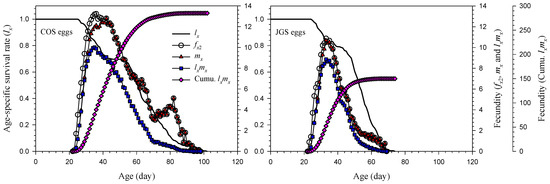
Figure 2.
Age-specific survival rate (lx), age-specific fecundity (fx2), age-specific fecundity of population (mx), age-specific net reproductive rate of population (lxmx), and cumulative fecundity of Anastatus japonicus parasitizing COS and JGS eggs.
3.3. Life Expectancy and Reproductive Value of A. japonicus Parasitizing COS and JGS Eggs
The life expectancy values (exj) of a newly laid A. japonicus egg on COS eggs and JGS eggs were 58.99 and 51.09 days, respectively (Figure 3). The highest exj for the adult females on COS eggs (43.63 days) was higher than that on JGS eggs (33.49 days) at 23 and 22 days of age, respectively. The highest exj for the adult males on COS eggs (11.27 days) was also higher than that on JGS eggs (7.66 days) at the age of 22 days.
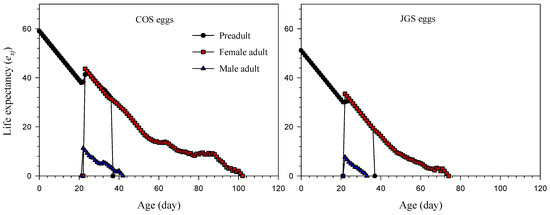
Figure 3.
Life expectancy (exj) of Anastatus japonicus parasitizing COS and JGS eggs.
The reproductive value of A. japonicus (vxj) first gradually increased with age; reaching the peak at the ages of 32 days (86.69 d−1) and 30 days (65.39 d−1) when parasitizing COS eggs and JGS eggs, respectively, and then gradually decreased with age (Figure 4). The vxj of adult females dramatically increased when they started oviposition (Figure 4). The reproductive value at the age of zero (v01) was equal to the finite rate of increase (λ). The reproductive values of newly laid eggs of A. japonicus parasitizing COS eggs and JGS eggs were 1.16 and 1.15, respectively.
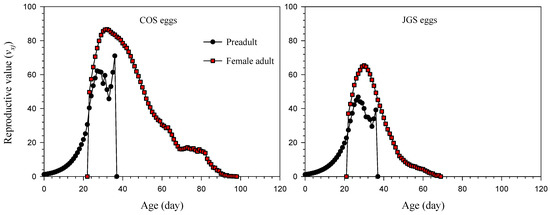
Figure 4.
Reproductive values (vxj) of Anastatus japonicus parasitizing COS and JGS eggs.
3.4. Population Parameters of A. japonicus Parasitizing COS and JGS Eggs
The population demographic parameters of A. japonicus parasitizing COS and JGS eggs are shown in Table 2. The net reproduction rate (R0), intrinsic rate of increase (r), finite rate of increase (λ), and mean generation time (T) of A. japonicus parasitizing COS eggs were all significantly higher than those parasitizing JGS eggs. There were significant differences in the numbers of female and male offspring produced between parasitoids parasitizing COS eggs and JGS eggs. The net non-reproductive parasitism rate (G0) on COS eggs was zero. The total net parasitism rate (P0) (that was the sum of female and male offspring and the net non-reproductive parasitism) was higher on the COS than JGS eggs. The transformation rate (Qp) was significantly lower on the COS than JGS eggs.
Table 2.
Population parameters of Anastatus japonicus parasitizing COS and JGS eggs.
4. Discussion
In this study, we compared the demographic parameters and parasitism rates of A. japonicus parasitizing COS and JGS eggs. We demonstrated that the parasitoid could be efficiently reared on the alternative (or factitious) host COS eggs, and reared parasitoids from the COS eggs had a high biological control potential against the target host JGS.
The two-sex life table analysis allows a thorough understanding of the demography and parasitic effectiveness of A. japonicus and comparisons of some important life-history traits and key demographic parameters of the parasitoid parasitizing these two different hosts. Overall, we showed that A. japonicus was more effective on COS than JGS eggs. Most of these key demographic parameters, such as adult longevity, female ovipositional period, lifetime fecundity, and net parasitism rate, were higher when parasitizing COS eggs than JGS eggs. This indicates that the parasitoid has a higher per capita reproductive potential on the COS than JGS eggs. Previously, the COS egg was also shown to be a suitable factious host for rearing other egg parasitoids such as T. dendrolimi Matsumura, T. chilonis Ishii [11], and Aprostocetus brevipedicellus Yang and Cao [47]. Here, we further confirm that A. japonicus can be efficiently reared on COS eggs.
The different demographics and parasitic effectiveness of A. japonicus on COS and JGS eggs may be caused by several factors. First, both host quality and host egg surface characteristics (e.g., host egg size and age, eggshell shape, and chorion thickness) could affect host acceptance and the resultant parasitism by an egg parasitoid [20,48,49,50,51]. In this study, we used naturally laid JGS eggs, which have a hard surface and thick eggshell [20]. In contrast, the COS eggs we used were extracted from virgin female moths, washed to remove the outer layer surface substrates, and the eggshell was still soft [37]. It could be harder to penetrate a naturally laid JGS egg than a treated COS egg by the ovipositor of female A. japonicus. There may involve higher costs for A. japonicus to parasitize JGS than COS eggs in terms of host handling time and energy investment per oviposition, thereby reducing the parasitoid’s longevity and lifetime fecundity when parasitizing the JGS eggs [50,52]. Indeed, previous studies found that extracted and washed COS eggs from virgin female moths largely facilitated ovipositor penetration by several egg parasitoids [38]. These studies also indicated that the hardness of the host eggshell negatively reduced a female parasitoid’s oviposition rate and longevity [50]. For example, the fecundity of Trichogramma confusum Viggiani on the eggs of Corcyra cephalonica Stainton (eggshell thickness of 4.2 um) and COS eggs (eggshell thickness of 48.0 µm) was 147 and 47, respectively [53]. Second, natal host species could affect a parasitoid’s performance on different host species. Since A. japonicus had been reared for six generations on COS eggs in the laboratory and all tested wasps were reared from COS eggs, it is likely that the parasitoid might have adapted to this natal host. Many studies showed that many parasitoids performed better in or on the host they previously developed due to preimaginal learning [54,55,56,57]. For example, the parasitism rate and longevity of Aphidius microlophii Pennacchio and Tremblay were significantly increased after the parasitoid was reared on the non-natural host of the pea aphid Acyrthosiphon pisum Harris for several generations [54].
Regardless of the host species, the overall patterns of these life-history traits (age-stage or age-specific survival rate, age-specific fecundity, life expectancy, reproductive values, Figure 1, Figure 2, Figure 3 and Figure 4) of A. japonicus were similar when parasitizing COS and JGS eggs; the differences were the peak values of these curves that were higher on the COS than JGS eggs. One only exception was that the sxj peak of female A. japonicus was slightly higher on the JGS than COS eggs (Figure 1). We also noticed that although a JGS egg (egg length ≈ 2.40 mm, width ≈ 1.49 mm) was smaller than a COS egg (egg length ≈ 3.21 mm, width ≈ 2.82 mm), the percentage of female A. japonicus offspring on the JGS eggs (79.08%) was slightly higher than that on the COS eggs (72.03%). The sxj refers to the probability of an egg surviving to age x and stage j and could be affected by host quality (i.e., host suitability). Offspring sex allocation decisions could also be affected by host quality, among various other factors (e.g., female age, host species, host density, female wasp density, superparasitism, and environmental conditions) [57]. A previous study reported that female A. japonicus adults were able to detect the size and estimate the quality of a host egg through receptors located on the ovipositor so that they were capable of adjusting sex allocation decisions based on host quality [58]. In general, arrhenotokous parasitoids allocate more female offspring to high-quality hosts and more male offspring to low-quality hosts [57]. Similar findings were reported for A. japonicus on six different host species; there was a significantly positive correlation between the size of the host egg and the percentage of female offspring of the parasitoid [59]. These results suggest that the natural host is more suitable than the alternative host for offspring survival, and JGS eggs may have a higher quality than COS eggs. In addition, the COS eggs used in this study were not fertilized. Several studies corroborated that egg parasitoids preferred fertilized over unfertilized eggs [58,60]. For example, Krugner found that the egg parasitoids Gonatocerus morrilli Howard and Telenomus coloradensis Crawford preferred to parasitize fertilized than unfertilized eggs of Homalodisca vitripennis Germar [61]. Although a high survival rate and a high percentage of female offspring are favorable for effective mass rearing for introduction and augmentative releases of parasitoids, these differences are generally marginal for A. japonicus parasitizing COS and JGS eggs. It is worth of mentioning that the cumulative fecundity (lxmx) reached 90% at the ages of 38 and 24 days when parasitizing COS and JGS eggs, respectively (Figure 2). This suggests that female adults older than 38 days are no longer reproductive and should not be used for the rearing of this parasitoid on COS eggs under similar conditions as the current study. Such information could help improve mass-rearing techniques and laboratory bioassays or field release when selecting the suitable age of females for rearing or studies.
In conclusion, A. japonicus has a high biological control potential against JGS as it exhibits a long longevity and ovipositional period as well as a high fecundity on JGS eggs. As demonstrated in this study, A. japonicus reared from COS eggs are callable of killing about 160 JGS eggs per female lifetime under the tested conditions. Effective methods for the rearing of A. japonicus and other related egg parasitoids on COS eggs have been investigated previously. The current study further demonstrated that A. japonicus reared from COS eggs had a high killing capacity on the target JGS. Our results thus have important implications for mass rearing of A. japonicus on this factitious host for augmentative release against JGS. However, further studies may be needed to evaluate the efficiency of this parasitoid in the field and environmental conditions (e.g., temperature and humidity) that may affect its efficiency in the field.
Author Contributions
Conceptualization, L.-S.Z.; methodology, X.-Y.W., Y.-M.C. and L.-S.Z.; validation, L.-S.Z. and X.W.; formal analysis, L.-S.Z., Y.-M.C. and X.W.; investigation, X.-Y.W. and Y.-M.C.; resources, L.-S.Z. and R.-E.L.; data curation, X.-Y.W., Y.-M.C. and L.-S.Z.; writing—original draft preparation, X.-Y.W., Y.-M.C. and L.-S.Z.; writing—review and editing, L.-S.Z. and X.W.; visualization, L.-S.Z. and X.W.; supervision, L.-S.Z. and X.W.; project administration, L.-S.Z.; funding acquisition, L.-S.Z. and R.-E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key R&D Program of China (2017YFD0201000) and Key R&D Programs (Agriculture) of Gansu Province (20YF3NK030).
Data Availability Statement
Not applicable.
Acknowledgments
We appreciate Gary Gibson (Honorary Research Associate, Agriculture and Agri-Food Canada, Canadian National Collection of Insects) for the species confirmation of Anastatus japonicus and Hsin Chi (National Chung Hsing University) for reviewing and editing this paper before submission.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, Y.M.; Gibson, G.A.P.; Peng, L.F.; Iqbal, A.; Zang, L.S. Anastatus Motschulsky (Hymenoptera, Eupelmidae): Egg parasitoids of Caligula japonica Moore (Lepidoptera, Saturniidae) in China. ZooKeys 2019, 881, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.H.; Ge, W.G.; Wang, Y.F. Occurrence regularity and control measures of Ginkgo biloba. Jilin For. Sci. Technol. 1997, 6, 19–20. [Google Scholar]
- Huang, Y.Z. Prevention and control of walnut tree by Ginkgo biloba. Gansu Sci. Technol. 2014, 30, 161–162. [Google Scholar]
- Jiang, D.A. Study on the occurrence and control of Dictypoloca japonica. For. Sci. Technol. 2003, 1, 25–27. [Google Scholar]
- Qiao, X.; Wang, Y.C.; Wu, G.; Wang, S.C.; Hu, J.Z.; Liu, T.X.; Feng, S.Q. Occurrence reasons and control measures of Caligula Japonica in Longnan City of China. Plant Dis. Pests 2014, 5, 38–41. [Google Scholar] [CrossRef]
- Wu, N.M.; Yang, S.Z.; Chen, J.; Du, Y.B.; Cai, J.X.; Fu, C.S. A study on the Anastatus spp. released in forest for control of Dictyoploca japonica Moore. Nat. Enemies Insects 2001, 23, 26–29. [Google Scholar]
- Dong, D.J. Toxicity determination and control effect of several pesticides against Dictyoploca japonica Butler. For. Pest Dis. 2013, 32, 38–40. [Google Scholar]
- Wang, Y.X.; Qiao, X.; Wang, Z.K. Pharmacodynamic study on different insecticide for Caligula japonica Moore. Mod. Agric. Sci. Technol. 2012, 14, 95–96. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Huang, N.X.; Jaworski, C.C.; Desneux, N.; Zhang, F.; Yang, P.Y.; Wang, S. Long-term, large-scale releases of Trichogramma promotes pesticide decrease in maize in Jilin province, Northern China. Entomol. Gen. 2020, 40, 331–335. [Google Scholar] [CrossRef]
- Zang, L.S.; Wang, S.; Zhang, F.; Desneux, N. Biological control with Trichogramma in China: History, present status and perspectives. Annu. Rev. Entomol. 2021, 66, 463–484. [Google Scholar] [CrossRef]
- Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the distribution of exotic egg parasitoids of Halyomorpha halys in Europe with a large-scale monitoring program. Insects 2021, 12, 316. [Google Scholar] [CrossRef] [PubMed]
- Stahl, J.M.; Babendreier, D.; Marazzi, C.; Caruso, S.; Costi, E.; Maistrello, L.; Haye, T. Can Anastatus bifasciatus be used for augmentative biological control of the brown marmorated stink bug in fruit orchards? Insects 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Stahl, J.M.; Babendreier, D.; Haye, T. Using the egg parasitoid Anastatus bifasciatus against the invasive brown marmorated stink bug in Europe: Can non-target effects be ruled out? J. Pest Sci. 2018, 91, 1005–1017. [Google Scholar] [CrossRef] [PubMed]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef][Green Version]
- Lalitha, Y.; Ballal, C.R.; Gupta, A. Interaction between Anastatus acherontiae and Anastatus bangalorensis (Hymenoptera: Chalcidoidea) two potential parasitoids of Litchi stink bug Tessaratoma javanica Thunberg. In Conference on National Priorities in Plant Health; SV Agricultural College: Tirupati, India, 2016; pp. 4–5. [Google Scholar]
- Choudhary, J.S.; Prabhakar, C.S.; Moanaro; Das, B.; Kumar, S. Litchi stink bug (Tessaratoma javanica) outbreak in Jharkhand, India, on litchi. Phytoparasitica 2013, 41, 73–77. [Google Scholar] [CrossRef]
- Li, D.S.; Liao, C.; Zhang, B.X.; Song, Z.W. Biological control of insect pests in litchi orchards in China. Biol. Control 2014, 68, 23–36. [Google Scholar] [CrossRef]
- Han, S.C.; Liu, W.H.; Chen, Q.X.; Zeng, B.K.; Chen, N.R.; Lin, J.X.; Yu, F.X. Mass releasing Anastatus japonicus to control Tessaratoma papillosa in Hong Kong. J. Biol. Control 1999, 15, 54–56. [Google Scholar]
- Chen, Y.M.; Qu, X.R.; Li, T.H.; Iqbal, A.; Wang, X.G.; Ren, Z.Y.; Desneux, N.; Zang, L.S. Performances of six eupelmid egg parasitoids from China on Japanese giant silkworm Caligula japonica with different host age regimes. J. Pest Sci. 2020, 94, 309–319. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhang, X.; Zang, L.S.; Du, W.M.; Hou, Y.Y.; Ruan, C.C.; Desneux, N. Advantages of diapause in Trichogramma dendrolimi mass production via eggs of the Chinese silkworm, Antheraea pernyi. Pest Manag. Sci. 2018, 74, 959–965. [Google Scholar] [CrossRef]
- Li, T.H.; Tian, C.Y.; Zang, L.S.; Hou, Y.Y.; Ruan, C.C.; Yang, X.B.; Monticelli, L.; Desneux, N. Multiparasitism with Trichogramma dendrolimi on egg of Chinese oak silkworm, Antheraea pernyi, enhances emergence of Trichogramma ostriniae. J. Pest Sci. 2019, 92, 707–713. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Mitra, S.; Mobarak, S.H.; Barik, A. Age-stage, two-sex life table of the biocontrol agent, Altica cyanea on three Ludwigia species. Biologia 2021, 76, 101–112. [Google Scholar] [CrossRef]
- del Pino, M.; Gallego, J.R.; Suárez, E.H.; Cabello, T. Effect of temperature on life history and parasitization behavior of Trichogramma achaeae Nagaraja and Nagarkatti (Hym.: Trichogrammatidae). Insects 2020, 11, 482. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, C.L.; Yang, X.B.; Chi, H.; Dai, P.; Desneux, N.; Benelli, G.; Zang, L.S. Yacon as an alternative host plant for Encarsia formosa mass-rearing: Validating a multinomial theorem for bootstrap technique in life table research. Pest Manag. Sci. 2021, 77, 2324–2336. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2020, 28, 602–610. [Google Scholar] [CrossRef]
- Ali, S.; Li, S.; Jaleel, W.; Khan, M.M.; Wang, J.T.; Zhou, X.M. Using a two-sex life table tool to calculate the fitness of Orius strigicollis as a predator of Pectinophora gossypiella. Insects 2020, 11, 275. [Google Scholar] [CrossRef]
- Farooq, M.; Shakeel, M.; Iftikhar, A.; Shahid, M.R.; Zhu, X. Age-stage, two-sex life tables of the lady beetle (Coleoptera: Coccinellidae) feeding on different aphid species. J. Econ. Entomol. 2018, 111, 575–585. [Google Scholar] [CrossRef]
- Bussaman, P.; Sa-uth, C.; Chandrapatya, A.; Atlihan, R.; Gokce, A.; Saska, P.; Chi, H. Fast population growth in physogastry reproduction of Luciaphorus perniciosus rack (Acari: Pygmephordae) at different temperatures. J. Econ. Entomol. 2017, 110, 1397–1403. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chi, H.; Liu, T.X. Demography and parasitic effectiveness of Aphelinus asychis reared from Sitobion avenae as a biological control agent of Myzus persicae reared on chili pepper and cabbage. Biol. Control 2016, 92, 111–119. [Google Scholar] [CrossRef]
- Reddy, G.V.P.; Chi, H. Demographic comparison of sweet potato weevil reared on a major host, Ipomoea batatas, and an alternative host, I. triloba. Sci. Rep. 2015, 5, 11871. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2021. Available online: http://140.120.197.173/Ecology (accessed on 9 February 2022).
- Chi, H. CONSUME-MSChart: A Computer Program for Consumption Rate Analysis Based on the Age Stage, Two-Sex Life Table. 2021. Available online: http://140.120.197.173/ecology (accessed on 9 February 2022).
- Chi, H.; You, M.S.; Atlıhan, R.; Smith, C.L.; Kavousi, A.; Özgökçe, M.S.; Güncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2019, 40, 103–124. [Google Scholar] [CrossRef]
- Chen, Y.M.; Iqbal, A.; Lv, R.E.; Wang, X.G.; Desneux, N.; Zang, L.S. Chinese oak silkworm Antherae pernyi egg, a suitable factitious host for rearing eupelmid egg parasitoids. Pest Manag. Sci. 2022, 78. [Google Scholar] [CrossRef]
- Chen, Y.M.; Sun, J.W.; Iqbal, A.; Lu, R.; Wang, H.; Zang, L.S. An investigation of Caligula japonica (Lepidoptera: Saturniidae) egg distribution and associated parasitoids on walnut trees (Juglans regia L.) in northwestern China. Int. J. Pest Manag. 2020, 1–8. [Google Scholar] [CrossRef]
- Gibson, G.A.P. Revision of the Old world genus Mesocomys Cameron (Hymenoptera: Eupelmidae). Zootaxa 2021, 4901, 1–92. [Google Scholar] [CrossRef]
- Amir-Maafi, M.; Chi, H.; Chen, Z.Z.; Xu, Y.Y. Innovative bootstrap-match technique for life table set up. Entomol. Gen. 2022, 42. [Google Scholar] [CrossRef]
- Mou, D.F.; Lee, C.C.; Smith, C.L.; Chi, H. Using viable eggs to accurately determine the demographic and predation potential of Harmonia dimidiata (Coleoptera: Coccinellidae). J. Appl. Entomol. 2015, 139, 579–591. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 1936. [Google Scholar] [CrossRef]
- Wei, M.F.; Chi, H.; Guo, Y.F.; Li, X.W.; Zhao, L.L.; Ma, R.Y. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri and P. communis (Rosales: Rosaceae) pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, Y.M.; Yang, X.B.; Lv, R.E.; Desneux, N.; Zang, L.S. Parasitism and suitability of Aprostocetus brevipedicellus on Chinese oak silkworm, Antheraea pernyi, a dominant factitious host. Insects 2021, 12, 694. [Google Scholar] [CrossRef]
- Mochiah, M.B.; Ngi-Song, A.J.; Overholt, W.A.; Botchey, M. Host suitability of four cereal stem borers (Lepidoptera: Crambidae, Noctuidae) for different geographic populations of Cotesia sesamiae (Cameron) (Hymenoptera: Braconidae) in Kenya. Biol. Control 2001, 21, 285–292. [Google Scholar] [CrossRef]
- Le Masurier, A.D. Effect of host size on clutch size in Cotesia glomerata. J. Anim. Ecol. 1991, 60, 107–118. [Google Scholar] [CrossRef]
- Jiang, X.J.; Zhou, Z.J. Effects of substitute host pupal stages on feeding and oviposition of Scleroderma sichuanensis (Hymenoptera: Bethylidae). Guangxi For. Sci. 2005, 34, 179–180. [Google Scholar]
- Wang, X.Y.; Yang, Z.Q. Host adaptations of the generalist parasitoids and some factors influencing the choice of hosts. Acta Ecol. Sin. 2010, 30, 1615–1627. [Google Scholar]
- Vinson, S.B. The general host selection behavior of parasitoid Hymenoptera and a comparison of initial strategies utilized by larvaphagous and oophagous species. Biol. Control 1998, 11, 79–96. [Google Scholar] [CrossRef]
- Zhang, M. The influence of the thickness of host egg chorion on the longevity and fecundity of Trichogramma confusum. Nat. Enemies Insects 1999, 21, 150–151. [Google Scholar]
- Pennacchio, F.; Digilio, M.C.; Tremblay, E.; Tranfaglia, A. Host recognition and acceptance behaviour in two aphid parasitoid species: Aphidius ervi and Aphidius microlophii (Hymenoptera: Braconidae). Bull. Entomol. Res. 1994, 84, 57–64. [Google Scholar] [CrossRef]
- Shin, S.S.; Chun, Y.S.; Ryoo, M.I. Functional and numerical responses of Anisopteromalus calandrae and Lariophagus distinguendus (Hymenoptera: Pteromalidae) to the various densities of an alternative host, Callosobruchus chinensis. Korean J. Entomol. 1994, 24, 199–206. [Google Scholar]
- Xu, W.; Wen, X.Y.; Hou, Y.Y.; Desneux, N.; Ali, A.; Zang, L.S. Suitability of Chinese oak silkworm eggs for the multigenerational rearing of the parasitoid Trichogramma leucaniae. PLoS ONE 2020, 15, e0231098. [Google Scholar] [CrossRef] [PubMed]
- Giunti, G.; Canale, A.; Messing, R.H.; Donati, E.; Stefanini, C.; Michaud, J.P.; Benelli, G. Parasitoid learning: Current knowledge and implications for biological control. Biol. Control 2015, 90, 208–219. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.B.; Lin, Y.; Zang, L.S.; Tian, C.Y.; Ruan, C.C. Effect of fertilized, unfertilized, and UV-irradiated hosts on parasitism and suitability for Trichogramma parasitoids. Entomol. Exp. Appl. 2016, 161, 50–56. [Google Scholar] [CrossRef]
- Lu, A.P. Sexual differentiation of parasitic Hymenoptera through nutrition 3. The changes of sexual ratio after the transplantation of egg. Suppl. J. Sun YatSen Univ. 1995, 3, 80–83. [Google Scholar]
- Yang, X.; Qu, Y.L.; Wu, Z.Y.; Lin, Y.; Ruan, C.C.; Desneux, N.; Zang, L.S. Parasitism and suitability of fertilized and nonfertilized eggs of the rice striped stem borer, Chilo suppressalis (Lepidoptera: Crambidae), for Trichogramma parasitoids. J. Econ. Entomol. 2016, 109, 1524–1528. [Google Scholar] [CrossRef]
- Krugner, R. Suitability of non-fertilized eggs of Homalodisca vitripennis for the egg parasitoid Gonatocerus morrilli. BioControl 2014, 59, 167–174. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).