
Host Plant Association and Distribution of the Onion Thrips, Thrips tabaci Cryptic Species Complex



Simple Summary
Abstract
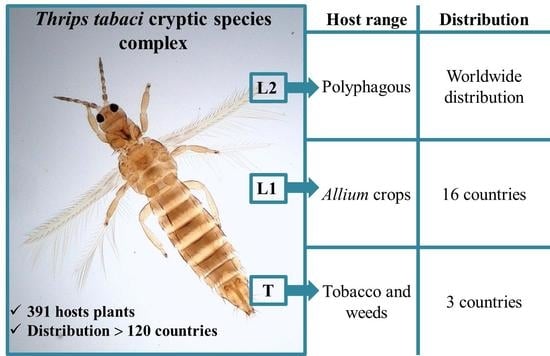
1. Introduction
2. Materials and Methods
2.1. Host Plant Range and Distribution in the Onion Thrips Cryptic Species Complex
2.2. Host Plant Range of Onion Thrips Regardless of Lineages
3. Results and Discussion
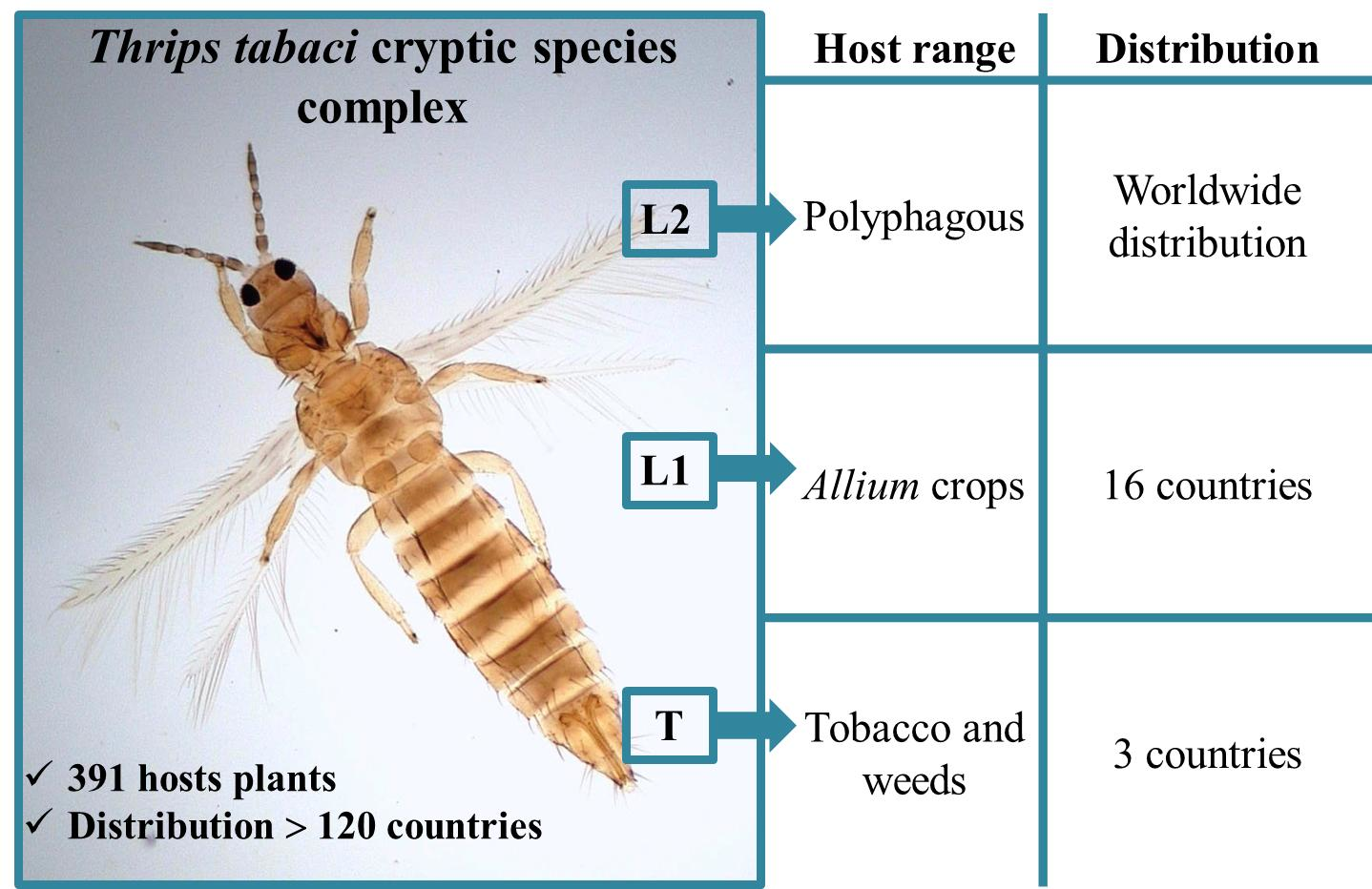
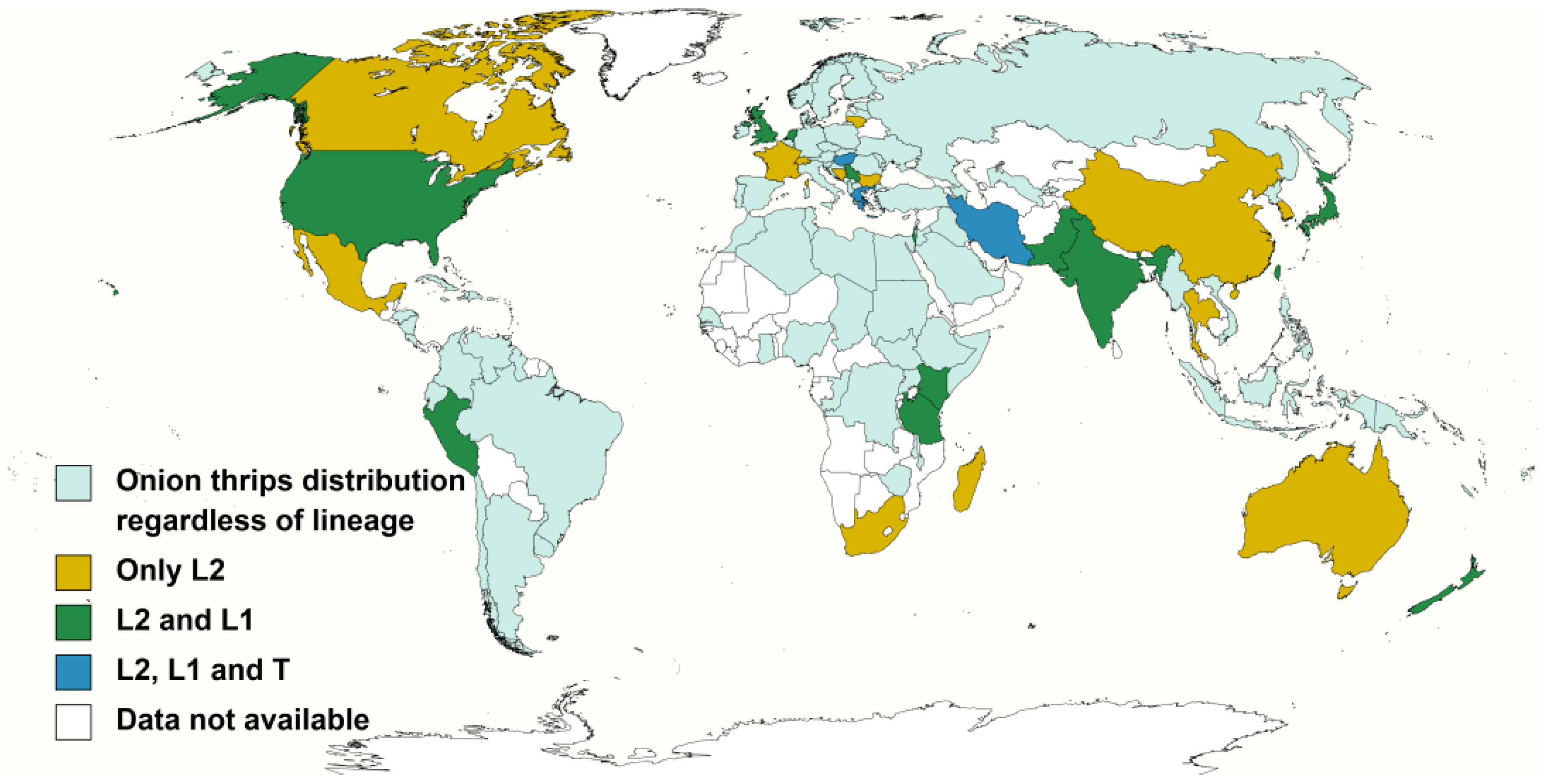
3.1. Host Plants and Distribution of the Onion Thrips Cryptic Species Complex Determined by Molecular Identification Tools

{kind=link}
{kind=link}
{kind=link}
| Plant Species | Thrips tabaci Lineages | |
|---|---|---|
| L2 | L1 | |
| Amaryllidaceae | - | - |
| Allium ampeloprasum L. | A [27,34]; N/A [40] | A [27,34]; N/A [40,67]; R [42,68] |
| A. cepa L. | A [30,36,41,69,70,71]; L [33]; N/A [40,72,73]; R [38] | A [30,41,70]; A, L [33]; N/A [40,42,67,72,74,75]; R [76] |
| A. fistulosum L. | A [77,78]; N/A [15,40] | A [77,78]; N/A [15] |
| A. sativum L. | N/A [40] | - |
| A. tuberosum Rottler ex Spr. | A [69,77] | A [77] |
| A. × wakegi Araki | N/A [40] | - |
| Allium spp. | N/A [79] | - |
| Apiaceae | - | - |
| Ammi majus L. | A, L [80] | - |
| Coriandrum sativum L. | N/A [42] | - |
| Eryngium sp. | N/A [73] | - |
| Asparagaceae | - | - |
| Asparagus officinalis L. | N/A [15,40,73] | - |
| Asteraceae | - | - |
| Arctotheca calendula (L.) Levyns | A [36] | - |
| Bidens pilosa L. | - | N/A [81] |
| Chrysanthemum sp. | A [36] | - |
| Cichorium endivia L. | N/A [73] | - |
| Dittrichia viscosa (L.) Gr. | A [29] | - |
| Erigeron annuus (L.) Pers. | N/A [42] | - |
| Liatris sp. | N/A [40] | N/A [40] |
| Santolina chamaecyparissus L. | N/A [42] | - |
| Balsaminaceae | - | - |
| Impatiens sp. | A [36] | - |
| Boraginaceae | - | - |
| Echium plantagineum L. | A, L [80] | - |
| Brassicaceae | - | - |
| Brassica carinata A.Braun | - | N/A [72] |
| B. oleracea L. | A [71]; N/A [79]; R [38,42] | R [32,33,38] |
| Brassica spp. | N/A [79] | N/A [81] |
| Raphanus raphanistrum subsp. sativus (L.) Domin | N/A [40,79] | - |
| R. raphanistrum L. | N/A [79] | - |
| Caprifoliaceae | - | - |
| Lonicera caprifolium L. | N/A [42] | - |
| Caryophyllaceae | - | - |
| Dianthus sp. | N/A [40] | N/A [40] |
| Cucurbitaceae | - | - |
| Cucumis sativus L. | A [29]; N/A [67] | - |
| Ebenaceae | - | - |
| Diospyros kaki L.f. | A, L [14]; N/A [40] | - |
| Fabaceae | - | - |
| Medicago sativa L. | A [36]; A, L [80] | - |
| Phaseolus vulgaris L. | N/A [67]; R [36,70] | - |
| Pisum sativum L. | N/A [73] | N/A [81] |
| Trifolium repens L. | N/A [40] | - |
| Vicia faba L. | R [14,15,39,40,77] | R [40,77] |
| Gentianaceae | - | - |
| Eustoma grandiflorum (Raf.) Sh. | N/A [40] | - |
| Lamiaceae | - | - |
| Ocimum basilicum L. | N/A [74] | N/A [74] |
| Malvaceae | - | - |
| Gossypium hirsutum L. | A, L [80] | - |
| Poaceae | - | - |
| Secale cereale L. | N/A [79] | - |
| Rosaceae | - | - |
| Dasiphora fruticosa (L.) Rydb. | N/A [42] | - |
| Filipendula vulgaris Moench | N/A [42] | - |
| Fragaria×ananassa (Duchesne ex Weston) Duchesne ex Rozier | N/A [73] | - |
| Sorbaria sorbifolia (L.) A.Br. | N/A [42] | - |
| Rutaceae | - | - |
| Citrus unshiu (Yu.Tanaka ex Swingle) Marcow | N/A [40] | - |
| Solanaceae | - | - |
| Capsicum annuum L. | N/A [67] | - |
| Nicotiana tabacum L. | A [27,30] | A [27] |
| Solanum lycopersicum L. | - | N/A [81] |
| S. nigrum L. | A [36] | - |
| S. tuberosum L. | A [36]; N/A [40] | - |
| Urticaceae | - | - |
| Urtica dioica L. | A [29] | - |
| Verbenaceae | - | - |
| Verbena tenuisecta Briq. | A [80] | - |
| Plant species not determined | A [29,82]; N/A [31,40,81]; N/A [83] (GenBank KM535733, KM529437 and KM528653); N/A [84] (KF534480-KF534483) | A [29]; N/A [31,81]; N/A [85] (GenBank accessions FN546159-FN546164) |

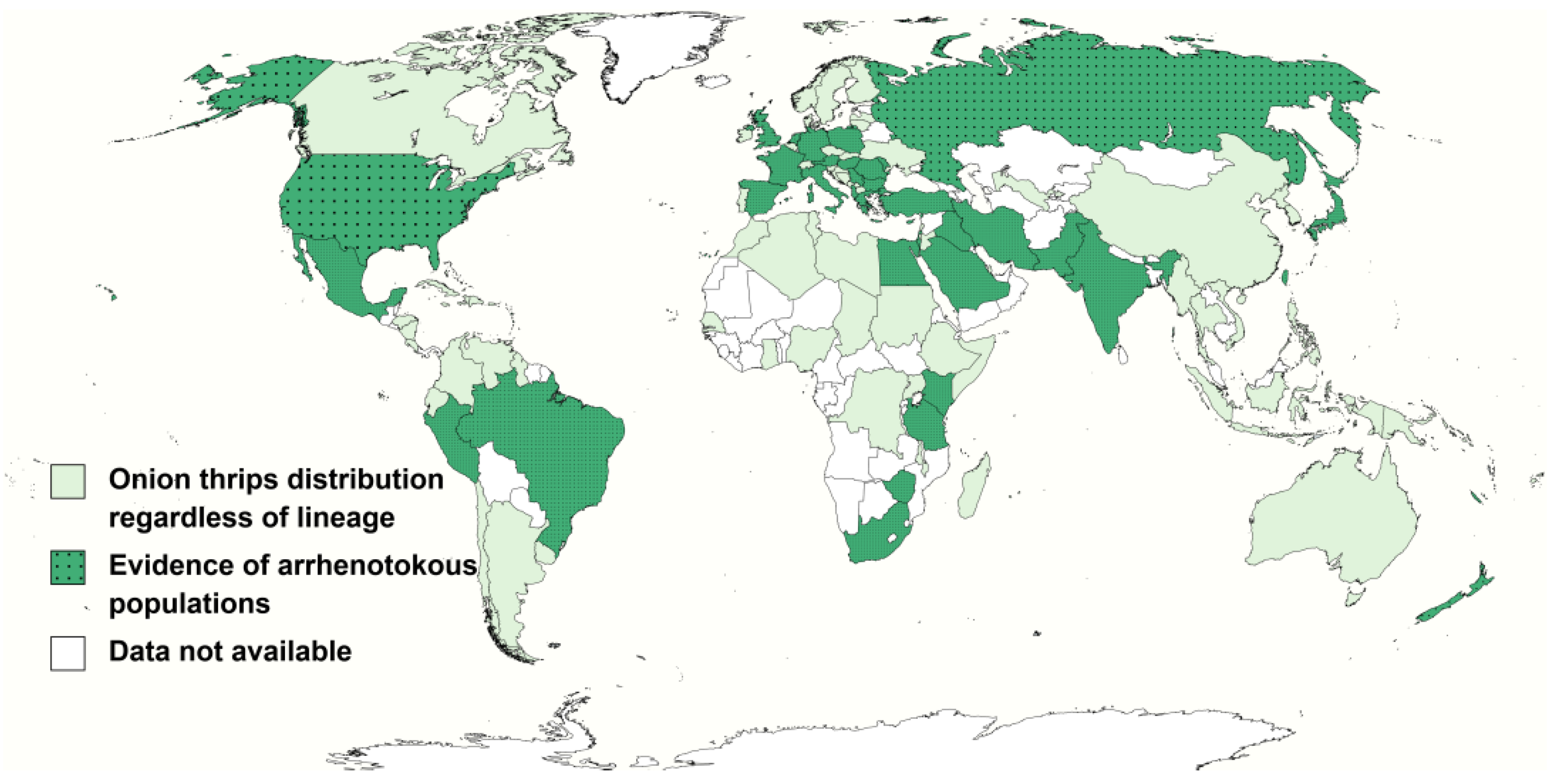
3.2. Host Plants and Distribution of the Onion Thrips Cryptic Species Complex Based on Their Reproductive Mode
| Plant Species | Onion Thrips Reproductive Mode | |
|---|---|---|
| Thelytokous | Arrhenotokous | |
| Amaranthaceae | - | - |
| Amaranthus retroflexus L. | - | R [87] |
| Chenopodium urbicum L. | - | R [28] |
| Amaryllidaceae | - | - |
| Allium ampeloprasum L. | N/A [46]; R [10,88] | R [10,28,42] |
| A. cepa L. | L [33,43]; N/A [40,79]; R [32] | L [33,43]; N/A [42,79]; R [28,32] |
| A. fistulosum L. | A [77] | A [77] |
| A. schoenoprasum L. | - | R [28] |
| A. tuberosum Rottler ex Spreng. | A [77] | A [77] |
| A. × wakegi Araki | N/A [40] | - |
| Allium sp. | N/A [37,79] | - |
| Apiaceae | - | - |
| Coriandrum sativum L. | N/A [42] | - |
| Asparagaceae | - | - |
| Asparagus officinalis L. | N/A [40] | - |
| Asteraceae | - | - |
| Achillea millefolium L. | - | R [28] |
| Arctotheca calendula (L.) Levyns | A [36] | - |
| Chrysanthemum sp. | A [36] | - |
| Cineraria sp. | A [57] | - |
| Emilia coccinea (Sims) G.Don | R [89] | - |
| E. sonchifolia (L.) DC. ex DC. | R [37] | R [37] |
| Erigeron annuus (L.) Pers. | N/A [42] | - |
| Galinsoga parviflora Cav. | - | R [28] |
| Lactuca serriola L. | - | R [87] |
| Santolina chamaecyparissus L. | N/A [42] | - |
| Sonchus oleraceus (L.) L. | - | R [87] |
| Taraxacum officinale Web | - | R [28] |
| Balsaminaceae | - | - |
| Impatiens sp. | A [36] | R [45] |
| Brassicaceae | - | - |
| Brassica oleracea L. | N/A [32,79]; R [33,38,42,43,75] | N/A [32]; R [33,38,43] |
| Brassica sp. | N/A [79] | - |
| Raphanus raphanistrum subsp. sativus (L.) Domin | N/A [37,40,79] | - |
| R. raphanistrum L. | N/A [37,79] | - |
| Caprifoliaceae | - | - |
| Lonicera caprifolium L. | N/A [42] | - |
| Cucurbitaceae | - | - |
| Cucumis sativus L. | R [90] | - |
| Ebenaceae | - | - |
| Diospyros kaki L.f. | A, L [14,39]; N/A [40] | - |
| Fabaceae | - | - |
| Medicago sativa L. | A [36] | - |
| Phaseolus vulgaris L. | R [36,57,70,91] | R [45] |
| Trifolium repens L. | N/A [40] | - |
| Fabaceae | - | - |
| Vicia faba L. | R [14,15,39,40,77] | R [77] |
| V. sativa subsp. nigra (L.) Ehrh. | A [14,39] | - |
| Lamiaceae | - | - |
| Glechoma hederacea L. | - | R [28] |
| Lamium album L. | - | N/A [28] |
| Lamium spp. | - | R [28] |
| Malvaceae | - | - |
| Gossypium hirsutum L. | R [92] | - |
| Papaveraceae | - | - |
| Chelidonium majus L. | - | R [28] |
| Plantaginaceae | - | - |
| Plantago major L. | - | R [28] |
| Poaceae | - | - |
| Bromus inermis Leyss. | - | R [28] |
| Elymus repens (L.) Gould | - | R [28] |
| Poa annua L. | - | R [28] |
| Secale cereale L. | N/A [37,79] | - |
| Primulaceae | - | - |
| Cyclamen sp. | A [57] | - |
| Rosaceae | - | - |
| Dasiphora fruticosa (L.) Rydb. | N/A [42] | - |
| Filipendula vulgaris M. | N/A [42] | - |
| Sorbaria sorbifolia (L.) A.Braun | N/A [42] | - |
| Rutaceae | - | - |
| Citrus unshiu (Yu.Tanaka ex Swingle) Marcow | N/A [40] | - |
| Solanaceae | - | - |
| Capsicum sp. | A [57] | - |
| Datura stramonium L. | R [10,45] | R [10,28,45,87] |
| Nicotiana tabacum L. | - | A [10,46]; R [10,28,42,44,87] |
| Solanum nigrum L. | A [36] | R [28,87] |
| S. tuberosum L. | A [36]; N/A [40] | R [28] |

3.3. Host Plant Range of Onion Thrips Regardless of Lineage
| Plant Species | References |
|---|---|
| Aizoaceae | - |
| Mesembryanthemum forskahlii Hochst. ex Boiss. | [23] |
| Alstroemeriaceae | - |
| Alstroemeria sp. †,‡,§,¶ | [8] |
| Amaranthaceae | - |
| Amaranthus albus L. † | [89] |
| A. caudatus L. † | [23] |
| A. retroflexus L. †,‡ | [7,22,89] |
| Atriplex hortensis L. | [22] |
| A. sagittata Borkh. | [22] |
| A. tatarica L. | [119] |
| Beta vulgaris L. † | [22,120] |
| Chenopodium album L. †,‡ | [22,23,121] |
| C. murale L. † | [23] |
| C. quinoa Willd. †,‡,§ | [122] |
| C. urbicum L. † | [28] |
| Dysphania ambrosioides (L.) Mosyakin and Clemants † | [23] |
| Spinacia oleracea L. † | [22,120] |
| Amaryllidaceae | - |
| Allium ampeloprasum L. †,‡ | [10,21,108] |
| A. ascalonicum L. | [123] |
| A. cepa L. †,‡ | [21,38,43] |
| A. fistulosum L. ‡ | [94,124,125] |
| A. oleraceum L. | [22] |
| A. sativum L. †,‡ | [22,111,124] |
| A. schoenoprasum L. ‡ | [28] |
| Allium sp. | [9] |
| Narcissus sp. | [126] |
| Apiaceae | - |
| Ammi majus L. † | [21,23,80] |
| Anethum graveolens L. | [112] |
| Anthriscus sylvestris (L.) Hoffm. | [118] |
| Apium graveolens L. † | [120] |
| Conium maculatum L. † | [52] |
| Daucus carota L. † | [89] |
| D. sativus Roehl. | [22] |
| Daucus sp. | [112] |
| Petroselinum crispum (Mill.) Fuss | [22,127] |
| Apocynaceae | - |
| Asclepias syriaca L. | [17,52] |
| Araceae | - |
| Arum sp. †,§ | [113,127] |
| Araliaceae | - |
| Polyscias guilfoylei L.H.Bailey | [89] |
| Schefflera sp. | [8] |
| Asparagaceae | - |
| Asparagus officinalis L. | [124,128] |
| Asparagus sp. | [24] |
| Hyacinthus sp. | [129] |
| Yucca sp. | [89] |
| Asteraceae | - |
| Acanthospermum australe (Loefl.) Kuntze | [89] |
| Achillea clypeolata Sm. | [22] |
| A. millefolium L. † | [22,24,28] |
| A. nobilis L. | [22] |
| A. ptarmica L. | [24] |
| A. salicifolia Besser | [118] |
| Achillea spp. | [130] |
| Ageratum conyzoides (L.) L. † | [23,89,128] |
| Ambrosia artemisiifolia L. † | [7,17,119] |
| A. maritima L. | [21] |
| Anthemis arvensis L. † | [7,22,111] |
| A. melampodina Delile | [21,23] |
| A. retusa Delile | [23] |
| Arctium minus (Hill) Bernh. †,‡ | [52] |
| Arctotheca calendula (L.) Levyns † | [80] |
| Artemisia princeps Pamp. | [125] |
| A. vulgaris L. † | [24] |
| Aster spp. † | [24] |
| Bidens pilosa L. † | [80,89,128] |
| Brachyscome iberidifolia Benth. † | [131] |
| Calendula arvensis (Vaill.) L. † | [132] |
| C. officinalis L. † | [59] |
| Callistephus chinensis (L.) Nees † | [128] |
| Carduus acanthoides L. † | [22,133] |
| C. pycnocephalus L. † | [22] |
| Carlina vulgaris L. | [118] |
| Centaurea calcitrapa L. † | [23] |
| C. cyanus L. † | [22] |
| Chrysanthemum spp. †,‡,§ | [24,59,89] |
| Cichorium endivia L. † | [23,120] |
| C. intybus L. †,‡ | [23,120,121] |
| Cineraria sp. † | [24,113] |
| Cirsium arvense (L.) Scop. † | [22,52] |
| C. syriacum (L.) Gaertn. | [21,23] |
| Cota tinctoria (L.) J.Gay † | [22] |
| Cyanthillium cinereum (L.) H.Rob. | [89] |
| Dahlia pinnata Cav. | [22] |
| Dahlia sp. †,§ | [8] |
| Emilia coccinea (Sims) G.Don † | [89,128] |
| E. sonchifolia (L.) DC. Ex DC. †,§,¶ | [37,134] |
| Emilia spp. † | [128] |
| Erigeron annuus (L.) Pers. | [135] |
| Erigeron bonariensis L. † | [23] |
| E. canadensis L. †,‡ | [22,89,128] |
| Eupatorium cannabinum L. † | [22] |
| Galinsoga parviflora Cav. † | [7,17,119] |
| Helianthus annuus L. †,‡ | [22] |
| Helminthotheca echioides (L.) Holub † | [133] |
| Hypochaeris radicata L. | [24] |
| Inula candida (L.) Cass. | [22] |
| I. helenium L. † | [22] |
| I. salicifolia Gueldenst. | [136] |
| Jacobaea maritima (L.) Pelser and Meijden † | [22] |
| Lactuca sativa L. †,§ | [22,121,128] |
| L. serriola L. †,‡ | [64,87] |
| Laphangium luteoalbum (L.) Tzv. | [23] |
| Launaea nudicaulis (L.) Hook.f. | [23] |
| Leucanthemum vulgare Lam. † | [8,22] |
| Matricaria chamomilla L. † | [22,121,133] |
| Matricaria spp. | [24] |
| Pluchea indica (L.) Less. | [89] |
| Pulicaria arabica (L.) Cass. | [23] |
| Santolina chamaecyparissus L. | [137] |
| Senecio cruentus (L’Hér.) DC. †,§ | [12] |
| S. glaucus subsp. coronopifolius C. Alex. | [21] |
| S. pogonias Cabrera | [121] |
| S. umbrosus Waldst. and Kit. | [118] |
| S. viscosus L. | [24] |
| S. vulgaris L. †,‡ | [119,135] |
| Senecio spp. † | [24,130] |
| Silybum marianum (L.) Gaertn. † | [23] |
| Solidago canadensis L. † | [52] |
| S. gigantea Aiton | [118] |
| S. virgaurea L. | [118] |
| Solidago spp. † | [24] |
| Sonchus asper (L.) Hill †,‡ | [138] |
| S. oleraceus (L.) L. †,‡ | [23,58,80] |
| Tanacetum cinerariifolium (Trevir.) Sch.Bip. † | [21,22] |
| T. vulgare L. | [118] |
| Taraxacum officinale (L.) Weber ex F.H.Wigg. †,‡ | [7,28,52] |
| Tithonia rotundifolia (Mill.) S.F.Blake † | [89] |
| Tragopogon dubius Scop. ‡ | [64] |
| T. porrifolius L. † | [22] |
| Tripleurospermum indorum (L.) Sch.Bip. † | [22] |
| T. maritmum (L.) Koch | [119] |
| Verbesina encelioides (Cav.) Benth. and Hook.f. ex A.Gray † | [55] |
| Xanthium strumarium L. † | [80] |
| Zinnia elegans L. † | [22] |
| Balsaminaceae | - |
| Impatiens sp. †,‡,§,¶ | [45] |
| Boraginaceae | - |
| Anchusa azurea Mill. † | [111] |
| Bothriospermum tenellum (Hornem.) Fisch. and Mey. | [89] |
| Echium plantagineum L. | [58,80,133] |
| Heliotropium europaeum L. † | [22] |
| Brassicaceae | - |
| Alyssum spp. | [26] |
| Armoracia rusticana P.Gaertn., B.Mey. and Scherb. | [139] |
| Barbarea vulgaris R.Br. † | [52] |
| Berteroa incana (L.) DC. | [22] |
| Biscutella auriculata L. | [111] |
| Brassica cretica Lam. | [22] |
| B. juncea (L.) Czern. | [140] |
| B. napus L. † | [80] |
| B. nigra (L.) K.Koch | [89] |
| B. oleracea L. † | [21,38,60] |
| B. rapa L. † | [22,133] |
| Brassica sp. | [125] |
| Capsella bursa-pastoris (L.) Medik. † | [52,80,121] |
| Cochlearia sp. | [24] |
| Descurainia sophia (L.) Webb ex Prantl ‡ | [64] |
| Diplotaxis sp. | [111] |
| Erophila verna (L.) DC. | [22] |
| Eruca vesicaria (L.) Cav. | [21,23,121] |
| Erysimum sp. | [24] |
| Lepidium bonariense L. | [80] |
| L. didymum L. † | [89] |
| L. draba L. † | [22,135] |
| L. latifolium L. | [22] |
| L. virginicum L. † | [52] |
| Lobularia spp. | [26] |
| Raphanus raphanistrum subsp. sativus (L.) Domin | [23,89,133] |
| R. raphanistrum L. † | [52] |
| Raphanus sp. | [141] |
| Rapistrum rugosum (L.) All. | [58,80,133] |
| Sinapis alba L. | [112] |
| S. arvensis L. †,‡ | [22,52] |
| Sisymbrium irio L. | [21,23,121] |
| S. officinale (L.) Scop. | [22] |
| Sisymbrium sp. | [111] |
| Thlaspi arvense L. | [52] |
| Bromeliaceae | - |
| Ananas comosus (L.) Merr. † | [89,128] |
| Cactaceae | - |
| Opuntia sp. † | [24] |
| Calceolariaceae | - |
| Calceolaria sp. † | [127] |
| Campanulaceae | - |
| Campanula sp. † | [24] |
| Caricaceae | - |
| Carica papaya L. † | [120,142] |
| Caryophyllaceae | - |
| Dianthus barbatus L. | [8] |
| D. campestris M.Bieb. | [22] |
| D. caryophyllus L. | [21,59] |
| D. corymbosus Sm. | [22] |
| Dianthus spp. † | [8,24,89] |
| Spergularia rubra (L.) J.Pr. and C.Pr. | [24] |
| Stellaria media (L.) Vill. † | [7,51,119] |
| Commelinaceae | - |
| Commelina sp. | [120] |
| Murdannia nudiflora (L.) Brenan | [120] |
| Tradescantia sp. † | [24] |
| Convolvulaceae | - |
| Calystegia sepium (L.) R. Br. † | [52] |
| Convolvulus arvensis L. †,‡ | [7,21,121] |
| Ipomoea batatas (L.) Lam. † | [22,125] |
| I. indica (Burm.) Merr. † | [128] |
| Operculina aequisepala (Domin) | [80] |
| Crassulaceae | - |
| Sedum acre L. | [118] |
| Sedum sp. † | [24] |
| Cucurbitaceae | - |
| Citrullus lanatus (Thunb.) Matsum. and Nakai † | [21,22] |
| Cucumis melo L. † | [22] |
| C. sativus L. †,§ | [8,90,125] |
| Cucurbita maxima Duchesne † | [22] |
| C. moschata Duchesne † | [21,22] |
| C. pepo L. † | [21,22] |
| Ebenaceae | - |
| Diospyros kaki L.f. | [14] |
| Ericaceae | - |
| Ledum palustre L. | [118] |
| Euphorbiaceae | - |
| Chrozophora tinctoria (L.) A.Juss. | [21] |
| Euphorbia hirta L. | [89] |
| Ricinus communis L. †,‡ | [23] |
| Fabaceae | - |
| Anthyllis vulneraria L. | [118] |
| Anthyllis sp. | [24] |
| Arachis hypogaea L. † | [22] |
| Astragalus cicer L. | [22] |
| Bauhinia sp. | [23] |
| Cajanus cajan (L.) Millsp. † | [89] |
| Cicer arietinum L. † | [117] |
| Crotalaria juncea L. † | [89] |
| C. saltiana Andrews | [89] |
| C. spectabilis Roth † | [89] |
| Cytisus nigricans L. | [135] |
| Delonix regia (Boj.ex Hook.) Raf. | [21] |
| Galega officinalis L. | [133] |
| Glycine max (L.) Merr. †,§ | [22,143] |
| Hippocrepis emerus (L.) Lassen | [22] |
| Lathyrus sp. | [24] |
| Lens culinaris Medik † | [21] |
| Lotus corniculatus L. | [22] |
| L. tenuis Waldst. and Kit. | [133] |
| Lotus sp. | [24] |
| Lupinus angustifolius L. † | [144] |
| Lupinus sp. † | [89] |
| Medicago lupulina L. † | [22,52] |
| M. polymorpha var. vulgaris (Benth.) Shinners | [80] |
| M. sativa L. | [21,80,89] |
| Melilotus albus Medik. | [118] |
| M. indicus (L.) All. † | [21,23] |
| M. messanensis (L.) All. | [23] |
| M. officinalis (L.) Pall. † | [22,119] |
| Mimosa pudica L. | [89] |
| Ornithopus compressus L. | [22] |
| Phaseolus vulgaris L. †,¶ | [21,22,121] |
| Pisum sativum L. †,§ | [89,117,128] |
| P. sativum subsp. elatius (M.Bieb.) Asch. and Graebn. | [22] |
| Senna occidentalis (L.) Link † | [89] |
| Trifolium alexandrinum L. | [21,112] |
| T. pratense L. | [22,112] |
| T. repens L. † | [7,22,133] |
| Trifolium resupinatum L. | [23] |
| Trigonella foenum-graecum L. | [21] |
| T. laciniata L. | [23] |
| Vicia faba L. †,§,¶ | [21,77,80] |
| V. sativa L. ‡ | [80,121] |
| Vigna mungo (L.) Hepper † | [145] |
| V. unguiculata subsp. unguiculata (L.) Walp. † | [89] |
| Gentianaceae | - |
| Eustoma russellianum (Hook.) G.Don †,‡ | [146] |
| Geraniaceae | - |
| Erodium cicutarium (L.) L’Hér. ‡ | [22] |
| Geranium pratense L. | [118] |
| G. pusillum L. | [22] |
| G. pyrenaicum Burm.f. | [22] |
| Gesneriaceae | - |
| Saintpaulia brevipilosa B.L.Burtt | [131] |
| Hypericaceae | - |
| Hypericum perforatum L. | [22] |
| Iridaceae | - |
| Freesia sp. | [8] |
| Gladiolus spp. † | [8] |
| Juncaginaceae | - |
| Triglochin maritima L. | [24] |
| Lamiaceae | - |
| Glechoma hederacea L. | [22,28] |
| Lamium album L. | [28] |
| L. amplexicaule L. † | [7,23,121] |
| L. purpureum L. † | [22,52] |
| Lamium spp. | [28] |
| Nepeta cataria L. † | [52] |
| Nepeta spp. | [24] |
| Origanum vulgare L. | [121] |
| Rosmarinus spp. | [26] |
| Salvia sp. † | [24] |
| Teucrium scorodonia L. | [24] |
| Linaceae | - |
| Linum usitatissimum L. | [147] |
| Lythraceae | - |
| Cuphea hyssopifolia Kunth | [89] |
| Malvaceae | - |
| Abutilon theophrasti Medik. | [22] |
| Alcea rosea L. | [21,22] |
| Althaea officinalis L. † | [22] |
| Althaea sp. †,§ | [24] |
| Gossypium arboreum L. | [22] |
| Gossypium barbadense L. † | [22] |
| G. hirsutum L. † | [22,80] |
| Gossypium spp. | [21] |
| Malva neglecta Wallr. † | [22,52,64] |
| M. parviflora L. † | [121] |
| M. sylvestris L. † | [22] |
| Malva sp. † | [89] |
| Sphaeralcea miniata (Cav.) Spach | [121] |
| Tilia cordata Mill. | [22] |
| T. tomentosa Moench | [22] |
| Waltheria indica L. | [89] |
| Moraceae | - |
| Ficus carica L. | [148] |
| Morus alba L. | [21,22] |
| M. nigra L. | [22] |
| Myrtaceae | - |
| Eucalyptus spp. | [26] |
| Nyctaginaceae | - |
| Boerhavia glabrata Blume | [80] |
| Onagraceae | - |
| Oenothera biennis L. † | [52] |
| Oxalidaceae | - |
| Oxalis corniculata L. † | [23] |
| O. debilis Kunth | [89] |
| O. stricta L. † | [52] |
| Oxalis sp. † | [24] |
| Papaveraceae | - |
| Chelidonium majus L. | [28] |
| Fumaria capreolata L. | [80] |
| Papaver rhoeas L. † | [111] |
| Phytolaccaceae | - |
| Phytolacca acinosa Roxb. | [89] |
| Plantaginaceae | - |
| Digitalis sp. † | [24] |
| Linaria vulgaris Mill. | [22,112] |
| Plantago lanceolata L. †,‡ | [52,121] |
| P. major L. † | [28] |
| P. maritima L. | [24] |
| Veronica austriaca L. | [22] |
| V. persica Poir. † | [121] |
| Plumbaginaceae | - |
| Armeria sp. | [24] |
| Poaceae | - |
| Avena fatua L. † | [80] |
| A. sativa L. | [149] |
| Avena sp. | [52] |
| Bromus inermis Leyss. ‡ | [28] |
| Cenchrus americanus (L.) Morr. | [22] |
| Dactylis glomerata L. | [111] |
| Digitaria sanguinalis (L.) Scop. † | [22,89] |
| D. violascens Link | [89] |
| Eleusine indica (L.) Gaertn. †,‡ | [89] |
| Elymus repens (L.) Gould | [28] |
| Hordeum maritimum With. | [21,23] |
| H. vulgare L. | [21,60,149] |
| Lolium sp. | [52] |
| Phleum pratense L. | [149] |
| Poa annua L. † | [28] |
| Polypogon monspeliensis (L.) Desf | [23] |
| P. viridis (Gouan) Breistr. | [23] |
| Secale cereale L. | [150] |
| Setaria verticillata (L.) P.Beauv. | [22] |
| S. viridis (L.) P.Beauv. †,‡ | [22] |
| Sorghum halepense (L.) Pers. | [22] |
| Triticum aestivum L. ‡ | [58,149,150] |
| Polygonaceae | - |
| Persicaria maculosa Gray † | [22] |
| P. orientalis (L.) Spach | [80] |
| Polygonum aviculare L. † | [80] |
| Rumex crispus L. †,‡ | [58,80] |
| R. dentatus L. | [23] |
| R. obtusifolius L. | [119] |
| Portulacaceae | - |
| Portulaca oleracea L. †,‡,§ | [89] |
| Primulaceae | - |
| Cyclamen coum Mill. | [22] |
| Cyclamen spp. † | [8,127] |
| Ranunculaceae | - |
| Helleborus odorus Waldst. and Kit. | [22] |
| Ranunculus arvensis L. † | [22] |
| R. polyanthemos L. | [22] |
| Resedaceae | - |
| Reseda spp. | [26] |
| Rosaceae | - |
| Fragaria × ananassa (Duchesne ex Weston) Duchesne ex Rozier | [151] |
| Malus domestica Borkh. | [22] |
| Potentilla alba L. | [22] |
| Prunus avium (L.) L. | [152] |
| P. domestica L. | [22] |
| P. persica (L.) Batsch | [24] |
| Prunus spp. | [26] |
| Pyrus communis L. | [22] |
| Rosa canina L. | [22] |
| R. damascena Mill. | [22] |
| Rosa sp. †,‡,§ | [9,127] |
| Rubiaceae | - |
| Galium aparine L. † | [22] |
| G. verum L. † | [118,119] |
| Galium sp. † | [24] |
| Richardsonia scabra L. † | [89,128] |
| Rutaceae | - |
| Citrus unshiu (Yu.Tanaka ex Swingle) Marcow | [153] |
| Citrus spp. | [154] |
| Salicaceae | - |
| Salix mucronata Thunb. | [21] |
| Salix sp. | [23] |
| Scrophulariaceae | - |
| Verbascum lychnitis L. | [118] |
| V. thapsus L. † | [52] |
| Solanaceae | - |
| Alkekengi officinarum Moench † | [22] |
| Atropa bella-donna L. † | [22] |
| Capsicum annuum L. †,‡,§ | [120,155] |
| Datura stramonium L. †,‡,§,¶ | [28,45,120] |
| Hyoscyamus niger L. † | [22] |
| Lycium barbarum L. | [22] |
| Nicandra physalodes (L.) Gaertn. † | [89] |
| Nicotiana glauca Graham † | [128] |
| N. rustica L. †,‡,§ | [22] |
| N. tabacum L. †,§,¶ | [10,28,107] |
| Nicotiana sp. † | [112] |
| Petunia hybrida Vilm †,‡,§ | [22] |
| Solanum americanum Mill. † | [89,128] |
| S. dulcamara L. † | [22] |
| S. lycopersicum L. †,‡,§ | [21,22,23] |
| S. melongena L. †,§ | [22,120,155] |
| S. nigrum L. †,‡ | [22,28,87] |
| S. tuberosum L. †,‡,§ | [21,28,120] |
| Tropaeolaceae | - |
| Tropaeolum majus L. †,§ | [22] |
| Urticaceae | - |
| Urtica dioica L. † | [52] |
| Verbenaceae | - |
| Lantana camara L. | [89] |
| Pitraea cuneato-ovata (Cav.) Caro | [121] |
| Stachytarpheta cayennensis (Rich.) Vahl | [89] |
| Verbena bonariensis L. | [89] |
| V. officinalis L. † | [80] |
| V. tenuisecta Briq. | [58] |
| Viburnaceae | - |
| Sambucus ebulus L. | [22] |
| Vitaceae | - |
| Vitis vinifera L. | [22] |
| Zygophyllaceae | - |
| Tribulus terrestris L. †,‡ | [22,80] |
4. Conclusions
Author Contributions
Funding
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lewis, T.; Mound, L.A.; Nakahara, S.; Childers, C.C. Major crops infested by thrips with main symptoms and predominant injurious species. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 675–709. [Google Scholar]
- CAB International. Thrips Tabaci (Onion Thrips). Available online: https://www.cabi.org/isc/datasheet/53746 (accessed on 10 September 2021).
- Jenser, G.; Lipcsei, S.; Szénási, A.; Hudák, K. Host range of the arrhenotokous populations of Thrips tabaci (Thysanoptera: Thripidae). Acta Phytopathol. Entomol. Hung. 2006, 41, 297–303. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mound, L.A.; Marullo, R. The Thrips of Central and South America: An Introduction (Insecta: Thysanoptera); Virendra, K.G., Ed.; Associated Publishers: Gainesville, FL, USA, 1996; pp. 16–18, 199–200. [Google Scholar]
- Shelton, A.M.; North, R.C. Species composition and phenology of Thysanoptera within field crops adjacent to cabbage fields. Environ. Entomol. 1986, 15, 513–519. [Google Scholar] [CrossRef]
- Orosz, S.; Éliás, D.; Balog, E.; Tóth, F. Investigation of Thysanoptera populations in Hungarian greenhouses. Acta Univ. Sapientiae Agric. Environ. 2017, 9, 140–158. [Google Scholar] [CrossRef]
- Mantel, W.P.; van de Vrie, M. A contribution to the knowledge of Thysanoptera in ornamental and bulbous crops in the Netherlands. Acta Phytopathol. Entomol. Hung. 1988, 23, 301–311. [Google Scholar]
- Vierbergen, G. Thysanoptera intercepted in the Netherlands on plant products from Ethiopia, with description of two new species of the genus Thrips. Zootaxa 2014, 3765, 269–278. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K.; Peters, D.; Katis, N.I. The efficiency by which Thrips tabaci populations transmit Tomato spotted wilt virus depends on their host preference and reproductive strategy. Phytopathology 2002, 92, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Schwartz, H.F.; Cramer, C.S.; Havey, M.J.; Pappu, H.R. Iris yellow spot virus (Tospovirus: Bunyaviridae): From obscurity to research priority. Mol. Plant Pathol. 2015, 16, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Rasoulpour, R.; Izadpanah, K. Characterisation of cineraria strain of Tomato yellow ring virus from Iran. Australas. Plant Pathol. 2007, 36, 286. [Google Scholar] [CrossRef]
- Hassani-Mehraban, A.; Dullemans, A.M.; Verhoeven, J.T.J.; Roenhorst, J.W.; Peters, D.; van der Vlugt, R.A.A.; Kormelink, R. Alstroemeria yellow spot virus (AYSV): A new orthotospovirus species within a growing Eurasian clade. Arch. Virol. 2019, 164, 117–126. [Google Scholar] [CrossRef]
- Morishita, M. Pyrethroid-resistant onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), infesting persimmon fruit. Appl. Entomol. Zool. 2008, 43, 25–31. [Google Scholar] [CrossRef]
- Aizawa, M.; Watanabe, T.; Kumano, A.; Miyatake, T.; Sonoda, S. Cypermethrin resistance and reproductive types in onion thrips, Thrips tabaci (Thysanoptera: Thripidae). J. Pestic. Sci. 2016, 41, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.K.; Garg, H.; Gill, A.K.; Gillett-Kaufman, J.L.; Nault, B.A. Onion thrips (Thysanoptera: Thripidae) biology, ecology, and management in onion production systems. J. Integr. Pest Manag. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Jenser, G.; Almási, A.; Kazinczi, G.; Takács, A.; Szénási, Á.; Gáborjányi, R. Ecological background of the epidemics of Tomato spotted wilt virus in central Europe. Acta Phytopathol. Entomol. Hung. 2009, 44, 213–223. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K. Management of the spread of Tomato spotted wilt virus in tobacco crops with insecticides based on estimates of thrips infestation and virus incidence. Plant Dis. 2008, 92, 1012–1020. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gent, D.H.; du Toit, L.J.; Fichtner, S.F.; Mohan, S.K.; Pappu, H.R.; Schwartz, H.F. Iris yellow spot virus: An emerging threat to onion bulb and seed production. Plant Dis. 2006, 90, 1468–1480. [Google Scholar] [CrossRef]
- Lewis, T. Flight and dispersal. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Wallingford, UK, 1997; pp. 175–196. [Google Scholar]
- Ghabn, A.A.A.E. Contribution to the knowledge of the biology of Thrips tabaci Lind. in Egypt. Bull. Soc. Entomol. Egypte 1948, 32, 123–174. [Google Scholar]
- Dimitrov, A. Host plants of Thrips tabaci Lind. in Bulgaria. Plant Sci. 1977, 15, 139–144. [Google Scholar]
- Priesner, H. A Monograph of the Thysanoptera of the Egyptian Deserts; Publications de Institut du Désert d’Égypte: Cairo, Egypt, 1960; pp. 22–41. [Google Scholar]
- Morison, G.D. Notes on Thysanoptera found on flax (Linum usitatissimum L.) in the British isles. Ann. Appl. Biol. 1943, 30, 251–259. [Google Scholar] [CrossRef]
- Mound, L.A. Homologies and host-plant specificity: Recurrent problems in the study of thrips. Fla. Entomol. 2013, 96, 318–322. [Google Scholar] [CrossRef]
- Marullo, R. Host-plant range and relationships in the Italian thrips fauna. Acta Phytopathol. Entomol. Hung. 2004, 39, 243–254. [Google Scholar] [CrossRef]
- Brunner, P.C.; Chatzivassiliou, E.K.; Katis, N.I.; Frey, J.E. Host-associated genetic differentiation in Thrips tabaci (Insecta; Thysanoptera), as determined from mtDNA sequence data. Heredity 2004, 93, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Zawirska, I. Untersuchungen über zwei biologische typen von Thrips tabaci Lind. (Thysanoptera, Thripidae) in der VR Polen. Arch. Phytopathol. Pflanzenschutz 1976, 12, 411–422. [Google Scholar] [CrossRef]
- Glover, R.H.; Collins, D.W.; Walsh, K.; Boonham, N. Assessment of loci for DNA barcoding in the genus Thrips (Thysanoptera:Thripidae). Mol. Ecol. Resour. 2010, 10, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Fekrat, L.; Manzari, S.; Shishehbor, P. Molecular and morphometric variation of Thrips tabaci Lindeman (Thysanoptera: Thripidae) populations on onion and tobacco in Iran. J. Agric. Sci. Technol. 2014, 16, 1505–1516. [Google Scholar]
- Iftikhar, R.; Ashfaq, M.; Rasool, A.; Hebert, P.D.N. DNA barcode analysis of thrips (Thysanoptera) diversity in Pakistan reveals cryptic species complexes. PLoS ONE 2016, 11, e0146014. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Wang, P.; Fail, J.; Shelton, A.M. Detection of gene flow from sexual to asexual lineages in Thrips tabaci (Thysanoptera: Thripidae). PLoS ONE 2015, 10, e0138353. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.L.; Nault, B.A.; Vargo, E.L.; Kennedy, G.G. Restricted gene flow among lineages of Thrips tabaci supports genetic divergence among cryptic species groups. PLoS ONE 2016, 11, e0163882. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yoshimura, J.; Hasegawa, E. Coexistence of sexual individuals and genetically isolated asexual counterparts in a thrips. Sci. Rep. 2013, 3, 3286. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hasegawa, E. A female-biased sex ratio reduces the twofold cost of sex. Sci. Rep. 2016, 6, 23982. [Google Scholar] [CrossRef]
- Westmore, G.C.; Poke, F.S.; Allen, G.R.; Wilson, C.R. Genetic and host-associated differentiation within Thrips tabaci Lindeman (Thysanoptera: Thripidae) and its links to Tomato spotted wilt virus-vector competence. Heredity 2013, 111, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.L.; Kennedy, G.G. Specific insect-virus interactions are responsible for variation in competency of different Thrips tabaci isolines to transmit different Tomato spotted wilt virus isolates. PLoS ONE 2013, 8, e54567. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Fail, J.; Wang, P.; Feng, J.; Shelton, A.M. Performance of arrhenotokous and thelytokous Thrips tabaci (Thysanoptera: Thripidae) on onion and cabbage and its implications on evolution and pest management. J. Econ. Entomol. 2014, 107, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Toda, S.; Morishita, M. Identification of three point mutations on the sodium channel gene in pyrethroid-resistant Thrips tabaci (Thysanoptera: Thripidae). J. Econ. Entomol. 2009, 102, 2296–2300. [Google Scholar] [CrossRef] [PubMed]
- Toda, S.; Murai, T. Phylogenetic analysis based on mitochondrial COI gene sequences in Thrips tabaci Lindeman (Thysanoptera: Thripidae) in relation to reproductive forms and geographic distribution. Appl. Entomol. Zool. 2007, 42, 309–316. [Google Scholar] [CrossRef]
- Kobayashi, K.; Hasegawa, E. Discrimination of reproductive forms of Thrips tabaci (Thysanoptera: Thripidae) by PCR with sequence specific primers. J. Econ. Entomol. 2012, 105, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Farkas, P.; György, Z.; Tóth, A.; Sojnóczki, A.; Fail, J. A simple molecular identification method of the Thrips tabaci (Thysanoptera: Thripidae) cryptic species complex. Bull. Entomol. Res. 2020, 110, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Nault, B.A.; Shelton, A.M.; Gangloff-kaufmann, J.L.; Clark, M.E.; Werren, J.L.; Cabrera-la Rosa, J.C.; Kennedy, G.G. Reproductive modes in onion thrips (Thysanoptera: Thripidae) populations from New York onion fields. Environ. Entomol. 2006, 35, 1264–1271. [Google Scholar] [CrossRef]
- Woldemelak, W.A. The existence of deuterotokous reproduction mode in the T. tabaci (Thysanoptera: Thripidae) cryptic species complex. J. Hortic. Res. 2020, 28, 21–28. [Google Scholar] [CrossRef]
- Wijkamp, I. Distinct levels of specificity in thrips transmission of tospoviruses. Phytopathology 1995, 85, 1069. [Google Scholar] [CrossRef]
- Karadjova, O.; Krumov, V. TSWV transmission efficiency of an arrhenotokous and a thelytokous population of Thrips tabaci. Acta Phytopathol. Entomol. Hung. 2008, 43, 289–292. [Google Scholar] [CrossRef]
- Hristova, D.; Karadjova, O.; Yankulova, M.; Heinze, C.; Adam, G. A survey of tospoviruses in Bulgaria. J. Phytopathol. 2001, 149, 745–749. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K.; Nagata, T.; Katis, N.I.; Peters, D. Transmission of Tomato spotted wilt tospovirus by Thrips tabaci populations originating from leek. Plant Pathol. 1999, 48, 700–706. [Google Scholar] [CrossRef]
- Inoue, T.; Murai, T.; Natsuaki, T. An effective system for detecting Iris yellow spot virus transmission by Thrips tabaci. Plant Pathol. 2010, 59, 422–428. [Google Scholar] [CrossRef]
- Jenser, G.; Gaborjanyi, R.; Szenasi, A.; Almasi, A.; Grasselli, M. Significance of hibernated Thrips tabaci Lindeman (Thysan., Thripidae) adults in the epidemic of Tomato spotted wilt virus. J. Appl. Entomol. 2003, 127, 7–11. [Google Scholar] [CrossRef]
- Jenser, G.; Vierbergen, B.; Szénási, A. Thysanoptera larvae living on chickweed (Stellaria media Linnaeus) under continental climatic conditions. Acta Phytopathol. Entomol. Hung. 2007, 42, 387–390. [Google Scholar] [CrossRef]
- Smith, E.A.; Ditommaso, A.; Fuchs, M.; Shelton, A.M.; Nault, B.A. Weed hosts for onion thrips (Thysanoptera: Thripidae) and their potential role in the epidemiology of Iris yellow spot virus in an onion ecosystem. Environ. Entomol. 2011, 40, 194–203. [Google Scholar] [CrossRef]
- Trdan, S.; Žnidarčič, D.; Valič, N.; Rozman, L.; Vidrih, M. Intercropping against onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae) in onion production: On the suitability of orchard grass, lacy phacelia, and buckwheat as alternatives for white clover. J. Plant Dis. Prot. 2006, 113, 24–30. [Google Scholar]
- Buckland, K.R.; Alston, D.G.; Reeve, J.R.; Nischwitz, C.; Drost, D. Trap crops in onion to reduce onion thrips and Iris yellow spot virus. Southwest. Entomol. 2017, 42, 73–90. [Google Scholar] [CrossRef]
- Calvert, F.; Hollingsworth, R.G.; Wall, M.; Follett, P.A. Survey of flowering plants in Hawaii as potential banker plants of anthocorid predators for thrips control. J. Asia Pac. Entomol. 2019, 22, 638–644. [Google Scholar] [CrossRef]
- Ren, X.; Wu, S.; Xing, Z.; Gao, Y.; Cai, W.; Lei, Z. Abundances of thrips on plants in vegetative and flowering stages are related to plant volatiles. J. Appl. Entomol. 2020, 144, 732–742. [Google Scholar] [CrossRef]
- Tavella, L.; Tedeschi, R.; Mason, G.; Roggero, P. Efficiency of north-western Italian thrips populations in transmitting tospoviruses. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; Marullo, R., Mound, L.A., Eds.; Australian National Insect Collection: Canberra, Australia, 2002; pp. 81–86. [Google Scholar]
- Milne, M.; Walter, G.H. Host species and plant part specificity of the polyphagous onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), in an Australian cotton-growing area. Aust. J. Entomol. 1998, 37, 115–119. [Google Scholar] [CrossRef]
- Zamar, M.I.; Neder, L.E.; Linares, M.A.; Hamity, V.C.; Contreras, E.F.; Gómez, G. Tisanópteros (Insecta) asociados a plantas ornamentales de Jujuy (Argentina). Rev. Agron. Noroeste Argent. 2014, 34, 261–262. [Google Scholar]
- de Borbón, C.; Gracia, O.; De Santis, L. Survey of Thysanoptera occurring on vegetable crops as potential tospovirus vectors in Mendoza, Argentina. Rev. Soc. Entomol. Argen. 1999, 58, 59–66. [Google Scholar]
- Teulon, D.A.J.; Groninger, J.W.; Cameron, E.A. Distribution and host plant associations of Taeniothrips inconsequens (Thysanoptera: Thripidae). Environ. Entomol. 1994, 23, 587–611. [Google Scholar] [CrossRef]
- Parrella, G.; Gognalons, P.; Gebre-Selassiè, K.; Vovlas, C.; Marchoux, G. An update of the host range of Tomato spotted wilt virus. J. Plant Pathol. 2003, 85, 227–264. [Google Scholar]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019; p. 2936. [Google Scholar]
- Szostek, S.; Schwartz, H.F. Overwintering sites of Iris yellow spot virus and Thrips tabaci (Thysanoptera: Thripidae) in Colorado. Southwest. Entomol. 2015, 40, 273–290. [Google Scholar] [CrossRef]
- Schwartz, H.F.; Gent, D.H.; Fichtner, S.M.; Otto, K.; Boateng, C.O.; Szostek, S.; Cranshaw, W.S.; Mahaffey, L.A. Thrips tabaci (Thysanoptera: Thripidae) and Iris yellow spot virus associated with onion transplants, onion volunteers, and weeds in Colorado. Southwest. Entomol. 2014, 39, 691–704. [Google Scholar] [CrossRef]
- Hassani-Mehraban, A.; Saaijer, J.; Peters, D.; Goldbach, R.; Kormelink, R. A new tomato-infecting tospovirus from Iran. Phytopathology 2005, 95, 852–858. [Google Scholar] [CrossRef]
- Cvrković, T.; Jović, J.; Mitrović, M.; Krstić, O.; Toševski, I. Genetic variability in Thrips tabaci (Insecta: Thysanoptera) living on vegetables in Serbia. In Proceedings of the International Symposium on Current Trends in Plant Protection, Belgrade, Serbia, 25–28 September 2012; Marisavljevic, D., Ed.; Institute for Plant Protection and Environment: Belgrade, Serbia, 2012; pp. 477–482. [Google Scholar]
- Schmidt, K. Thrips obscuratus in New Zealand Vineyards: Its Biology and Effects on Botrytis Cinerea Infection. Ph.D. Thesis, Lincoln University, Christchurch, New Zealand, 2007. [Google Scholar]
- Li, X.; Zhang, Z.; Zhang, J.; Huang, J.; Wang, L.; Li, Y.; Hafeez, M.; Lu, Y. Population genetic diversity and structure of Thrips tabaci (Thysanoptera: Thripidae) on Allium hosts in China, inferred from mitochondrial COI gene sequences. J. Econ. Entomol. 2020, 113, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Gawande, S.J.; Anandhan, S.; Ingle, A.A.; Jacobson, A.; Asokan, R. Heteroplasmy due to coexistence of mtCOI haplotypes from different lineages of the Thrips tabaci cryptic species group. Bull. Entomol. Res. 2017, 107, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Nault, B.A.; Kain, W.C.; Wang, P. Seasonal changes in Thrips tabaci population structure in two cultivated hosts. PLoS ONE 2014, 9, e101791. [Google Scholar] [CrossRef] [PubMed]
- Kadirvel, P.; Srinivasan, R.; Hsu, Y.-C.; Su, F.-C.; de la Peña, R. Application of cytochrome oxidase I sequences for phylogenetic analysis and identification of thrips species occurring on vegetable crops. J. Econ. Entomol. 2013, 106, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Tseng, L.Y.; Chang, N.T.; Tseng, M.J.; Yeh, W.B. Genetic variation of Thrips tabaci Lindeman (Thysanoptera: Thripidae) in the Pacific rim. Formos. Entomol. 2010, 30, 219–234. [Google Scholar] [CrossRef]
- Srinivasan, R.; Guo, F.; Riley, D.G.; Diffie, S.; Gitaitis, R.; Sparks, A.N.; Jeyaprakash, A. Assessment of variation among Thrips tabaci populations from Georgia and Peru based on polymorphisms in mitochondrial cytochrome oxidase I and ribosomal ITS2 sequences. J. Entomol. Sci. 2011, 46, 191–203. [Google Scholar] [CrossRef]
- Sojnóczki, A.; Pájtli, R.D.; Farkas, P.; Fail, J. Comparative study of Thrips tabaci (Lindeman) cytochrome-c oxidase gene subunit I (COI) sequences data. Bodenkultur 2015, 66, 41–45. [Google Scholar]
- Li, X.W.; Fail, J.; Shelton, A.M. Female multiple matings and male harassment and their effects on fitness of arrhenotokous Thrips tabaci (Thysanoptera: Thripidae). Behav. Ecol. Sociobiol. 2015, 69, 1585–1595. [Google Scholar] [CrossRef] [PubMed]
- Sogo, K.; Miura, K.; Aizawa, M.; Watanabe, T.; Stouthamer, R. Genetic structure in relation to reproduction mode in Thrips tabaci (Insecta: Thysanoptera). Appl. Entomol. Zool. 2015, 50, 73–77. [Google Scholar] [CrossRef]
- Takeuchi, R.; Toda, S. Discrimination of two reproductive forms of Thrips tabaci by PCR-RFLP, and distribution of arrhenotokous T. tabaci in Tottori prefecture. Jpn. J. Appl. Entomol. Zool. 2011, 55, 254–257. [Google Scholar] [CrossRef]
- Jacobson, A.L.; Booth, W.; Vargo, E.L.; Kennedy, G.G. Thrips tabaci population genetic structure and polyploidy in relation to competency as a vector of Tomato spotted wilt virus. PLoS ONE 2013, 8, e54484. [Google Scholar] [CrossRef]
- Silva, R.; Hereward, J.P.; Walter, G.H.; Wilson, L.J.; Furlong, M.J. Seasonal abundance of cotton thrips (Thysanoptera: Thripidae) across crop and non-crop vegetation in an Australian cotton producing region. Agric. Ecosyst. Environ. 2018, 256, 226–238. [Google Scholar] [CrossRef]
- Macharia, I.; Backhouse, D.; Skilton, R.; Ateka, E.; Wu, S.-B.; Njahira, M.; Maina, S.; Harvey, J. Diversity of thrips species and vectors of Tomato spotted wilt virus in tomato production systems in Kenya. J. Econ. Entomol. 2015, 108, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Timm, A.E.; Stiller, M.; Frey, J.E. A molecular identification key for economically important thrips species (Thysanoptera: Thripidae) in southern Africa. Afr. Entomol. 2008, 16, 68–75. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; DeWaard, J.R. Barcoding Canada Data Release; University of Guelph, Biodiversity Institute of Ontario: Guelph, ON, Canada, 2014; Unpublished work. [Google Scholar]
- Surviliene, E.; Bernotiene, R.; Valiuskaite, A.; Duchovskiene, L.; Tamosiunas, R.; Rasiukeviciute, N. The Study of Genetic Differentiation of Thrips tabaci (Insecta: Thysanoptera) and Tetranychus urticae (Acari: Tetranychidae) in Lithuania; Nature Research Centre, Institute of Ecology: Vilnius, Lithuania, 2013; Unpublished work. [Google Scholar]
- Glover, R.H.; Collins, D.W. COI DNA Barcodes for the Identification of Thrips; Food and Environment Research Agency, Novel Diagnostic Methods: York, UK, 2009; Unpublished work. [Google Scholar]
- Eckel, C.S.; Cho, K.; Walgenbach, J.F.; Kennedy, G.G.; Moyer, J.W. Variation in thrips species composition in field crops and implications for tomato spotted wilt epidemiology in North Carolina. Entomol. Exp. Appl. 1996, 78, 19–29. [Google Scholar] [CrossRef]
- Chatzivassiliou, E.K.; Peters, D.; Katis, N.I. The role of weeds in the spread of Tomato spotted wilt virus by Thrips tabaci (Thysanoptera: Thripidae) in tobacco Crops. J. Phytopathol. 2007, 155, 699–705. [Google Scholar] [CrossRef]
- Marullo, R.; Grazia, A. Notes on sex ratio and reproductive modes in field populations of two pest thrips species (Thysanoptera, Thripidae) in Italy. Acta Phytopathol. Entomol. Hung. 2012, 47, 117–123. [Google Scholar] [CrossRef]
- Sakimura, K. Life history of Thrips tabaci L. on Emilia sagittata and its host plant range in Hawaii. J. Econ. Entomol. 1932, 25, 884–891. [Google Scholar] [CrossRef]
- van Rijn, P.C.J.; Mollema, C.; Steenhuis-Broers, G.M. Comparative life history studies of Frankliniella occidentalis and Thrips tabaci (Thysanoptera: Thripidae) on cucumber. Bull. Entomol. Res. 1995, 85, 285–297. [Google Scholar] [CrossRef]
- Tedeschi, R.; Ciuffo, M.; Mason, G.; Roggero, P.; Tavella, L. Transmissibility of four tospoviruses by a thelytokous population of Thrips tabaci from Liguria, northwestern Italy. Phytoparasitica 2001, 29, 37–45. [Google Scholar] [CrossRef]
- Klein, M.; Gafni, R. Morphological and molecular variations in thrips population collected on onion plants in Israel. Folia Entomol. Hung. 1996, 57, 57–59. [Google Scholar]
- Murai, T. Parthenogenetic reproduction in Thrips tabaci and Frankliniella intonsa (Insecta: Thysanoptera). In Advances in Invertebrate Reproduction; Hoshi, M., Yamashita, O., Eds.; Elsevier: Amsterdam, The Netherlands, 1990; Volume 5, pp. 357–362. [Google Scholar]
- Aizawa, M.; Watanabe, T.; Kumano, A.; Tamagaki, K.; Sonoda, S. Biotic performances of thelytokous and arrhenotokous strains of Thrips tabaci (Thysanoptera: Thripidae) showing resistance to cypermethrin. Appl. Entomol. Zool. 2018, 53, 11–17. [Google Scholar] [CrossRef]
- Almási, A.; Tóbiás, I.; Bujdos, L.; Jenser, G. Molecular characterisation of Thrips tabaci Lindeman, 1889 (Thysanoptera: Thripidae) populations in Hungary based on the ITS2 sequences. Acta Zool. Acad. Sci. Hung. 2016, 62, 157–164. [Google Scholar] [CrossRef]
- Monteiro, R.C.; Mound, L.A.; Zucchi, R.A. Thrips (Thysanoptera) as pests of plant production in Brazil. Rev. Bras. Entomol. 1999, 43, 163–171. [Google Scholar]
- Etienne, J.; Ryckewaert, P.; Michel, B. Thrips (Insecta: Thysanoptera) of Guadeloupe and Martinique: Updated check-list with new information on their ecology and natural enemies. Fla. Entomol. 2015, 98, 298–304. [Google Scholar] [CrossRef]
- Loredo Varela, R.C. Hungarian University of Agriculture and Life Sciences; Budapest, Hungary, 2019; Unpublished work. [Google Scholar]
- Bhatti, J.S. Species of the genus Thrips from India (Thysanoptera). Syst. Entomol. 1980, 5, 109–166. [Google Scholar] [CrossRef]
- Mound, L.A.; Palmer, J.M. Notes on Thysanoptera from Israel. Entomol.’s Mon. Mag. 1973, 109, 102–106. [Google Scholar]
- Priesner, H. Zur thysanopteren-fauna Albaniens. Sitz. Kais. Akad. Wiss. 1919, 128, 115–144. [Google Scholar]
- Priesner, H. Beitrag zur kenntnis der thysanopteren Oberösterreichs. Jahrb. Oberösterr. Musealver. 1920, 78, 50–63. [Google Scholar]
- Karadjova, O.; Hristova, D.; Adam, G. Epidemiology of tomato spotted wilt tospovirus in Bulgarian tobacco field. Bulg. J. Agric. Sci. 2001, 7, 583–594. [Google Scholar]
- Özden, Ö. The Biodiversity of Invertebrates in Cyprus Ecosystems. Ph.D. Thesis, University of Exeter, Exeter, UK, 2009. [Google Scholar]
- Pizzol, J.; Reynaud, P.; Bresch, C.; Rabasse, J.M.; Biondi, A.; Desneux, N.; Parolin, P.; Poncet, C. Diversity of Thysanoptera species and associated host plants in southern France. J. Mediterr.Ecol. 2017, 15, 13–27. [Google Scholar]
- Boness, M.; Titschack, E.; Schliephake, G. Thysanopterenfunde aus Nord- und Westdeutschland. Faun. Ökol. Mitt. 1992, 6, 319–330. [Google Scholar]
- Lindeman, K.Ė. Die schädlichsten insekten des tabak in Bessarabien. Bull. Soc. Imp. Nat. Moscou 1888, 2, 10–77. [Google Scholar]
- Vierbergen, G.; Ester, A. Natural enemies and sex ratio of Thrips tabaci (Thysanoptera: Thripidae), a major pest of Allium porrum in the Netherlands. Med. Fac. Landbouw. Univ. Gent 2000, 65, 335–342. [Google Scholar]
- Vasiliu-Oromulu, L. The dynamics of the sex ratio index of thrips populations in mountainous meadows. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; Marullo, R., Mound, L.A., Eds.; Australian National Insect Collection: Canberra, Australia, 2002; pp. 315–324. [Google Scholar]
- Rozhina, V.I.; Vierbergen, G. Thrips (Thysanoptera) in the meadows of Kaliningrad province. Entomol. Rev. 2018, 98, 197–203. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Lacasa, A.; Bielza, P.; Meco, R. Dinámica poblacional de Thrips tabaci Lind. (Thysanoptera: Thripidae) sobre liliáceas hortícolas en Castilla-La Mancha. Bol. Sanid. Veg. Plagas 1994, 20, 661–677. [Google Scholar]
- Blunck, H. Thysanopteren aus der Türkei (Thysanoptera). Beitr. Entomol. 1958, 8, 98–111. [Google Scholar] [CrossRef]
- Speyer, E.R. Some common species of the genus Thrips (Thysanoptera). Ann. Appl. Biol. 1934, 21, 120–152. [Google Scholar] [CrossRef]
- Bournier, J.P.; Mound, L.A. Inventaire commenté des Thysanoptères de Nouvelle-Calédonie. Bull. Soc. Entomol. Fr. 2000, 105, 231–240. [Google Scholar] [CrossRef]
- Mound, L.A.; Masumoto, M. The Genus Thrips (Thysanoptera, Thripidae) in Australia, New Caledonia and New Zealand, 1st ed.; Magnolia Press: Auckland, New Zealand, 2005; Volume 1020, ISBN 1877407224. [Google Scholar]
- Jenser, G.; Szénási, Á. Review of the biology and vector capability of Thrips tabaci Lindeman (Thysanoptera: Thripidae). Acta Phytopathol. Entomol. Hung. 2004, 39, 137–155. [Google Scholar] [CrossRef]
- Kirk, W.D.J. A key to the larvae of some common Australian flower thrips (Insecta, Thysanoptera), with a host-plant survey. Aust. J. Zool. 1987, 35, 173. [Google Scholar] [CrossRef]
- Kucharczyk, H. Comparative Morphology of the Second Larval Instar of the Thrips Genus Species (Thysanoptera: Thripidae) Occurring in Poland; Mantis Publishing Company: Olsztyn, Poland, 2010. [Google Scholar]
- Orosz, S.; Juhász, M.; Tôkés, G.; Tóth, F. Occurrence of Thrips tabaci larvae on TSWV host weeds in the surroundings of sweet pepper greenhouses. Acta Phytopathol. Entomol. Hung. 2008, 43, 329–336. [Google Scholar] [CrossRef]
- Sakimura, K. On the host plants of some Hawaiian thrips. Proc. Hawaii. Entomol. Soc. 1939, 10, 251–254. [Google Scholar]
- de Borbón, C. Clave para la identificación del segundo estadío larval de algunos trips comunes (Thysanoptera: Thripidae). Mendoza, Argentina. Rev. Fac. Cienc. Agrar. Univ. Nac. Cuyo 2007, 39, 69–81. [Google Scholar]
- Bulger, M.A. Transmission and field spread of Raspberry bushy dwarf virus. Plant Dis. 1990, 74, 514. [Google Scholar] [CrossRef]
- Hardy, V.G.; Teakle, D.S. Transmission of Sowbane mosaic virus by Thrips tabaci in the presence and absence of virus-carrying pollen. Ann. Appl. Biol. 1992, 121, 315–320. [Google Scholar] [CrossRef]
- Monteiro, R.C. The Thysanoptera fauna of Brazil. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 2–7 July 2001; Marullo, R., Mound, L.A., Eds.; Australian National Insect Collection: Canberra, Australia, 2002; pp. 325–340. [Google Scholar]
- Miyazaki, M.; Kudo, I. Descriptions of thrips larvae which are noteworthy on cultivated plants (Thysanoptera: Thripidae). I. Species occurring on solanaceous and cucurbitaceous crops. Akitu 1986, 79, 209–212. [Google Scholar]
- Blanton, F.S. Notes on some thrips collected in the vicinity of Babylon, Long Island, NY. J. N. Y. Entomol. Soc. 1939, 47, 83–94. [Google Scholar]
- Speyer, E.R.; Parr, W.J. The external structure of some thysanopterous larvae. Trans. R. Entomol. Soc. Lond. 1941, 91, 559–635. [Google Scholar] [CrossRef]
- Sakimura, K. A survey of host ranges of thrips in and around Hawaiian pineapple fields. Proc. Hawaii. Entomol. Soc. 1937, 9, 415–427. [Google Scholar]
- Franssen, C.J.H.; Mantel, W.P. Thrips tabaci op hyacintebollen. Neth. J. Plant Pathol. 1965, 71, 67–71. [Google Scholar] [CrossRef]
- Morison, G.D. Thysanoptera of the London area, part II. Lond. Nat. 1948, 27, 37–75. [Google Scholar]
- Thungrabeab, M.; Blaeser, P.; Sengonca, C. Effect of temperature and host plant on the efficacy of different entomopathogenic fungi from Thailand against Frankliniella occidentalis (Pergande) and Thrips tabaci Lindeman (Thysanoptera: Thripidae) in the laboratory. J. Plant Dis. Prot. 2006, 113, 181–187. [Google Scholar] [CrossRef]
- Belaam-Kort, I. Etude des Thrips (Thysanoptera) en Vergers d’agrumes en Tunisie en vue de l’établissement d’une stratégie de gestion Raisonnée des Espèces Nuisibles. Ph.D. Thesis, Institute National Agronomique de Tunisie, Tunis, Tunisia, 2020. [Google Scholar]
- Carrizo, P.; Amela García, M.T. Vegetación espontánea en el cinturón hortícola platense hospedante de thripidae (Thysanoptera) vectores de tospovirus: Riesgo relativo como componente epidemiológico. RIA Rev. Investig. Agropecu. 2019, 45, 383–394. [Google Scholar]
- Sakimura, K. Two species of thrips non-vectors of the spotted wilt virus. J. Econ. Entomol. 1946, 39, 398–399. [Google Scholar] [CrossRef]
- Orosz, S. Hajtatott Paprika Állományokban és Azok Környezetében élő Thysanoptera Populációk Vizsgálata. Ph.D. Thesis, Szent István Egyetem, Gödöllő, Hungary, 2012. [Google Scholar]
- Przybylska, A.; Fiedler, Ż.; Kucharczyk, H.; Obrępalska-Stęplowska, A. Detection of the quarantine species Thrips palmi by loop-mediated isothermal amplification. PLoS ONE 2015, 10, e0122033. [Google Scholar] [CrossRef]
- Goldarazena, A.; Loomans, A.J.M.; Jordama, R. Parasitic and parasitoid enemies of thrips (Insecta, Thysanoptera) in northern Spain, an introduction. In Proceedings of the Sixth International Symposium on Thysanoptera, Antalya, Turkey, 27 April–1 May 1998; Vierbergen, G., Tunç, I., Eds.; Akdeniz University: Antalya, Turkey, 1999; pp. 37–42. [Google Scholar]
- Nischwitz, C.; Srinivasan, R.; Sundaraj, S.; Mullis, S.W.; McInnes, B.; Gitaitis, R.D. Geographical distribution and survival of Iris yellow spot virus in spiny sowthistle, Sonchus asper, in Georgia. Plant Dis. 2012, 96, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, C. Thysanoptera associated with horseradish in Illinois. Entomol. News 1979, 90, 236–238. [Google Scholar]
- Burgess, L.; Weegar, H.H. Thrips (Thysanoptera) in canola crops in Saskatchewan. Can. Entomol. 1988, 120, 815–819. [Google Scholar] [CrossRef]
- Franssen, C.J.H.; Mantel, W.P. Lijst van in Nederland aangetroffen Thysanoptera, met beknopte aantekeningen over hun levenswijze en hun betekenis voor onze cultuurgewassen. Tijdschr. Entomol. 1962, 105, 33–97. [Google Scholar]
- Holdaway, F.G. Insect pests of papaya and their control. In Papaya Production in the Hawaiian Islands; Jones, W.W., Storey, W.B., Parris, G.K., Holdaway, F.G., Eds.; Hawaii Agricultural Experiment Station of the University of Hawaii: Honolulu, HI, USA, 1941; pp. 46–51. [Google Scholar]
- Ábrahám, R. Thrips species associated with soybean in Hungary. Acta Phytopathol. Entomol. Hung. 2008, 43, 211–218. [Google Scholar] [CrossRef]
- Hurej, M.; Kucharczyk, H.; Twardowski, J.P.; Kozak, M. Thrips (Thysanoptera) associated with narrow-leafed lupin (Lupinus angustifolius L., 1753) intercropped with spring triticale (x Triticosecale Wittm. ex A. Camus, 1927). Rom. Agric. Res. 2014, 31, 337–345. [Google Scholar]
- Amin, P.W.; Palmer, J.M. Identification of groundnut Thysanoptera. Trop. Pest Manag. 1985, 31, 286–291. [Google Scholar] [CrossRef]
- Kritzman, A.; Beckelman, H.; Alexandrov, S.; Cohen, J.; Lampel, M.; Zeidan, M.; Raccah, B.; Gera, A. lisianthus leaf necrosis: A new disease of lisianthus caused by Iris yellow spot virus. Plant Dis. 2000, 84, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Hurej, M.; Kucharczyk, H.; Twardowski, J.P.; Kotecki, A. Thrips (Thysanoptera) associated with two genetically modified types of linseed (Linum usitatissimum L.). J. Plant Dis. Prot. 2017, 124, 81–91. [Google Scholar] [CrossRef]
- Takahashi, A.; Yamauchi, T. Biology of thrips which attack the inside of fig fruits (Ficus carica L.). Bull. Shizuoka Pref. Citrus Exp. Sta. 1986, 22, 21–31. [Google Scholar]
- North, R.C.; Shelton, A.M. Overwintering of the onion thrips, Thrips tabaci (Thysanoptera: Thripidae), in New York. Environ. Entomol. 1986, 15, 695–699. [Google Scholar] [CrossRef]
- Zawirska, I.; Walkowski, W. Fauna and importance of thrips (Thysanoptera) for rye and winter wheat in Poland. Part I. J. Plant Prot. Res. 2000, 40, 35–55. [Google Scholar]
- Nielsen, H.; Sigsgaard, L.; Kobro, S.; Jensen, N.L.; Jacobsen, S.K. Species composition of thrips (Thysanoptera: Thripidae) in strawberry high tunnels in Denmark. Insects 2021, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- de Borbón, C.M.; Becerra, V.; Bonomo, V.; Mazzitelli, E.; Calvo, M. Trips (Insecta: Thysanoptera) en montes de cerezo en Mendoza, Argentina. Rev. Fac. Cienc. Agrar. Univ. Nac. Cuyo 2008, 40, 1–10. [Google Scholar]
- Tsuchiya, M. Infestation and oviposition of Thrips tabaci (Lindeman) on Satsuma mandarin (Citrus unshiu Marc.). Jpn. J. Appl. Entomol. Zool. 2002, 46, 217–224. [Google Scholar] [CrossRef]
- Navarro-Campos, C.; Aguilar, A.; Garcia-Marí, F. Aggregation pattern, sampling plan, and intervention threshold for Pezothrips kellyanus in citrus groves. Entomol. Exp. Appl. 2012, 142, 130–139. [Google Scholar] [CrossRef]
- Madadi, H.; Kharazi-Pakdel, A.; Ashouri, A.; Neyshabouri, J.M. Life history parameters of Thrips tabaci (Thys.: Thripidae) on cucumber, sweet pepper and eggplant under laboratory conditions. J. Entomol. Soc. Iran 2006, 25, 45–62. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loredo Varela, R.C.; Fail, J. Host Plant Association and Distribution of the Onion Thrips, Thrips tabaci Cryptic Species Complex. Insects 2022, 13, 298. https://doi.org/10.3390/insects13030298
Loredo Varela RC, Fail J. Host Plant Association and Distribution of the Onion Thrips, Thrips tabaci Cryptic Species Complex. Insects. 2022; 13(3):298. https://doi.org/10.3390/insects13030298
Chicago/Turabian StyleLoredo Varela, Roberto Carlos, and József Fail. 2022. "Host Plant Association and Distribution of the Onion Thrips, Thrips tabaci Cryptic Species Complex" Insects 13, no. 3: 298. https://doi.org/10.3390/insects13030298
APA StyleLoredo Varela, R. C., & Fail, J. (2022). Host Plant Association and Distribution of the Onion Thrips, Thrips tabaci Cryptic Species Complex. Insects, 13(3), 298. https://doi.org/10.3390/insects13030298