
Prolucanus beipiaoensis gen. et sp. nov.: The First Fossil Species of Lucaninae (Coleoptera: Lucanidae) from the Early Cretaceous of Northeastern China




{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
- Body length: length between the apex of mandible to the elytral apex along the midline;
- Elytral length: length between the basal border and the apex of elytra along suture;
- Elytral width: widest part of both elytra combined;
- Head length: length between the anterior apex of clypeus and the posterior margin of occiput along the midline;
- Head width: widest part of head;
- Mandible length: length from the apex of mandible to its base at anterior margin of the head;
- Pronotal length: length of the pronotum along the midline;
- Pronotal width: widest part of pronotum.
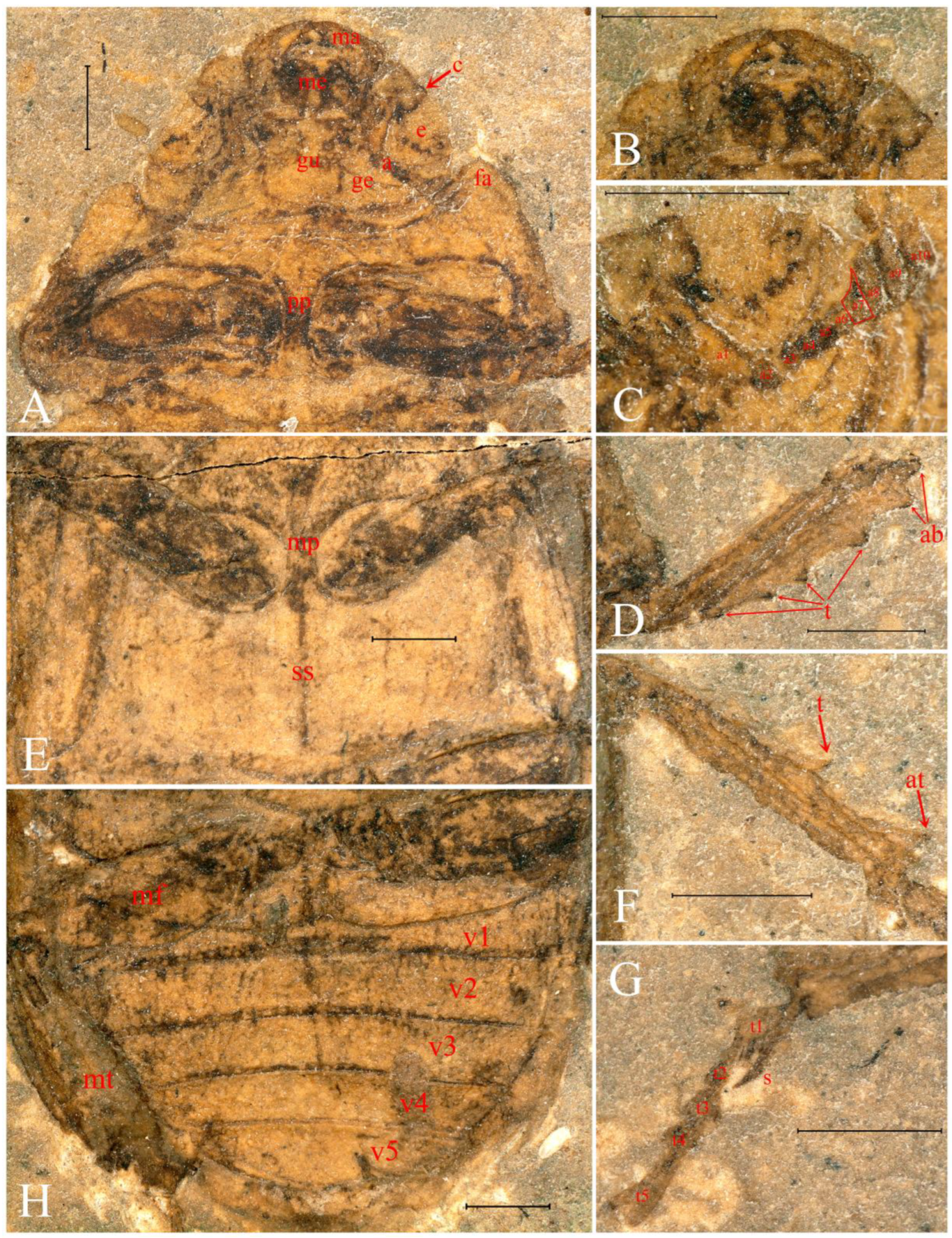
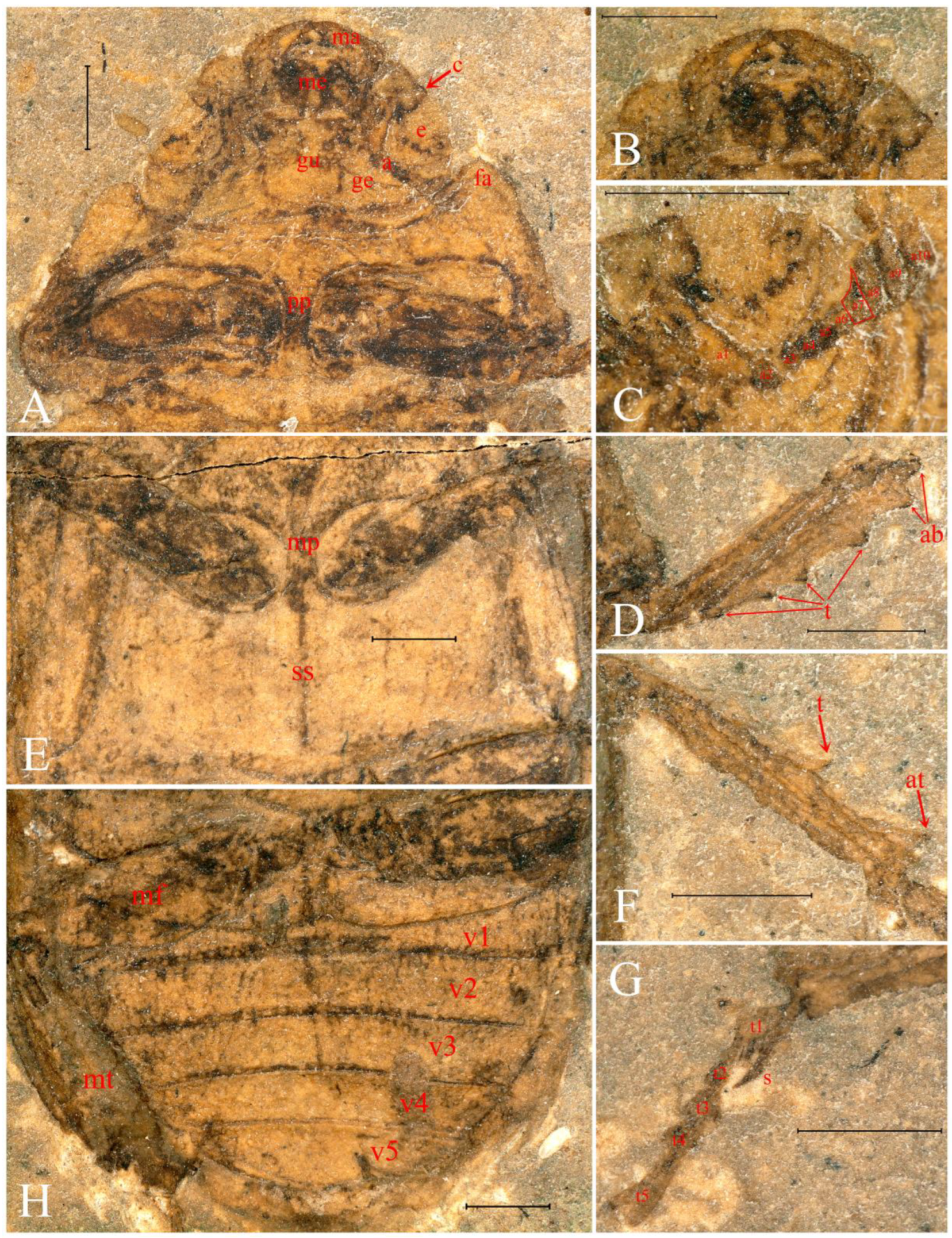
3. Results
- Antenna geniculate..............................................................................Prolucanus gen. nov.
- −
- Antenna partially or not geniculate....................................................................................2
- 2
- Head width exceeding more than half of pronotal width...............................................3
- −
- Head not broader than half the width of pronotum........................................................4
- 3
- Prosternal process broad.................................................Juraesaluss Nikolajev et al., 2011
- −
- Prosternal process narrow...............................Prosinodendron Bai, Ren, and Yang, 2012
- 4
- Length of scape longer than length of pedicel..............Sinaesalus Nikolajev et al., 2011
- −
- Length of scape equal to or shorter than length of pedicel.........................................................................................................................Litholamprima Nikolajev and Ren, 2015
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crowson, R.A. The Natural Classification of the Families of Coleoptera; E.W. Classey Ltd.: Middlesex, UK, 1967; 214p. [Google Scholar]
- Holloway, B.A. Fauna of New Zealand. Number 61. Lucanidae (Insecta: Coleoptera); Manaaki Whenua Press: Lincoln, Canterbury, New Zealand, 2007; 254p. [Google Scholar]
- Fujita, H. The Lucanid Beetles of the World; Mushi-Sha: Tokyo, Japan, 2010; 472p. [Google Scholar]
- Kim, S.I.; Farrell, B.D. Phylogeny of world stag beetles (Coleoptera: Lucanidae) reveals a Gondwanan origin of Darwin’s stag beetle. Mol. Phylogenetics Evol. 2015, 86, 35–48. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.D.; Shin, S.; Ahrens, D.; Balke, M.; Beza-Beza, C.; Clarke, D.J.; Donath, A.; Escalona, H.E.; Friedrich, F.; Letsch, H.; et al. The evolution and genomic basis of beetle diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 24729–24737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.Y.; Tihelka, E.; Giacomelli, M.; Lawrence, J.F.; Ślipiński, A.; Kundrata, R.; Yamamoto, S.; Thayer, M.K.; Newton, A.F.; Leschen, R.A.B.; et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. BioRxiv 2021. [Google Scholar] [CrossRef]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolajev, G.V.; Ren, D. A new fossil Lucanidae subfamily (Coleoptera) from the Mesozoic of China. Cauc. Entomol. Bull. 2015, 11, 15–18. [Google Scholar] [CrossRef]
- Qiu, T.F.; Lu, Y.Y.; Zhang, W.W.; Wang, S.; Yang, Y.X.; Bai, M. Electraesalopsis beuteli gen. & sp. nov., the first lucanid beetle from the Cretaceous Burmese amber (Coleoptera: Scarabaeoidea). Zool. Syst. 2017, 42, 390–394. [Google Scholar]
- Cai, C.Y.; Yin, Z.W.; Liu, Y.; Huang, D.Y. Protonicagus tani gen. et sp. nov., the first stag beetles from Upper Cretaceous Burmese amber (Coleoptera: Lucanidae: Aesalinae: Nicagini). Cretac. Res. 2017, 78, 109–112. [Google Scholar] [CrossRef]
- Nikolajev, G.V.; Wang, B.; Liu, Y.; Zhang, H.C. Stag beetles from the Mesozoic of Inner Mongolia, China (Scarabaeoidea: Lucanidae). Acta Palaeontol. Sin. 2011, 50, 41–47. [Google Scholar]
- Bai, M.; Ren, D.; Yang, X.K. Prosinodendron krelli from the Yixian Formation, China: A missing link among Lucanidae, Diphyllostomatidae and Passalidae (Coleoptera: Scarabaeoidea). Cretac. Res. 2012, 34, 334–339. [Google Scholar] [CrossRef]
- Krell, F.T. Catalogue of fossil Scarabaeoidea (Coleoptera: Polyphaga) of the Mesozoic and Tertiary–Version 2007. Denver Mus. Nat. Sci. 2007, 8, 81. [Google Scholar]
- Nikolajev, G.V. Mezozoiskii Etap Evolyutsii Plastinchatousykh (Insecta: Coleoptera: Scarabaeoidea); Kazak Universiteti: Almaty, Kazakhstan, 2007; pp. 17–30. [Google Scholar]
- Chang, M.M. The Jehol Biota: The Emergence of Feathered Dinosaurs, Beaked Birds and Flowering Plants; Shanghai Scientific & Technical Publishers: Shanghai, China, 2003; pp. 1–208. [Google Scholar]
- Swisher, C.C., III; Wang, Y.Q.; Wang, X.L.; Xu, X.; Wang, Y. Cretaceous age for the feathered dinosaurs of Liaoning, China. Nature 1999, 400, 58–61. [Google Scholar] [CrossRef]
- Holloway, B.A. Taxonomy and phylogeny in the Lucanidae (Insecta: Coleoptera). Rec. Dom. Mus. N. Z. 1960, 3, 321–365. [Google Scholar]
- Holloway, B.A. The relationships of Syndesus MacLeay and Sinodendron Schneider (Coleoptera: Lucanidae). N. Z. J. Sci. 1968, 11, 264–269. [Google Scholar]
- Holloway, B.A. Further studies on generic relationships in Lucanidae (Insecta: Coleoptera) with special reference to the ocular canthus. N. Z. J. Sci. 1969, 12, 958–977. [Google Scholar]
- Howden, H.F.; Lawrence, J.F. The New World Aesalinae, with notes on the North American lucanid subfamilies (Coleoptera, Lucanidae). Can. J. Zool. 1974, 52, 1505–1510. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Z.-H.; Tihelka, E.; Cai, C.-Y.; Song, H.-T.; Ai, H.-M. Prolucanus beipiaoensis gen. et sp. nov.: The First Fossil Species of Lucaninae (Coleoptera: Lucanidae) from the Early Cretaceous of Northeastern China. Insects 2022, 13, 272. https://doi.org/10.3390/insects13030272
Qi Z-H, Tihelka E, Cai C-Y, Song H-T, Ai H-M. Prolucanus beipiaoensis gen. et sp. nov.: The First Fossil Species of Lucaninae (Coleoptera: Lucanidae) from the Early Cretaceous of Northeastern China. Insects. 2022; 13(3):272. https://doi.org/10.3390/insects13030272
Chicago/Turabian StyleQi, Zhi-Hao, Erik Tihelka, Chen-Yang Cai, Hai-Tian Song, and Hong-Mu Ai. 2022. "Prolucanus beipiaoensis gen. et sp. nov.: The First Fossil Species of Lucaninae (Coleoptera: Lucanidae) from the Early Cretaceous of Northeastern China" Insects 13, no. 3: 272. https://doi.org/10.3390/insects13030272
APA StyleQi, Z.-H., Tihelka, E., Cai, C.-Y., Song, H.-T., & Ai, H.-M. (2022). Prolucanus beipiaoensis gen. et sp. nov.: The First Fossil Species of Lucaninae (Coleoptera: Lucanidae) from the Early Cretaceous of Northeastern China. Insects, 13(3), 272. https://doi.org/10.3390/insects13030272