
Inter- and Intrasexual Variation in Cuticular Hydrocarbons in Trichrysis cyanea (Linnaeus, 1758) (Hymenoptera: Chrysididae)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Species Determination
2.2. Gas Chromatography—Mass Spectrometry
2.3. DNA Extraction, DNA Barcoding and Genetic Analyses
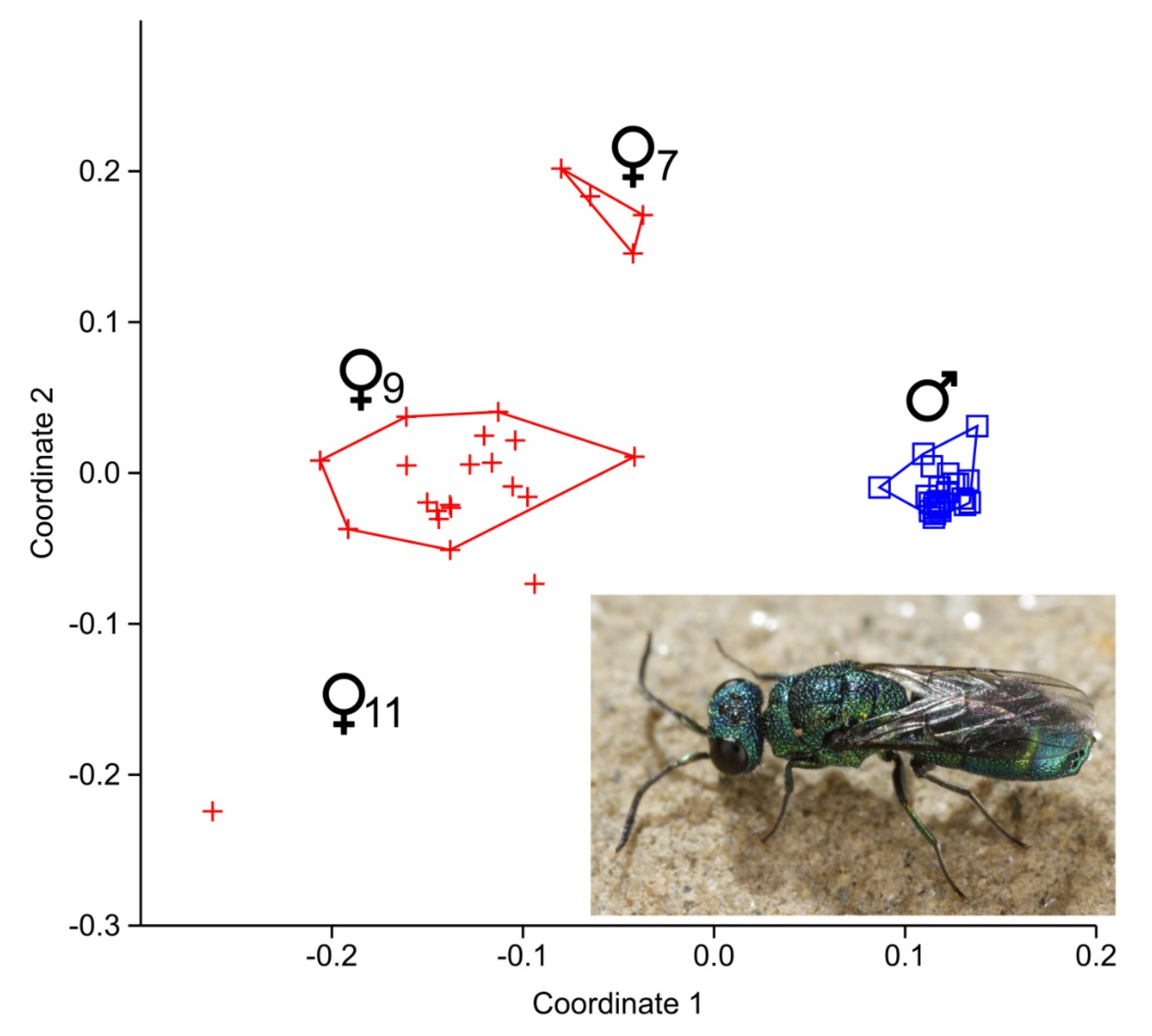
3. Results
3.1. Cuticular Hydrocarbon Patterns (CHCs)
3.2. DNA Barcoding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drijfhout, F.P.; Kather, R.; Martin, S.J. The role of cuticular hydrocarbons in insects. In Behavioral and Chemical Ecology; Zhang, W., Liu, H., Eds.; Nova Science Publishers: New York, NY, USA, 2010; ISBN 1607410990. [Google Scholar]
- Blomquist, G.J.; Bagnères, A.-G. (Eds.) Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-89814-0. [Google Scholar]
- Kather, R.; Martin, S.J. Cuticular hydrocarbon profiles as a taxonomic tool: Advantages, limitations and technical aspects. Physiol. Entomol. 2012, 37, 25–32. [Google Scholar] [CrossRef]
- Hartke, J.; Sprenger, P.P.; Sahm, J.; Winterberg, H.; Orivel, J.; Baur, H.; Beuerle, T.; Schmitt, T.; Feldmeyer, B.; Menzel, F. Cuticular hydrocarbons as potential mediators of cryptic species divergence in a mutualistic ant association. Ecol. Evol. 2019, 9, 9160–9176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soon, V.; Castillo-Cajas, R.F.; Johansson, N.; Paukkunen, J.; Rosa, P.; Ødegaard, F.; Schmitt, T.; Niehuis, O. Cuticular Hydrocarbon Profile Analyses Help Clarify the Species Identity of Dry-Mounted Cuckoo Wasps (Hymenoptera: Chrysididae), Including Type Material, and Reveal Evidence for a Cryptic Species. Insect Syst. Divers. 2021, 5, 3. [Google Scholar] [CrossRef]
- Chung, H.; Carroll, S.B. Wax, sex and the origin of species: Dual roles of insect cuticular hydrocarbons in adaptation and mating. Bioessays 2015, 37, 822–830. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Ginzel, M.D. Chemical Ecology, Biochemistry, and Molecular Biology of Insect Hydrocarbons. Annu. Rev. Entomol. 2021, 66, 45–60. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavioral, and Biochemical Aspects of Insect Hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- De Meeûs, T.; Renaud, F. Parasites within the new phylogeny of eukaryotes. Trends Parasitol. 2002, 18, 247–251. [Google Scholar] [CrossRef]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Colloquium paper: Homage to Linnaeus: How many parasites? How many hosts? Proc. Natl. Acad. Sci. USA 2008, 105 (Suppl. S1), 11482–11489. [Google Scholar] [CrossRef] [Green Version]
- Kilner, R.M.; Langmore, N.E. Cuckoos versus hosts in insects and birds: Adaptations, counter-adaptations and outcomes. Biol. Rev. Camb. Philos. Soc. 2011, 86, 836–852. [Google Scholar] [CrossRef]
- Feeney, W.E.; Welbergen, J.A.; Langmore, N.E. Advances in the Study of Coevolution Between Avian Brood Parasites and Their Hosts. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 227–246. [Google Scholar] [CrossRef]
- Haverkamp, A.; Smid, H.M. A neuronal arms race: The role of learning in parasitoid-host interactions. Curr. Opin. Insect Sci. 2020, 42, 47–54. [Google Scholar] [CrossRef]
- Lasalle, J.; Gauld, I.D. Parasitic Hymenoptera and the biodiversity crisis. REDIA 1991, 74, 315–334. [Google Scholar]
- Pennacchio, F.; Strand, M.R. Evolution of developmental strategies in parasitic hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef]
- Kimsey, L.S.; Bohart, R.M. The Chrysidid Wasps of the World; Oxford University Press: Oxford, UK, 1990; ISBN 0198540108. [Google Scholar]
- Wiesbauer, H.; Rosa, P.; Zettel, H. Die Goldwespen Mitteleuropas: Biologie, Lebensräume, Artenporträts; Ulmer: Stuttgart, Germany, 2020; ISBN 978-3-8186-1149-1. [Google Scholar]
- Wiśniowski, B. Cuckoo-Wasps (Hymenoptera: Chrysididae) of Poland: Diversity, Identification, Distribution; Ojców National Park: Ojców, Poland, 2015; ISBN 978-83-919746-1-2. [Google Scholar]
- Rosa, P. I Crisidi Della Valle d’Aosta: (Hymenoptera, Chrysididae); Museo Regionale di Scienze Naturali: Saint-Pierre, Italy, 2006. [Google Scholar]
- Kroiss, J.; Schmitt, T.; Strohm, E. Low level of cuticular hydrocarbons in a parasitoid of a solitary digger wasp and its potential for concealment. Entomol. Sci. 2009, 12, 9–16. [Google Scholar] [CrossRef]
- Polidori, C.; Ballesteros, Y.; Wurdack, M.; Asís, J.D.; Tormos, J.; Baños-Picón, L.; Schmitt, T. Low Host Specialization in the Cuckoo Wasp, Parnopes grandior, Weakens Chemical Mimicry but Does Not Lead to Local Adaption. Insects 2020, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Strohm, E.; Kroiss, J.; Herzner, G.; Laurien-Kehnen, C.; Boland, W.; Schreier, P.; Schmitt, T. A cuckoo in wolves’ clothing? Chemical mimicry in a specialized cuckoo wasp of the European beewolf (Hymenoptera, Chrysididae and Crabronidae). Front. Zool. 2008, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Wurdack, M.; Herbertz, S.; Dowling, D.; Kroiss, J.; Strohm, E.; Baur, H.; Niehuis, O.; Schmitt, T. Striking cuticular hydrocarbon dimorphism in the mason wasp Odynerus spinipes and its possible evolutionary cause (Hymenoptera: Chrysididae, Vespidae). Proc. Biol. Sci. 2015, 282, 1–9. [Google Scholar] [CrossRef]
- Winterhagen, P. Strategy for sneaking into a host’s home: The cuckoo wasp Omalus biaccinctus (Hymenoptera: Chrysididae) inserts its eggs into living aphids that are the prey of its host. Eur. J. Entomol. 2015, 112, 557–559. [Google Scholar] [CrossRef] [Green Version]
- Pauli, T.; Castillo-Cajas, R.F.; Rosa, P.; Kukowka, S.; Berg, A.; van den Berghe, E.; Fornoff, F.; Hopfenmüller, S.; Niehuis, M.; Peters, R.S.; et al. Phylogenetic analysis of cuckoo wasps (Hymenoptera: Chrysididae) reveals a partially artificial classification at the genus level and a species-rich clade of bee parasitoids. Syst. Entomol. 2019, 44, 322–335. [Google Scholar] [CrossRef]
- Linsenmaier, W. Revision der Familie Chrysididae (Hymenoptera) mit besonderer Berücksichtigung der europäischen Spezies. Mitteilungen der Schweizerischen Entomologischen Gesellschaft 1959, 32, 1–232. [Google Scholar] [CrossRef]
- Bodner, M.; Raspotnig, G. Millipedes that smell like bugs: (E)-alkenals in the defensive secretion of the julid diplopod Allajulus dicentrus. J. Chem. Ecol. 2012, 38, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.A.; Roan, C.S.; Yost, R.A.; Hector, J. Dimethyl disulfide derivatives of long chain alkenes, alkadienes, and alkatrienes for gas chromatography/mass spectrometry. Anal. Chem. 1989, 61, 1564–1571. [Google Scholar] [CrossRef]
- Van den Dool, H.; Dec Kratz, P. A generalization of the retention index system including linear temperature programmed gas—Liquid partition chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Richlen, M.L.; Barber, P.H. A technique for the rapid extraction of microalgal DNA from single live and preserved cells. Mol. Ecol. Notes 2005, 5, 688–691. [Google Scholar] [CrossRef]
- Zangl, L.; Kunz, G.; Berg, C.; Koblmüller, S. First records of the parthenogenetic Surinam cockroach Pycnoscelus surinamensis (Insecta: Blattodea: Blaberidae) for Central Europe. J. Appl. Entomol. 2019, 143, 308–313. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Aalysis Using Maximum Likelihood, Evolutionary Eistance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Moris, V.C.; Christmann, K.; Wirtgen, A.; Belokobylskij, S.A.; Berg, A.; Liebig, W.-H.; Soon, V.; Baur, H.; Schmitt, T.; Niehuis, O. Cuticular hydrocarbons on old museum specimens of the spiny mason wasp, Odynerus spinipes (Hymenoptera: Vespidae: Eumeninae), shed light on the distribution and on regional frequencies of distinct chemotypes. Chemoecology 2021, 31, 311–322. [Google Scholar] [CrossRef]
- Crozier, R.H.; Crozier, Y.C.; Mackinlay, A.G. The CO-I and CO-II Region of Honeybee Mitochondrial DNA: Evidence for Variation in Insect Mitochondrial Evolutionary Rates. Mol. Biol. Evol. 1989, 6, 399–411. [Google Scholar] [CrossRef]
- Kaltenpoth, M.; Showers Corneli, P.; Dunn, D.M.; Weiss, R.B.; Strohm, E.; Seger, J. Accelerated Evolution of Mitochondrial but Not Nuclear Genomes of Hymenoptera: New Evidence from Crabronid Wasps. PLoS ONE 2012, 7, e32826. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.C.S.G.; Raychoudhury, R.; Lavrov, D.V.; Werren, J.H. Rapidly Evolving Mitochondrial Genome and Directional Selection in Mitochondrial Genes in the Parasitic Wasp Nasonia (Hymenoptera: Pteromalidae). Mol. Biol. Evol. 2008, 25, 2167–2180. [Google Scholar] [CrossRef] [Green Version]
- Turčinavičienė, J.; Radzevičiūtė, R.; Budrienė, A.; Budrys, E. Species identification and genetic differentiation of European cavity-nesting wasps (Hymenoptera: Vespidae, Pompilidae, Crabronidae) inferred from DNA barcoding data. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 476–482. [Google Scholar] [CrossRef]
- Butterworth, N.J.; Drijfhout, F.P.; Byrne, P.G.; Keller, P.A.; Wallman, J.F. Major Transitions in Cuticular Hydrocarbon Expression Coincide with Sexual Maturity in a Blowfly (Diptera: Calliphoridae). J. Chem. Ecol. 2020, 46, 610–618. [Google Scholar] [CrossRef]
- Amsalem, E.; Hefetz, A. The appeasement effect of sterility signaling in dominance contests among Bombus terrestris workers. Behav. Ecol. Sociobiol. 2010, 64, 1685–1694. [Google Scholar] [CrossRef]
- Peeters, C.; Monnin, T.; Malosse, C. Cuticular hydrocarbons correlated with reproductive status in a queenless ant. Proc. R. Soc. Lond. B 1999, 266, 1323–1327. [Google Scholar] [CrossRef] [Green Version]
- Everaerts, C.; Farine, J.-P.; Cobb, M.; Ferveur, J.-F. Drosophila cuticular hydrocarbons revisited: Mating status alters cuticular profiles. PLoS ONE 2010, 5, e9607. [Google Scholar] [CrossRef]
- Arienti, M.; Antony, C.; Wicker-Thomas, C.; Delbecque, J.-P.; Jallon, J.-M. Ontogeny of Drosophila melanogaster female sex-appeal and cuticular hydrocarbons. Integr. Zool. 2010, 5, 272–282. [Google Scholar] [CrossRef]
- Kather, R.; Martin, S.J. Evolution of Cuticular Hydrocarbons in the Hymenoptera: A Meta-Analysis. J. Chem. Ecol. 2015, 41, 871–883. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Name | Retention Index | Abbreviation |
|---|---|---|
| 9- and 10-Heneicosene | 2076 | 9/(10)-C21:1 1 |
| 7-Heneicosene | 2082 | 7-C21:1 |
| Heneicosane | 2100 | C21 |
| 11-Tricosene | 2274 | 11-C23:1 |
| 9-Tricosene | 2277 | 9-C23:1 |
| 7-Tricosene | 2284 | 7-C23:1 |
| Tricosane | 2301 | C23 |
| 11-Pentacosene | 2472 | 11-C25:1 |
| 9-Pentacosene | 2477 | 9-C25:1 |
| 7-Pentacosene | 2484 | 7-C25:1 |
| Pentacosane | 2500 | C25 |
| 13-Heptacosene | 2670 | 13-C27:1 |
| 11-Heptacosene | 2671 | 11-C27:1 |
| 9-Heptacosene | 2677 | 9-C27:1 |
| 7-Heptacosene | 2685 | 7-C27:1 |
| Heptacosane | 2700 | C27 |
| 13- and 14-Nonacosene | 2868 | 13/14-C29:1 2 |
| 11-Nonacosene | 2871 | 11-C29:1 |
| 9-Nonacosene | 2878 | 9-C29:1 |
| 7-Nonacosene | 2886 | 7-C29:1 |
| Nonacosane | 2900 | C29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fröhlich, D.; Zangl, L.; Raspotnig, G.; Koblmüller, S. Inter- and Intrasexual Variation in Cuticular Hydrocarbons in Trichrysis cyanea (Linnaeus, 1758) (Hymenoptera: Chrysididae). Insects 2022, 13, 159. https://doi.org/10.3390/insects13020159
Fröhlich D, Zangl L, Raspotnig G, Koblmüller S. Inter- and Intrasexual Variation in Cuticular Hydrocarbons in Trichrysis cyanea (Linnaeus, 1758) (Hymenoptera: Chrysididae). Insects. 2022; 13(2):159. https://doi.org/10.3390/insects13020159
Chicago/Turabian StyleFröhlich, David, Lukas Zangl, Günther Raspotnig, and Stephan Koblmüller. 2022. "Inter- and Intrasexual Variation in Cuticular Hydrocarbons in Trichrysis cyanea (Linnaeus, 1758) (Hymenoptera: Chrysididae)" Insects 13, no. 2: 159. https://doi.org/10.3390/insects13020159
APA StyleFröhlich, D., Zangl, L., Raspotnig, G., & Koblmüller, S. (2022). Inter- and Intrasexual Variation in Cuticular Hydrocarbons in Trichrysis cyanea (Linnaeus, 1758) (Hymenoptera: Chrysididae). Insects, 13(2), 159. https://doi.org/10.3390/insects13020159