
Identification and Functional Characterization of Toxoneuron nigriceps Ovarian Proteins Involved in the Early Suppression of Host Immune Response

 ,
,  ,
,  ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Calyx Fluid Collection, Ovarian Protein Purification and RNA Extraction
2.3. Toxoneuron nigriceps Protein Database Building
2.4. HPLC Analysis of the Ovarian Proteins and Transfer on the PVDF Membrane
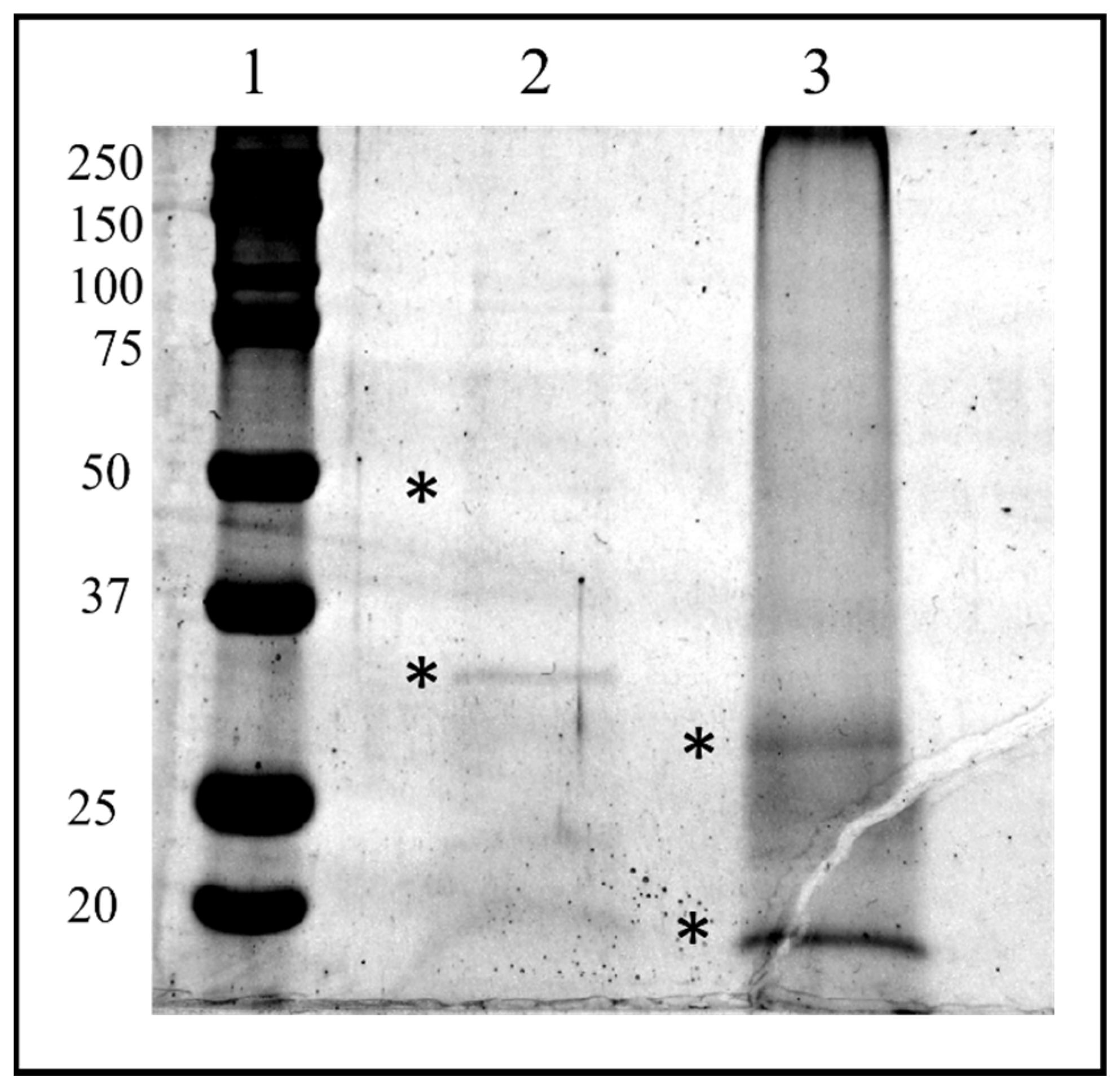
2.5. SDS-PAGE and In Situ Protein Digestion
2.6. LC–MS/MS and Protein Identification
2.7. Collection of Hemocytes from Larvae of H. virescens
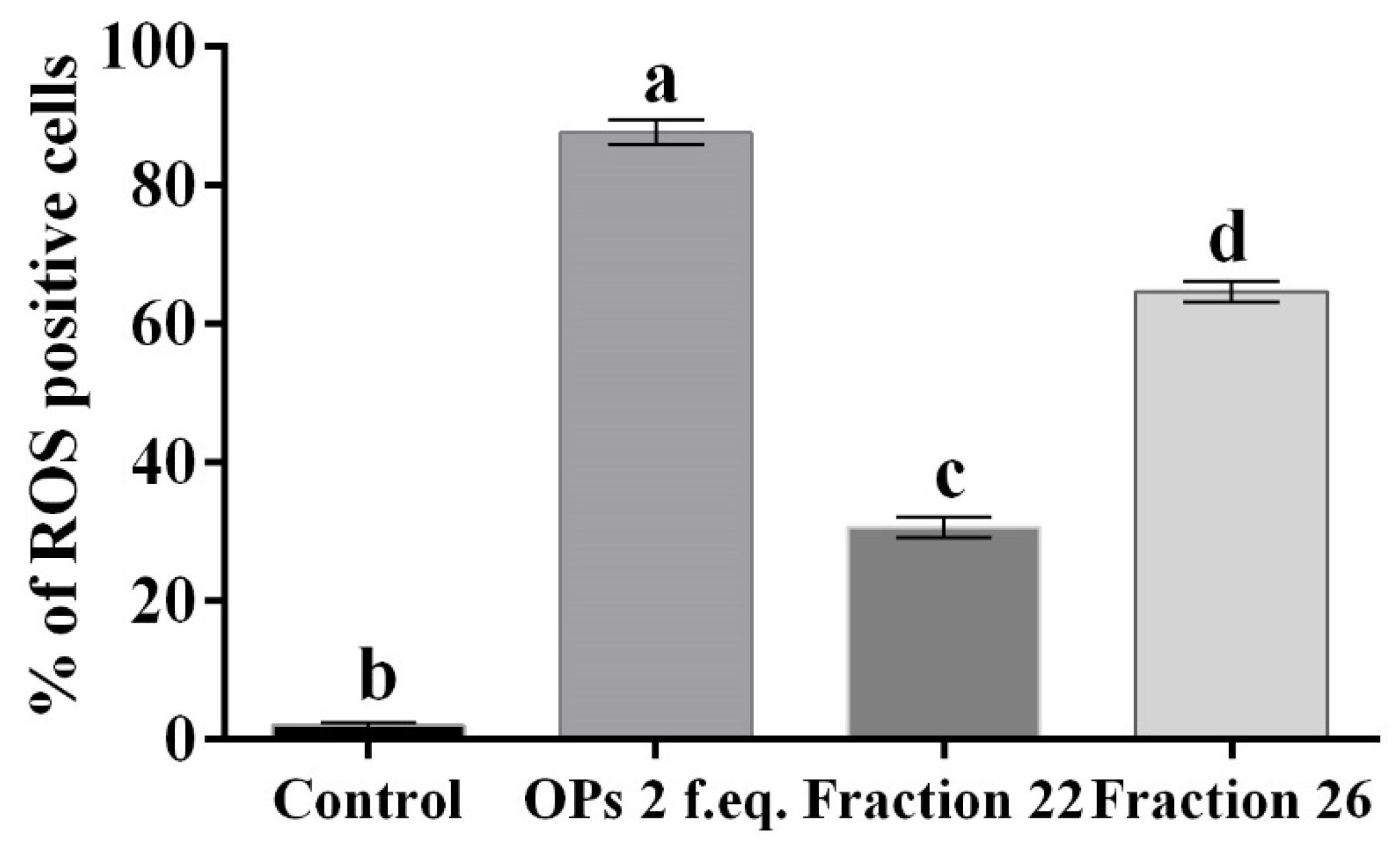
2.8. Cells Viability
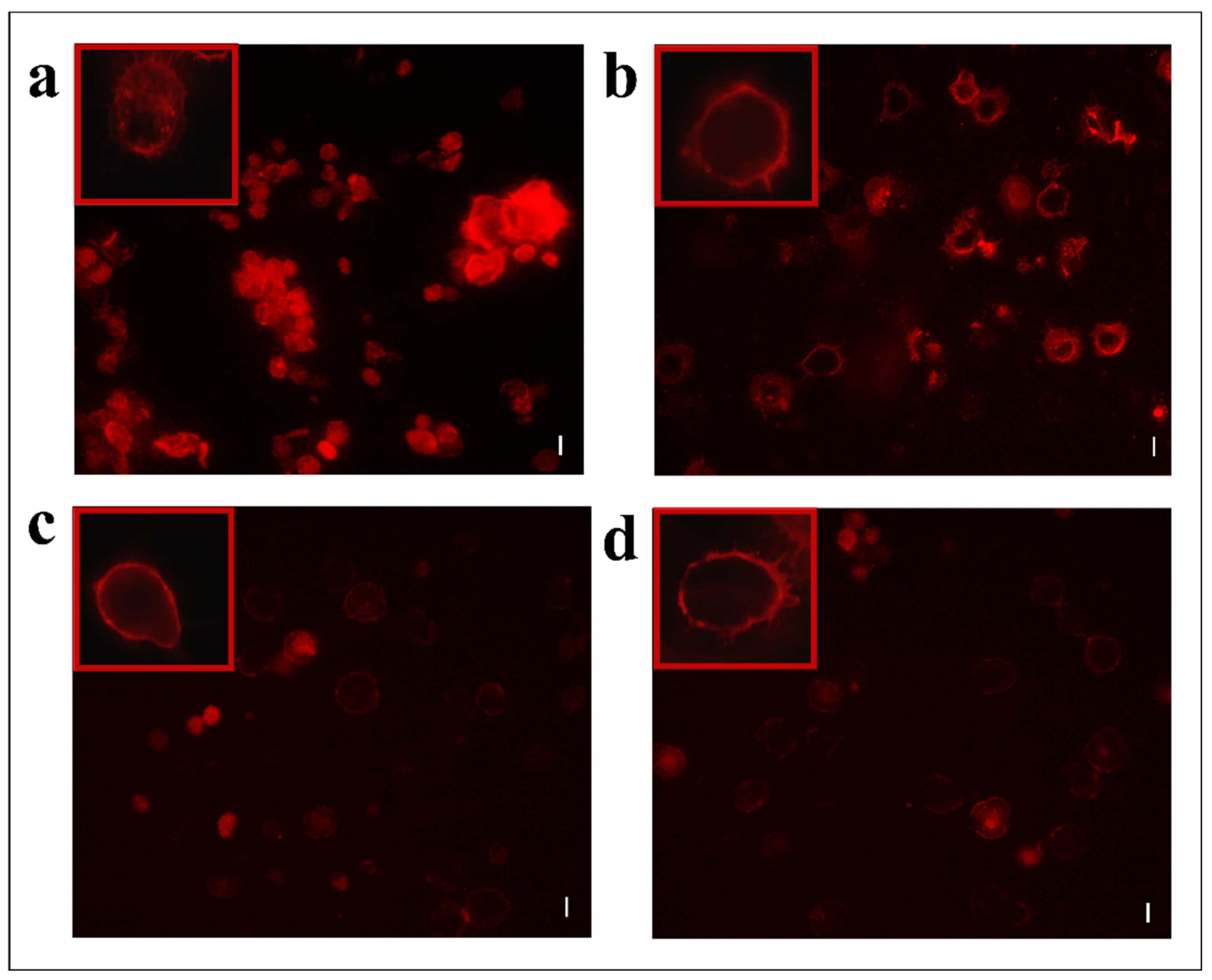
2.9. Light Microscopy Hemocyte Observations
2.10. Encapsulation Assay
2.11. Statistical Analysis of Data
3. Results
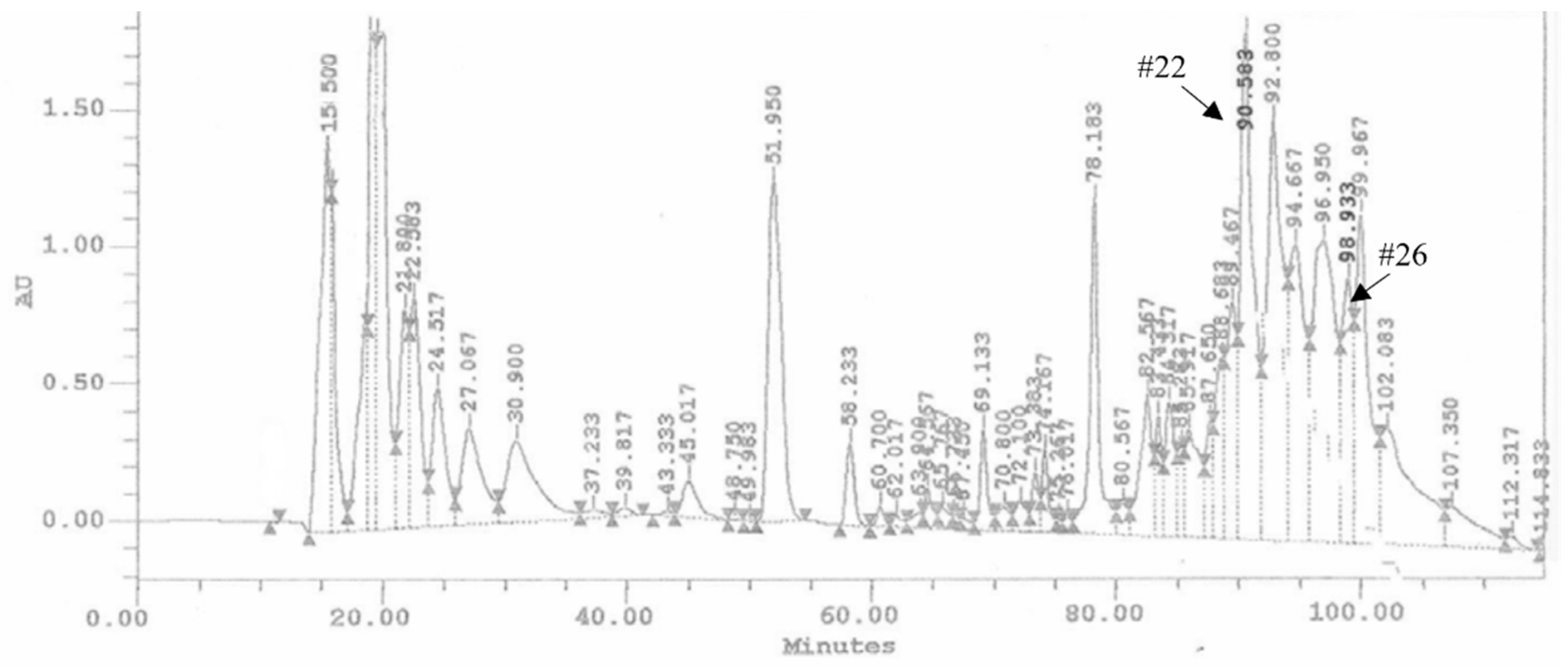
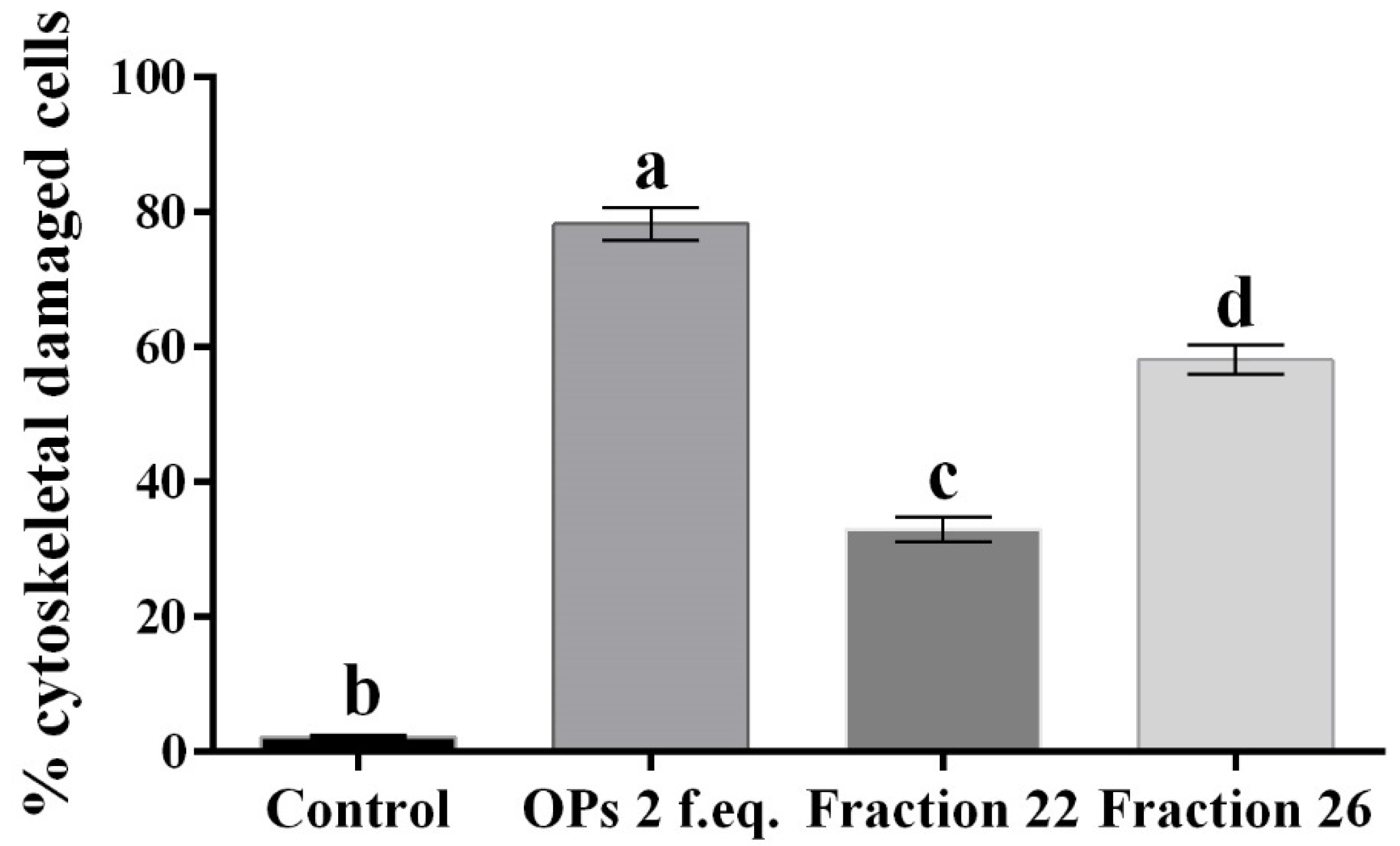
3.1. HPLC Fractions of Ovarian Proteins and Evaluation of Their Activity
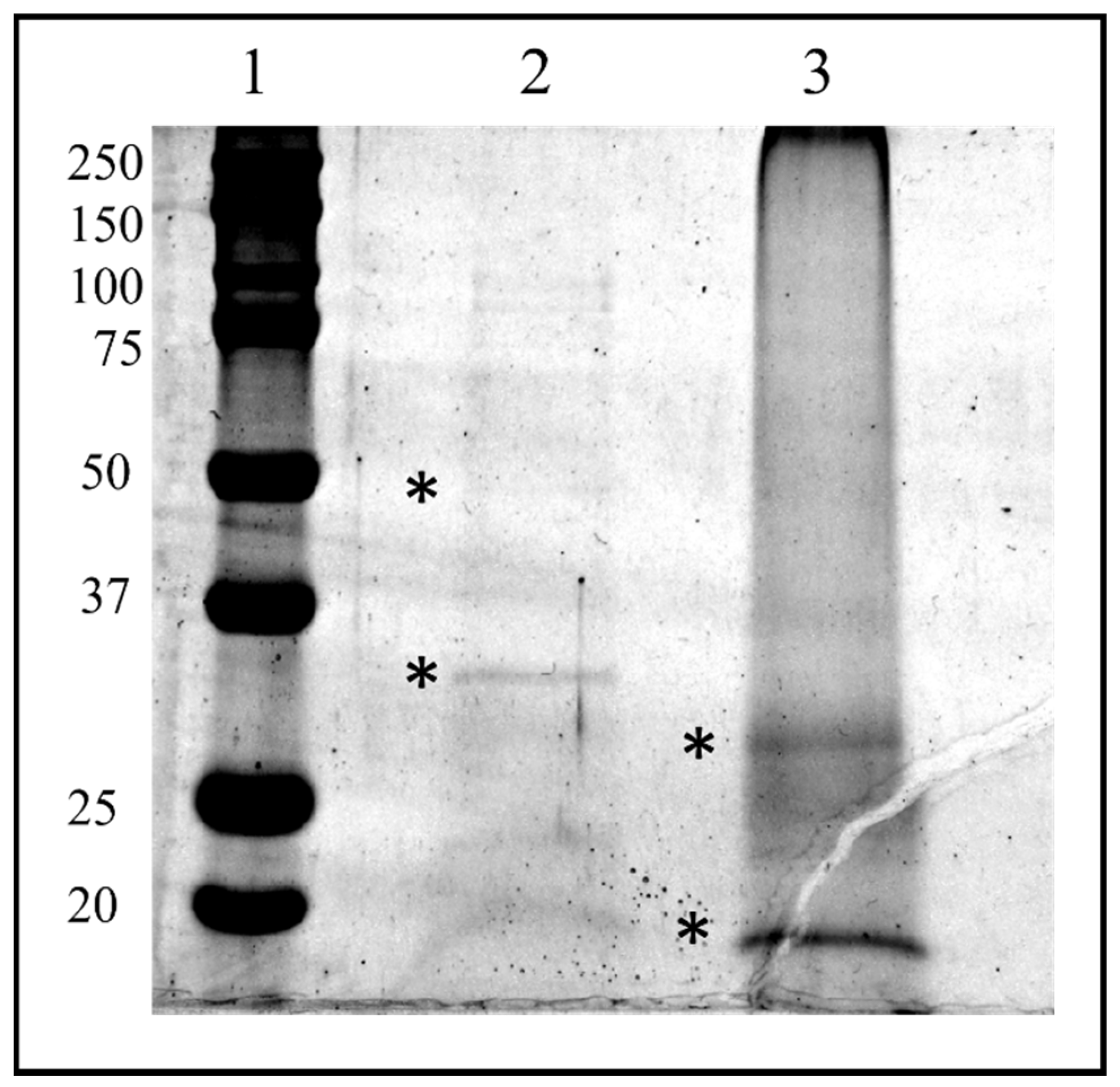
3.2. Identification of Proteins in HPLC Fraction #22 and #26
4. Discussion
- –
- Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (36 kDa, contig T_C201). It is a well-known key enzyme in glycolysis that catalyzes the first step of the pathway by converting D-glyceraldehyde 3-phosphate (G3P) into 3-phospho-D-glyceroyl phosphate [51,52]. However, it is reported in many novel cellular roles including apoptosis, tRNA export, and receptor-associated kinase [53,54,55].
- –
- Phosphoglycerate mutase (PGAM) [56] (30 kDa, contig T_C2966) is involved in metabolism, in particular it catalyzes the reversible reaction of 3-phosphoglycerate (3-PGA) to 2-phosphoglycerate (2-PGA) in the glycolytic pathway. It is reported that in mutant mice that overexpressed Pgam2, the reactive oxygen species (ROS) was increased [57].
- –
- Glutathione transferase (GST) (24.5 kDa, contig T_C2043) [58,59] is an enzyme that catalyzes the conjugation of glutathione (GSH) to a variety of electrophilic substances, but GST has also been shown to act as modulator of signal transduction pathways that control cell proliferation and cell death, modulating several signaling cascades [58].
- –
- Proliferating cell nuclear antigen (PCNA) (26 kDa, contig T_C349), a cell cycle marker protein [60]. It is an essential component for eukaryotic chromosomal DNA replication and repair. The recent proteomics approaches showed that PCNA interacts with more than 100 PCNA-interacting proteins, indicating the role of PCNA in several cellular functions. Among these, it could have a possible role in apoptosis; indeed, it has been shown that apoptotic cells expressed high levels of PCNA [61].
- –
- Apolipophorin-III (23.5 kDa, contig T_C1034, annotated as “uncharacterized protein” with a conserved domain of Apolipophorin-III superfamily) is involved in the transport of lipids [62]. However, it has been reported that in Galleria mellonella it plays a key role in immune response against bacteria, both Gram-negative and Gram-positive, fungi and yeasts; indeed, in G. mellonella larvae after immunization with Gram-negative bacteria Escherichia coli, Gram-positive bacteria Micrococcus luteus, yeast Candida albicans, and the filamentous fungus Fusarium oxysporum, the presence of this protein increased in the hemolymph, hemocytes, and fat body, enhancing the activity of antibacterial peptide such as cecropin [63,64]. The presence of this protein among the OPs could be easily explained indeed, if, on the one hand, the maternal parasitoids factors must inhibit the immune response against the parasitoid, on the other hand, they must guarantee the survival of the host, preventing the attack by other pathogens.
- –
- Cu/Zn−superoxide dismutase (SOD1) (16.5 kDa, contig T_C1185), found in the fraction #26, could modulate the physiology of the H. virescens. It is a ubiquitous enzyme that catalyzes the dismutation of superoxide radicals to oxygen and hydrogen peroxide [65]. Several oxidoreductases have been found in the venom of parasitoid insects, including T. nigriceps, but its role in parasitization is still unknown. It could be hypothesized that SOD1 could prevent the pupation since a recent study reported that ROS production and downregulation of superoxide dismutase are required for pupation in Bombyx mori [66].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moreau, S.; Asgari, S. Venom Proteins from Parasitoid Wasps and Their Biological Functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazi, E.M.; Abol Ella, S.M.; Bazzaz, A.; Khamis, O.; Abo Abd-Allah, L.M.Z. The calyx fluid of Microplitis rufiventris parasitoid and growth of its host Spodoptera littoralis larvae. J. Insect Physiol. 2005, 51, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Falabella, P. The mechanism utilized by Toxoneuron nigriceps in inhibiting the host immune system. Invert. Surviv. J. 2018, 15, 240–255. [Google Scholar]
- Heraty, J. Parasitoid Biodiversity and Insect Pest Management. In Insect Biodiversity; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 603–625. [Google Scholar]
- Schmidt, O.; Theopold, U.; Strand, M. Innate immunity and its evasion and suppression by hymenopteran endoparasitoids. BioEssays 2001, 23, 344–351. [Google Scholar] [CrossRef]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Scieuzo, C.; Salvia, R.; Franco, A.; Pezzi, M.; Cozzolino, F.; Chicca, M.; Scapoli, C.; Vogel, H.; Monti, M.; Ferracini, C.; et al. An integrated transcriptomic and proteomic approach to identify the main Torymus sinensis venom components. Sci. Rep. 2021, 11, 5032. [Google Scholar] [CrossRef]
- Nakamatsu, Y.; Tanaka, T. Venom of ectoparasitoid, Euplectrus sp. near plathypenae (Hymenoptera: Eulophidae) regulates the physiological state of Pseudaletia separata (Lepidoptera: Noctuidae) host as a food resource. J. Insect Physiol. 2003, 49, 149–159. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, J.-Y.; Qian, C.; Fang, Q.; Ye, G.-Y. Venom of The Parasitoid Wasp Pteromalus Puparum Contains an Odorant Binding Protein. Arch. Insect Biochem. Physiol. 2015, 88, 101–110. [Google Scholar] [CrossRef]
- Sim, A.D.; Wheeler, D. The venom gland transcriptome of the parasitoid wasp Nasonia vitripennis highlights the importance of novel genes in venom function. BMC Genom. 2016, 17, 571. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Chen, Y.-F.; Chen, X.; Huang, F.; Lou, Y.; Liu, S. Effects of venom/calyx fluid from the endoparasitic wasp Cotesia plutellae on the hemocytes of its host Plutella xylostella in vitro. J. Insect Physiol. 2007, 53, 22–29. [Google Scholar] [CrossRef]
- Li, Y.; Lu, J.-F.; Feng, C.-J.; Ke, X.; Fu, W.-J. Role of venom and ovarian proteins in immune suppression of Ostrinia furnacalis (Lepidoptera: Pyralidae) larvae parasitized by Macrocentrus cingulum (Hymenoptera: Braconidae), a polyembryonic parasitoid. Insect Sci. 2007, 14, 93–100. [Google Scholar] [CrossRef]
- Laurino, S.; Grossi, G.; Pucci, P.; Flagiello, A.; Bufo, S.A.; Bianco, G.; Salvia, R.; Vinson, S.B.; Vogel, H.; Falabella, P. Identification of major Toxoneuron nigriceps venom proteins using an integrated transcriptomic/proteomic approach. Insect Biochem. Mol. Biol. 2016, 76, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Pennacchio, F.; Strand, M.R. Evolution of Developmental Strategies in Parasitic Hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Digilio, M.C.; Isidoro, N.; Tremblay, E.; Pennacchio, F. Host castration by Aphidius ervi venom proteins. J. Insect Physiol. 2000, 46, 1041–1050. [Google Scholar] [CrossRef]
- Falabella, P.; Riviello, L.; Caccialupi, P.; Rossodivita, T.; Teresa Valente, M.; Luisa De Stradis, M.; Tranfaglia, A.; Varricchio, P.; Gigliotti, S.; Graziani, F. A γ-glutamyl transpeptidase of Aphidius ervi venom induces apoptosis in the ovaries of host aphids. Insect Biochem. Mol. Biol. 2007, 37, 453–465. [Google Scholar] [CrossRef]
- Pennacchio, F.; Mancini, D. Aphid Parasitoid Venom and its Role in Host Regulation. In Parasitoid Viruses; Elsevier: Amsterdam, The Netherlands, 2012; pp. 247–254. [Google Scholar]
- Webb, B.A.; Luckhart, S. Factors mediating short- and long-term immune suppression in a parasitized insect. J. Insect Physiol. 1996, 42, 33–40. [Google Scholar] [CrossRef]
- Webb, B.A.; Luckhart, S. Evidence for an early immunosuppressive role for related Campoletis sonorensis venom and ovarian proteins in Heliothis virescens. Arch. Insect Biochem. Physiol. 1994, 26, 147–163. [Google Scholar] [CrossRef]
- Salvia, R.; Scieuzo, C.; Grimaldi, A.; Fanti, P.; Moretta, A.; Franco, A.; Varricchio, P.; Vinson, S.B.; Falabella, P. Role of Ovarian Proteins Secreted by Toxoneuron nigriceps (Viereck) (Hymenoptera, Braconidae) in the Early Suppression of Host Immune Response. Insects 2021, 12, 33. [Google Scholar] [CrossRef]
- Pech, L.L.; Strand, M.R. Granular cells are required for encapsulation of foreign targets by insect haemocytes. J. Cell Sci. 1996, 109, 2053–2060. [Google Scholar] [CrossRef]
- Loret, S.M.; Strand, M.R. Follow-up of protein release from Pseudoplusia includens hemocytes: A first step toward identification of factors mediating encapsulation in insects. Eur. J. Cell Biol. 1998, 76, 146–155. [Google Scholar] [CrossRef]
- Edson, K.M.; Vinson, S.B.; Stoltz, D.B.; Summers, M.D. Virus in a Parasitoid Wasp: Suppression of the Cellular Immune Response in the Parasitoid’s Host. Science 1981, 211, 582–583. [Google Scholar] [CrossRef] [PubMed]
- Luckhart, S.; Webb, B.A. Interaction of a wasp ovarian protein and polydnavirus in host immune suppression. Dev. Comp. Immunol. 1996, 20, 1–21. [Google Scholar] [CrossRef]
- Krell, P.J.; Summers, M.D.; Vinson, S.B. Virus with a Multipartite Superhelical DNA Genome from the Ichneumonid Parasitoid Campoletis sonorensis. J. Virol. 1982, 43, 859–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoltz, D.B. The polydnavirus life cycle. In Parasites and Pathogens of Insects; Beckage, N.E., Thompson, S.N., Federici, B.A., Eds.; Academic Press: New York, NY, USA, 1993; pp. 167–187. [Google Scholar]
- Salvia, R.; Grossi, G.; Amoresano, A.; Scieuzo, C.; Nardiello, M.; Giangrande, C.; Laurenzana, I.; Ruggieri, V.; Bufo, S.A.; Vinson, S.B.; et al. The multifunctional polydnavirus TnBVANK1 protein: Impact on host apoptotic pathway. Sci. Rep. 2017, 7, 11775. [Google Scholar] [CrossRef] [PubMed]
- Theilmann, D.A.; Summers, M.D. Molecular Analysis of Campoletis sonorensis Virus DNA in the Lepidopteran Host Heliothis virescens. J. Gen. Virol. 1986, 67, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.A.; Cui, L. Relationships between polydnavirus genomes and viral gene expression. J. Insect Physiol. 1998, 44, 785–793. [Google Scholar] [CrossRef]
- Lavine, M.D.; Beckage, N.E. Polydnaviruses: Potent mediators of host insect immune dysfunction. Parasitol. Today 1995, 11, 368–378. [Google Scholar] [CrossRef]
- Strand, M.R.; Pech, L.L. Immunological Basis for Compatibility in Parasitoid-Host Relationships. Annu. Rev. Entomol. 1995, 40, 31–56. [Google Scholar] [CrossRef]
- Salvia, R.; Nardiello, M.; Scieuzo, C.; Scala, A.; Bufo, S.A.; Rao, A.; Vogel, H.; Falabella, P. Novel Factors of Viral Origin Inhibit TOR Pathway Gene Expression. Front. Physiol. 2018, 9, 1678. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.R.; Burke, G.R. Polydnaviruses: Nature’s Genetic Engineers. Annu. Rev. Virol. 2014, 1, 333–354. [Google Scholar] [CrossRef]
- Edwards, J.P.; Weaver, R.J. Endocrine Interactions of Insect Parasites and Pathogens; BIOS Scientific Publishers: Oxford, UK, 2001. [Google Scholar]
- Salvia, R.; Grimaldi, A.; Girardello, R.; Scieuzo, C.; Scala, A.; Bufo, S.A.; Vogel, H.; Falabella, P. Aphidius ervi Teratocytes Release Enolase and Fatty Acid Binding Protein Through Exosomal Vesicles. Front. Physiol. 2019, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- de Buron, I.; Beckage, N.E. Developmental changes in teratocytes of the braconid wasp Cotesia congregata in larvae of the tobacco hornworm, Manduca sexta. J. Insect Physiol. 1997, 43, 915–930. [Google Scholar] [CrossRef]
- Vinson, S.B.; Guillot, F.S.; Hays, D.B. Rearing of Cardiochiles nigriceps in the Laboratory, with Heliothis virescens as Hosts. Ann. Entomol. Soc. Am. 1973, 66, 1170–1172. [Google Scholar] [CrossRef]
- Vanderzant, E.S.; Richardson, C.D.; Fort, S.W., Jr. Rearing of the bollworm on artificial diet. J. Econ. Entomol. 1962, 55, 140. [Google Scholar] [CrossRef]
- Troise, F.; Monti, M.; Merlino, A.; Cozzolino, F.; Fedele, C.; Russo Krauss, I.; Sica, F.; Pucci, P.; D’Alessio, G.; De Lorenzo, C. A novel ErbB2 epitope targeted by human antitumor immunoagents. FEBS J. 2011, 278, 1156–1166. [Google Scholar] [CrossRef]
- Fusco, S.; Aulitto, M.; Iacobucci, I.; Crocamo, G.; Pucci, P.; Bartolucci, S.; Monti, M.; Contursi, P. The interaction between the F55 virus-encoded transcription regulator and the RadA host recombinase reveals a common strategy in Archaea and Bacteria to sense the UV-induced damage to the host DNA. Biochim. Biophys. Acta-Gene Regul. Mech. 2020, 1863, 194493. [Google Scholar] [CrossRef]
- Ferrarese, R.; Brivio, M.; Congiu, T.; Falabella, P.; Grimaldi, A.; Mastore, M.; Perletti, G.; Sciacca, L.; Tettamanti, G.; Valvassori, R.; et al. Early suppression of immune response in Heliothis virescens larvae by the endophagous parasitoid Toxoneuron nigriceps. Invertebr. Surv. J. 2005, 2, 60–68. [Google Scholar]
- Shubin, A.V.; Demidyuk, I.V.; Komissarov, A.A.; Rafieva, L.M.; Kostrov, S.V. Cytoplasmic vacuolization in cell death and survival. Oncotarget 2016, 7, 55863–55889. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, D.M.; Lichtveld, M.; Mazet, J.A.K.; Togami, E.; Miller, S.A. Plant health and its effects on food safety and security in a One Health framework: Four case studies. One Health Outlook 2021, 3, 6. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Rome. 2017. Available online: https://www.fao.org/3/i6583e/i6583e.pdf (accessed on 22 December 2021).
- Becchimanzi, A.; Avolio, M.; Bostan, H.; Colantuono, C.; Cozzolino, F.; Mancini, D.; Chiusano, M.L.; Pucci, P.; Caccia, S.; Pennacchio, F. Venomics of the ectoparasitoid wasp Bracon nigricans. BMC Genom. 2020, 21, 34. [Google Scholar] [CrossRef]
- Ohkuma, S.; Poole, B. Cytoplasmic vacuolation of mouse peritoneal macrophages and the uptake into lysosomes of weakly basic substances. J. Cell Biol. 1981, 90, 656–664. [Google Scholar] [CrossRef]
- Scieuzo, C.; Nardiello, M.; Salvia, R.; Pezzi, M.; Chicca, M.; Leis, M.; Bufo, S.A.; Vinson, S.B.; Rao, A.; Vogel, H.; et al. Ecdysteroidogenesis and development in Heliothis virescens (Lepidoptera: Noctuidae): Focus on PTTH-stimulated pathways. J. Insect Physiol. 2018, 107, 57–67. [Google Scholar] [CrossRef]
- Lubec, G.; Afjehi-Sadat, L.; Yang, J.W.; John, J.P. Searching for hypothetical proteins: Theory and practice based upon original data and literature. Prog. Neurobiol. 2005, 77, 90–127. [Google Scholar] [CrossRef]
- di Pietro, F.; Echard, A.; Morin, X. Regulation of mitotic spindle orientation: An integrated view. EMBO Rep. 2016, 17, 1106–1130. [Google Scholar] [CrossRef] [Green Version]
- Kabakov, A.E.; Budagova, K.R.; Bryantsev, A.L.; Latchman, D.S. Heat shock protein 70 or heat shock protein 27 overexpressed in human endothelial cells during posthypoxic reoxygenation can protect from delayed apoptosis. Cell Stress Chaperones 2003, 8, 335–347. [Google Scholar] [CrossRef] [Green Version]
- Pennington, K.; Chan, T.; Torres, M.; Andersen, J. The dynamic and stress-adaptive signaling hub of 14-3-3: Emerging mechanisms of regulation and context-dependent protein–protein interactions. Oncogene 2018, 37, 5587–5604. [Google Scholar] [CrossRef] [Green Version]
- Chong, I.; Ho, W. Glyceraldehyde-3-phosphate Dehydrogenase from Chironomidae Showed Differential Activity towards Metals. Protein Pept. Lett. 2013, 20, 970–976. [Google Scholar] [CrossRef]
- Chuang, D.-M.; Hough, C.; Senatorov, V.V. Glyceraldehyde-3-Phosphate Dehydrogenase, Apoptosis, And Neurodegenerative Diseases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 269–290. [Google Scholar] [CrossRef]
- Singh, R.; Green, M.R. Sequence-Specific Binding of Transfer RNA by Glyceraldehyde-3-Phosphate Dehydrogenase. Science 1993, 259, 365–368. [Google Scholar] [CrossRef]
- Tarze, A.; Deniaud, A.; Le Bras, M.; Maillier, E.; Molle, D.; Larochette, N.; Zamzami, N.; Jan, G.; Kroemer, G.; Brenner, C. GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene 2007, 26, 2606–2620. [Google Scholar] [CrossRef] [Green Version]
- Jedrzejas, M.J.; Chander, M.; Setlow, P.; Krishnasamy, G. Mechanism of catalysis of the cofactor-independent phosphoglycerate mutase from Bacillus stearothermophilus. Crystal structure of the complex with 2-phosphoglycerate. J. Biol. Chem. 2000, 275, 23146–23153. [Google Scholar] [CrossRef] [Green Version]
- Okuda, J.; Niizuma, S.; Shioi, T.; Kato, T.; Inuzuka, Y.; Kawashima, T.; Tamaki, Y.; Kawamoto, A.; Tanada, Y.; Iwanaga, Y.; et al. Persistent Overexpression of Phosphoglycerate Mutase, a Glycolytic Enzyme, Modifies Energy Metabolism and Reduces Stress Resistance of Heart in Mice. PLoS ONE 2013, 8, e72173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tew, K.D.; Townsend, D.M. Glutathione-S-Transferases as Determinants of Cell Survival and Death. Antioxid. Redox Signal. 2012, 17, 1728–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborde, E. Glutathione transferases as mediators of signaling pathways involved in cell proliferation and cell death. Cell Death Differ. 2010, 17, 1373–1380. [Google Scholar] [CrossRef] [Green Version]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, R.; Misund, K.; Holien, T.; Bachke, S.; Gilljam, K.M.; Våtsveen, T.K.; Rø, T.B.; Bellacchio, E.; Sundan, A.; Otterlei, M. Targeting proliferating cell nuclear antigen and its protein interactions induces apoptosis in multiple myeloma cells. PLoS ONE 2013, 8, e70430. [Google Scholar] [CrossRef] [Green Version]
- van der Horst, D.J.; van Hoof, D.; van Marrewijk, W.J.A.; Rodenburg, K.W. Alternative lipid mobilization: The insect shuttle system. Mol. Cell. Biochem. 2002, 239, 113–119. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, C.H.; Jeong, W.H.; Lee, J.H.; Seo, S.J.; Han, Y.S.; Lee, I.H. Effects of two hemolymph proteins on humoral defense reactions in the wax moth, Galleria mellonella. Dev. Comp. Immunol. 2005, 29, 43–51. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Sowa-Jasiłek, A.; Stączek, S.; Jakubowicz, T.; Cytryńska, M. Different forms of apolipophorin III in Galleria mellonella larvae challenged with bacteria and fungi. Peptides 2015, 68, 105–112. [Google Scholar] [CrossRef]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. The copper-zinc superoxide dismutase activity in selected diseases. Eur. J. Clin. Investig. 2019, 49, e13036. [Google Scholar] [CrossRef] [Green Version]
- Nojima, Y.; Bono, H.; Yokoyama, T.; Iwabuchi, K.; Sato, R.; Arai, K.; Tabunoki, H. Superoxide dismutase down-regulation and the oxidative stress is required to initiate pupation in Bombyx mori. Sci. Rep. 2019, 9, 14693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falabella, P.; Varricchio, P.; Gigliotti, S.; Tranfaglia, A.; Pennacchio, F.; Malva, C. Toxoneuron nigriceps polydnavirus encodes a putative aspartyl protease highly expressed in parasitized host larvae. Insect Mol. Biol. 2003, 12, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, R.; Wilson, R.; Vilaplana, L.; O’Reilly, D.R.; Falabella, P.; Douris, V.; Bernier-Cardou, M.; Pennacchio, F.; Iatrou, K.; Malva, C.; et al. Expression of a Toxoneuron nigriceps polydnavirus-encoded protein causes apoptosis-like programmed cell death in lepidopteran insect cells. J. Gen. Virol. 2005, 86, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Falabella, P.; Caccialupi, P.; Varricchio, P.; Malva, C.; Pennacchio, F. Protein tyrosine phosphatases of Toxoneuron nigriceps bracovirus as potential disrupters of host prothoracic gland function. Arch. Insect Biochem. Physiol. 2006, 61, 157–169. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvia, R.; Cozzolino, F.; Scieuzo, C.; Grimaldi, A.; Franco, A.; Vinson, S.B.; Monti, M.; Falabella, P. Identification and Functional Characterization of Toxoneuron nigriceps Ovarian Proteins Involved in the Early Suppression of Host Immune Response. Insects 2022, 13, 144. https://doi.org/10.3390/insects13020144
Salvia R, Cozzolino F, Scieuzo C, Grimaldi A, Franco A, Vinson SB, Monti M, Falabella P. Identification and Functional Characterization of Toxoneuron nigriceps Ovarian Proteins Involved in the Early Suppression of Host Immune Response. Insects. 2022; 13(2):144. https://doi.org/10.3390/insects13020144
Chicago/Turabian StyleSalvia, Rosanna, Flora Cozzolino, Carmen Scieuzo, Annalisa Grimaldi, Antonio Franco, S. Bradleigh Vinson, Maria Monti, and Patrizia Falabella. 2022. "Identification and Functional Characterization of Toxoneuron nigriceps Ovarian Proteins Involved in the Early Suppression of Host Immune Response" Insects 13, no. 2: 144. https://doi.org/10.3390/insects13020144
APA StyleSalvia, R., Cozzolino, F., Scieuzo, C., Grimaldi, A., Franco, A., Vinson, S. B., Monti, M., & Falabella, P. (2022). Identification and Functional Characterization of Toxoneuron nigriceps Ovarian Proteins Involved in the Early Suppression of Host Immune Response. Insects, 13(2), 144. https://doi.org/10.3390/insects13020144