Simple Summary
In this work, we aimed to resolve the identification of the peanut thrips, the key pest of Arachis hypogaea in South America. Based on morphological, biological, and molecular data, we conclude that the name historically applied to this pest, Enneothrips flavens, constitutes a misidentification and that the peanut thrips is actually an undescribed species, Enneothrips enigmaticus sp. n.
Abstract
The peanut thrips, Enneothrips enigmaticus sp. n., is the key pest of Arachis hypogaea L. in South America, where it can cause yield losses of up to 85%. This species has historically been identified as Enneothrips flavens, but access to the holotype of this species and freshly collected material from southeastern and northern Brazil revealed that specimens commonly collected on peanut crops are not conspecific with E. flavens. Biological, molecular, and morphological assessments were carried out and led to the conclusion that the key pest of A. hypogaea belongs to a previously undescribed species: Enneothrips enigmaticus sp. n.
1. Introduction
The peanut thrips is the key pest of peanuts crops, Arachis hypogaea L. (Fabaceae), in South America. This pest is found in Argentina, Brazil, and Paraguay and can cause losses of up to 85% [1,2]. The species is commonly found on leaflets, where it feeds on the tissue, causing bronze markings and deformation [1,3].
The first record of a name applied to the peanut thrips was Frankliniella fusca (Hinds), which was a misidentification subsequently corrected by Gallego de Sureda, who identified the species as Enneothrips (Enneothripiella) flavens Moulton [4]. This name was previously proposed by Moulton [5] to describe a species based on a unique female specimen collected in Minas Gerais state, Brazil, on “Indian tea” plants. Due to the identification by Gallego de Sureda [4], peanut thrips has been referred to as E. flavens and host-specific of A. hypogaea [6].
However, specimens of Enneothrips recently collected on plants other than A. hypogaea were identified as E. flavens. This fact suggests that either E. flavens is not host-specific or Enneothrips specimens previously collected from A. hypogaea were misidentified. Both hypotheses have a serious impact because if the species is not host-specific to A. hypogaea, the control tactics must take into account that alternative hosts may harbor the pest when peanut is not cultivated. On the other hand, if the name of the peanut thrips is not being properly used, the scientific nomenclature of all the literature on this species is incorrect. Thus, this taxonomic issue about the peanut thrips has not only systematic interest for a species, but also an economic impact.
Here, we report biological, molecular, and morphological evidence that reveals that the name currently applied to the peanut thrips is not appropriate. Therefore, we “unmask the villain” and reveal the real identity of this important pest species by proposing its reclassification as a new species to science—Enneothrips enigmaticus sp. n.
2. Materials and Methods
Fresh material of the peanut thrips was manually collected from this crop in Rio Branco (9°55′43.24″ S; 67°53′54.80″ W), Acre, and Jaboticabal (21°14′23.40″ S; 48°17′45.22″ W), São Paulo, Brazil. In addition, specimens to which the name Enneothrips flavens could be applied, were collected from fabaceous herbs and Adenanthera macrocarpa in Pedralva (22°15′55.00″ S; 45°24′37.00″ W), Minas Gerais, and from Campomanesia guazumifolia in Santo Antonio do Pinhal (22°48′38.55″ S; 45°42′14.24″ W), São Paulo, close to southern Minas Gerais, the type locality of the species. All of them were stored in ethanol 100% for slide mounting and DNA extraction. In addition, Enneothrips spp. specimens deposited in several collections were examined. Collection details are available in material examined. To correct the identification and description of the Enneothrips spp. collected on A. hypogaea and other hosts, we performed biological, morphological, and molecular assessments. In addition, we modelled the potential distribution of the new species based on 19 climactic factors.
2.1. Biological Assessment
Whenever possible, plant species with Enneothrips spp. were identified and, when available, larvae were collected to establish true host associations. Data available in [6] were used as evidence for host-specificity of the peanut thrips to A. hypogaea. As a consequence, second-instar larvae of both species discussed herein are described.
2.2. Morphological Assessment
As for morphological studies, comparison of the collected material with the holotype of E. flavens was carried out. In addition, the specimens were compared with type material of all other four valid Enneothrips species and with additional specimens whose morphology matches with E. flavens holotype in CHNUFPI, ESALQ, and USNM. Fresh material was prepared onto permanent microscope slides based on the technique proposed in [7] (adults) and in [8] (immatures). Photomicrographs of the specimens were taken under a Zeiss Axio Lab A1 microscope (Carl Zeiss Microscopy, Jena, Germany) with phase contrast and a Zeiss Axiocam ERc5 camera (Carl Zeiss Microscopy, Jena, Germany) attached.
In addition, SEM (Scanning Electron Microscopy) images were taken for specimens collected from peanuts on a Hitachi TableTop Scanning Microscope TM3000 (Hitachi High-Technologies Corporation, Tokyo, Japan) following the manufacturer’s guidelines.
Depositaries acronyms mentioned in this text are: CHNUFPI (Coleção de História Natural da Universidade Federal do Piauí, Floriano, Brazil), ESALQ (Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brazil), MLP (Museo de La Plata, La Plata, Argentina), CAS (California Academy of Sciences, San Francisco, CA, USA), and USNM (Smithsonian’s National Museum of Natural History, Washington, DC, USA).
2.3. Molecular Assessment
2.3.1. gDNA Extraction
Total genomic DNA was extracted from two individuals of E. flavens from Santo Antonio do Pinhal, SP, Brazil, and seven individuals of Enneothrips from peanuts from Jaboticabal (n = 4) and Rio Branco (n = 3), previously conserved in ethanol 100%. Non-destructive DNA extraction was performed with a modified protocol from [9]. Initially, insects were individualized in 1.5 mL Eppendorf tubes and were immersed in 0.3 mL of digestion buffer [3-mM CaCl2, 2% sodium dodecyl sulphate (SDS), 40 mM dithiothreitol (DTT), 100 mM Tris buffer pH 8, and 100 mM NaCl] with addition of 12.5 µL of proteinase K (20 µg ml−1) and incubated at 65 °C overnight (≈16 h). At the end of the incubation, the insects were removed and stored in 100% ethanol at −20 °C for morphological identification. After insect removal, the digestion buffer was mixed with 0.3 mL of chloroform + isoamyl alcohol (24:1) and centrifuged at 14,000 rpm for 20 min. The supernatant solution was transferred to a new Eppendorf with 30 µL of sodium acetate (3.0 M, pH 5.2), 2.5 µL of glycogen (5 mg ml−1), and 0.235 mL of iced 100% isopropanol. The mixture was gently vortexed and placed overnight at −20 °C and subsequently centrifuged at 14,000 rpm at 4 °C for 30 min. The liquid was removed, and the DNA pellet was serially washed with 0.4 mL of 70% ethanol and 95% ethanol. Lastly, the DNA pellet was air-dried, suspended in 25 µL of MilliQ-H2O, and stocked in a −20 °C freezer.
2.3.2. COI Amplification and Sequencing
The cytochrome c oxidase subunit I (COI) gene fragment was amplified by polymerase chain reaction (PCR) using the universal primers LCO1490 (5′ GGTCAACAAATCATAAAGATATTGG 3′) and HCO2198 (5′ TAAACTTCAGGGTGACCAAAAAATCA 3′) [10]. The PCR was performed in a total of 25 µL containing 3 µL genomic DNA (gDNA), 18.25 µL MilliQ-H2O, 0.25 µL 10X PCR Buffer Mg2+-free (Thermo Fisher Scientific ™), 1.25 µL MgCl2 (50 mM) (Thermo Fisher Scientific ™, Carlsbad, CA, USA), 0.125 µL dNTP (10 mM) (Sinapse Inc®) (Hollywood, FL, USA), 1 µL of each primer (5 µM), and 0.125 µL Platinum® (Carlsbad, CA, USA) Taq DNA Polymerase (5 U µL−1) (Thermo Fisher Scientific ™) (Carlsbad, CA, USA). The PCR amplification conditions used were 94 °C for 3 min for primary denaturation, then 35 cycles at 94 °C for 30 s, 53 °C for 45 s, 72 °C for 2 min, with a final extension at 72 °C for 10 min. PCR amplicons were visualized under ultraviolet light after electrophoresis using 3 μL amplicon in 2% (w v–1) agarose gel stained with SYBR Safe (Life Technologies). The subsequent purification process was performed using 1 µL (20 U µL−1) of Exonuclease I (Thermo Fisher Scientific ™) and 2 µL (1 U µL−1) of FastAP ™ Thermosensitive Alkaline Phosphatase (Thermo Fisher Scientific ™) for 10 µL of PCR final product. The conditions of thermocycler used for purification involved 30 min at 37 °C, followed by 15 min at 80 °C. The bidirectional sequencing was performed by the Sanger method using the same PCR primers in the Animal Biotechnology Laboratory at ESALQ, University of São Paulo.
2.4. Molecular Data Analysis
The chromatogram of each COI sequence was verified, aligned, and edited in a consensus sequence using Sequencher 4.8 (Gene Codes Corp., Ann Arbor, MI). Afterwards, COI sequences were aligned, and the presence of nuclear paralogs of mitochondrial origin (NumtS) [11] was observed following steps described in [12]: (i) insertions/deletions (indels), (ii) stop codons leading to premature protein termination, and (iii) increased rates of non-synonymous mutations. The presence of signatures (i) and (ii) would be enough to consider a sequence as a NumtS. The presence of signature (iii) would be used to confirm the NumtS status of the sequence. The genetic distance among haplotypes was estimated using the Kimura-two-parameters (K2P) model with 5000 bootstrap replications in MEGA X [13]. A Bayesian phylogenetic tree was estimated using the best substitution model of evolution GTR+I selected using the software MRMODELTEST v2.3 [14]. The Bayesian phylogenetic analyses were carried out in MRBAYES v3.1.2 [15], using two simultaneous runs of 25 million generations with one cold and three heated chains in each run. At the end of the runs, the first 25% of the trees were discarded as burn-in samples. The consensus tree of the two independent runs was obtained with posterior probabilities > 0.50. The COI sequence of Frankliniella occidentalis (NCBI accession number: HQ930545) was used as outgroup. No sequences of other Enneothrips spp. are available in NCBI; thus, sequences of the common species F. occidentalis were chosen exclusively to estimate the genetic distance, not phylogenetic inferences.
2.5. Species Distribution Modelling
As the distribution of E. enigmaticus sp. n. is not well studied, the potential distribution in the Americas was modelled based on taxonomic validated species records on peanuts from Thysanoptera collections from USNM, CHNUFPI MLP, and ESALQ (Table 1). For non-georeferenced records, we used the label information to retrieve the most probable coordinates. A total of 19 environmental variables were used to estimate the potential distribution, which included precipitation, temperature, moisture index, and radiation, at a 10 min spatial resolution. These were obtained from the WorldClim dataset and cover climate information in the period 1970–2000. Analysis for E. flavens was not performed because there are very few records of this species to make the analysis possible.

Table 1.
Approximate coordinates of reliable available records of Enneothrips enigmaticus sp. n. and Enneothrips flavens in South America.
Modelling was performed through multivariate analysis and maximum entropy using MaxEnt 3.4.4 [16]. MaxEnt estimates the probability of occurrence based on environmental parameters and has predictive power even with small datasets [17]. Model settings were defined after a bias analysis using the software R 4.0.2. The chosen model, with the least delta AIC, included linear features with a regularization multiplier of 1.0.
This information is relevant not only because of the economic importance of E. enigmaticus sp. n., but also because the model results provide indication of a possible shared distribution with E. flavens, which can help understand the evolutionary pathways of both species.
3. Results
Based on morphological, biological and molecular evidence detailed below, we describe the new species E. enigmaticus sp. n. as the pest on peanuts (Figure 1). We re-evaluate the morphological description of E. flavens while also describing the adult female and male and larvae II of both species. This information is essential for the recognition of both species and constitutes the first step towards the adoption of management tactics for the key peanut pest.

Figure 1.
Enneothrips enigmaticus sp. n., female, on peanut (Arachis hypogaea) (Photo: ESALQ Entomological Museum).
- Enneothrips enigmaticus sp. n.
Female macroptera. Color orange to reddish brown (Figure 2A). Antennal segments I and II light brown, III brown with base pale, IV and V brown with extreme base pale, VI–IX brown (Figure 2C). Head, thorax, and abdomen reddish brown, somewhat darker laterally (Figure 2D,F); fore wing reddish brown with pale basal fifth (Figure 2H); legs yellow; abdominal tergites uniformly reddish brown. Head with ocellar area without sculpture; ocellar setae III within the ocellar triangle (Figure 2D,E); pronotum and mesonotum transversely striate, around 20 lines of sculpture on mesonotum (Figure 2F,G); metanotum longitudinally sculptured on lateral thirds, transversely sculptured on anterior third, and reticulate on the median two-thirds, without internal lines inside reticles (Figure 2F,G). Abdominal tergites transversely striate laterally, about 15 lines of sculpture; tergite VIII with posteromarginal comb complete. Abdominal sternites II–VIII with microtrichia medially; posteromarginal setae anterior to margin.

Figure 2.
Enneothrips enigmaticus sp. n. (A) Female; (B) Male; (C) Antenna; (D) Head and pronotum; (E) Head and pronotum (SEM); (F) Meso- and metanotum; (G) Meso- and metanotum (SEM); (H) Fore wing (female); (I) Wings (male) (SEM); (J) Wings (male); (K) Abdominal sternites II–IV (male).
Measurements (holotype female in microns): Body length 1375. Head length 85; width 130; ocellar setae III length 15. Pronotum length 105; width 150; posteroangular setae length 35. Fore wing length 600. Antennal segments I–IX length 18, 32, 38, 42, 39, 39, 12, 10, 15.
Male brachyptera. Similar to female (Figure 2B), but smaller, brachypterous (Figure 2I,J), with a broad pore plate between the abdominal sternites II and III (Figure 2K).
Measurements (paratype male in microns): Body length 1000. Head length 65; width 110; ocellar setae III, length 12. Pronotum length 90; width 125; posteroangular setae length 30. Fore wing length 100. Pore plate length 30; width 78. Antennal segments I–IX length 12, 30, 30, 38, 32, 38, 10, 8, 10.
Larva II. Color, pale (Figure 3A) with brown areas anteromedially on head (Figure 3C), posteriorly on abdominal segment X (Figure 3F) and on the bases of femora and tibiae. Antennal segment I pale, II brown with pale apex, III brown with pale base and apex, IV–VII brown. Antennal segment IV with rows of microtrichia (Figure 3B). Body dorsal setae expanded, except for cephalic pair D3 acute (Figure 3C). Ventral setae acute. Body densely covered with sclerotized plates, absent on head, less numerous on pronotum and abdominal tergites IX and X (Figure 3F). Spiracles on mesonotum and abdominal tergites II and VIII; spiracle facets variably with zero or one pore (Figure 3D,E). Abdominal tergites IX and X with one pair of campaniform sensilla.

Figure 3.
Enneothrips spp. larvae II and hosts. Enneothrips enigmaticus sp. n.: (A) Habitus; (B) Antenna; (C) Head and pronotum; (D) Mesothoracic spiracle; (E) Abdominal tergite II spiracle; (F) Abdominal tergites VI-X. Enneothrips flavens: (G) Habitus; (H) Antenna; (I) Head and thorax; (J) Abdominal tergites. (K) Damage of E. enigmaticus sp. n. on peanut leaflets; (L) Campomanesia guazumifolia, host of E. flavens.
Measurements (in microns). Body length 1200. Head length 90; width 80; D1 setae length 13; D4 setae length 15. Pronotum length 85; width 150; D5 setae length 15; D6 setae length 22. Abdominal tergites IX length 60; width 75; setae D1 length 25; setae D2 length 30; X length 35; width 50. Antennal segments I–VII length 12, 27, 40, 42, 8, 10, 18.
Material studied. Holotype female. Brazil. São Paulo: Jaboticabal, UNESP-FCAV, Arachis hypogaea, 12.vii.2016 (J.R. Lima col.) (CHNUFPI).
Paratypes. BRAZIL. Same data as holotype, 4  4
4  4 larvae II (CHNUFPI). São Paulo: Campinas, same host, 18.i.1994, 4 (D. Gabriel col.) (ESALQ); Pindorama, same host, 15 2 xii.2016 (M. Michelotto col.) (CHNUFPI). Acre: Rio Branco, Embrapa Acre, Arachis pintoi, 17.vii.2014, 1 (R.S. Santos col.); same locality and host, 27.vii.2019 1 (E.F.B. Lima col.) (CHNUFPI).
4 larvae II (CHNUFPI). São Paulo: Campinas, same host, 18.i.1994, 4 (D. Gabriel col.) (ESALQ); Pindorama, same host, 15 2 xii.2016 (M. Michelotto col.) (CHNUFPI). Acre: Rio Branco, Embrapa Acre, Arachis pintoi, 17.vii.2014, 1 (R.S. Santos col.); same locality and host, 27.vii.2019 1 (E.F.B. Lima col.) (CHNUFPI).
4 4 larvae II (CHNUFPI). São Paulo: Campinas, same host, 18.i.1994, 4 (D. Gabriel col.) (ESALQ); Pindorama, same host, 15 2 xii.2016 (M. Michelotto col.) (CHNUFPI). Acre: Rio Branco, Embrapa Acre, Arachis pintoi, 17.vii.2014, 1 (R.S. Santos col.); same locality and host, 27.vii.2019 1 (E.F.B. Lima col.) (CHNUFPI).Non-type material. BRAZIL. São Paulo: Campinas, Arachis hypogaea, 14.xii.1993, 3 ; 21.xii.1993, 19 ; 4.i.1994, 49 2 (D. Gabriel col.); Piracicaba, same host, 10.iv.1991, 12 2 (R.C. Monteiro col.); Tupi Paulista, same host, no dates, 1 (Adalberto col.); Jaboticabal, same host, 1995, 2 larvae II (M.G.A. Lima col.) (ESALQ); Santa Adélia, same host, iii.2014, 2 ; Itápolis, same host, iii.2014, 1 (M. Michelotto col.) (CHNUFPI). Minas Gerais: Uberlândia, 30.vi.2020, 1 (G. Berchieri col.) (CHNUFPI) PARAGUAY. Boquerón, same host, 3.ii.2012, 2 (L.R.G Segnana col.) (CHNUFPI).
; 21.xii.1993, 19 ; 4.i.1994, 49 2 (D. Gabriel col.); Piracicaba, same host, 10.iv.1991, 12 2 (R.C. Monteiro col.); Tupi Paulista, same host, no dates, 1 (Adalberto col.); Jaboticabal, same host, 1995, 2 larvae II (M.G.A. Lima col.) (ESALQ); Santa Adélia, same host, iii.2014, 2 ; Itápolis, same host, iii.2014, 1 (M. Michelotto col.) (CHNUFPI). Minas Gerais: Uberlândia, 30.vi.2020, 1 (G. Berchieri col.) (CHNUFPI) PARAGUAY. Boquerón, same host, 3.ii.2012, 2 (L.R.G Segnana col.) (CHNUFPI).- Enneothrips flavens
Female macroptera. Similar to E. enigmaticus sp. n. (Figure 4A,C–E,G) but with darker brown shadings, especially medially on abdominal tergites (Figure 4A), mesonotum with around 35 lines of sculpture, and reticles on metanotum with internal markings (Figure 5E).

Figure 4.
Enneothrips flavens. (A) Female; (B) Male; (C) Antenna; (D) Head and pronotum; (E) Meso- and metanotum; (F) Abdominal sternites II–IV (male); (G) Fore wing.
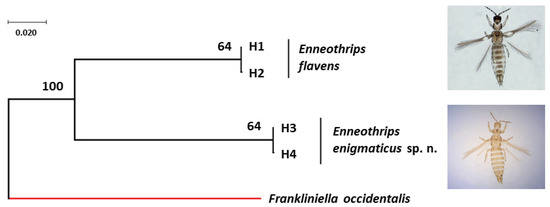
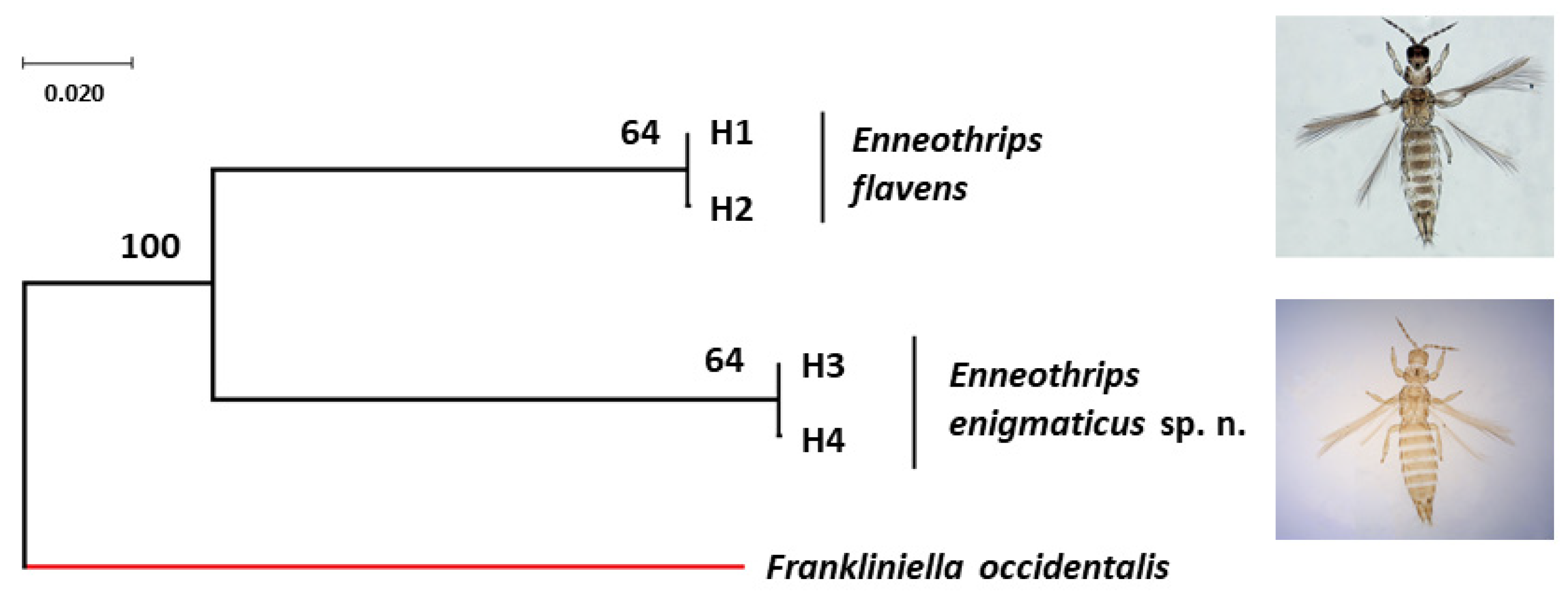
Figure 5.
Bayesian tree of Enneothrips flavens and Enneothrips enigmaticus sp. n. haplotypes with posteriori probability values for each node/branch based on cytochrome c oxidase subunit I (COI) gene fragment sequences. The COI sequence of Frankliniella occidentalis (NCBI accession number: HQ930545) was used as the outgroup.
Male macroptera. Similar to female (Figure 4B) but smaller and with a broad oval pore plate between the abdominal sternites II and III (Figure 4F).
Measurements (male in microns): Body length 1250. Head length 80; width 115; ocellar setae III, length 13. Pronotum length 105; width 130; posteroangular setae length 43. Fore wing length 570. Pore plate length 48; width 65. Antennal segments I–IX length 20, 30, 38, 48, 40, 38, 12, 10, 15.
Larva II. Similar to Enneothrips enigmaticus sp. n. (Figure 3G–J) Dorsal setae usually longer than in E. enigmaticus sp. n. (Figure 3H–J), but the measurements of specimens of both species overlap.
Measurements (in microns). Body length 1100. Head length 85; width 80; D1 setae length 18; D4 setae length 22. Pronotum length 85; width 150; D5 setae length 18; D6 setae length 28. Abdominal tergites IX length 40; width 50; setae D1 length 28; setae D2 length 30; X length 35; width 28. Antennal segments I–VII length 12, 25, 38, 42, 10, 8, 15.
Material studied. BRAZIL. Holotype female. Minas Gerais, India tea foliage, 23.v.1933 (E.J. Hambleton) (CAS). Santa Catarina: Nova Teutônia, Allophylus, 2.xi.1949, 2 2 (F. Plaumann col.) (USNM); Descanso (Linha Famoso), angico (Adenanthera macrocarpa) 18.vii.2019, 1 1 (E.F.B. Lima col.) (CHNUFPI). São Paulo: Piracicaba, shrubs, 13.iii.1997, 1 4 (L.A. Mound col.) (ESALQ); Santo Antonio do Pinhal, Camponesia guazumifolia, 21.x.2018, 5 2 larvae II (E.F.B. Lima col.) (CHNUFPI). Minas Gerais: Pedralva, herbaceous Fabaceae, 15.iv.2017, 3 3 , same data on Adenanthera macrocarpa, 1 ; Tiradentes, Serra de São José, 16.xii.2019, 2 (E.F.B. Lima col.) (CHNUFPI).
2 (F. Plaumann col.) (USNM); Descanso (Linha Famoso), angico (Adenanthera macrocarpa) 18.vii.2019, 1 1 (E.F.B. Lima col.) (CHNUFPI). São Paulo: Piracicaba, shrubs, 13.iii.1997, 1 4 (L.A. Mound col.) (ESALQ); Santo Antonio do Pinhal, Camponesia guazumifolia, 21.x.2018, 5 2 larvae II (E.F.B. Lima col.) (CHNUFPI). Minas Gerais: Pedralva, herbaceous Fabaceae, 15.iv.2017, 3 3 , same data on Adenanthera macrocarpa, 1 ; Tiradentes, Serra de São José, 16.xii.2019, 2 (E.F.B. Lima col.) (CHNUFPI).Morphological comparison of specimens collected in peanuts (E. enigmaticus sp. n.) with the types of species currently classified in Enneothrips confirmed that E. flavens is the closest related species. However, the following morphological differences between E. enigmaticus sp. n. and E. flavens were noticed: (i) internal markings between sculpturing lines on metanotum present only in E. flavens (Figure 2F and Figure 4E); (ii) sculpturing lines on mesonotum less numerous in E. enigmaticus sp. n. (Figure 2F,G); (iii) abdominal tergites lighter and more uniformly colored in E. enigmaticus sp. n. (Figure 2A,B) and shaded brown medially in E. flavens (Figure 4A,B). In addition, individuals morphologically identical to the E. flavens holotype were collected in the states of São Paulo and Minas Gerais and were examined from mounted material deposited in USNM from Santa Catarina state. Among the individuals, males of E. enigmaticus sp. n. were brachypterous (Figure 2B), while males of E. flavens were always macropterous (Figure 4B).
Forward and reverse sequences were analyzed together, but as they generated different sizes, a consensus sequence was made, according to the usual procedure for estimating haplotypes. COI sequences were edited in a consensus size of 672 pb. Evidence of NumtS presence was not found in the sequences. The two sequences of E. flavens resulted in two haplotypes (H1 and H2) separated by a single mutation step (Figure 5) (NCBI accession number: H1 = MT947751 and H2 = MT947752). The seven sequences of E. enigmaticus sp. n. resulted also in the presence of two haplotypes (H3 and H4) separated by a single mutation step and distributed in both sample sites, Jaboticabal (H3 = 2 and H4 = 1) and Rio Branco (H3 = 1 and H4 = 3) (NCBI accession number: H3 = MT947753-MT947755 and H4 = MT947756-MT947759) (Figure 5). The intraspecific genetic distance of the COI gene fragment under each species was 0.0015. The interspecific genetic distance (K2P) of the COI gene fragment between E. flavens and E. enigmaticus sp. n. varied from 0.1872 to 0.1892.
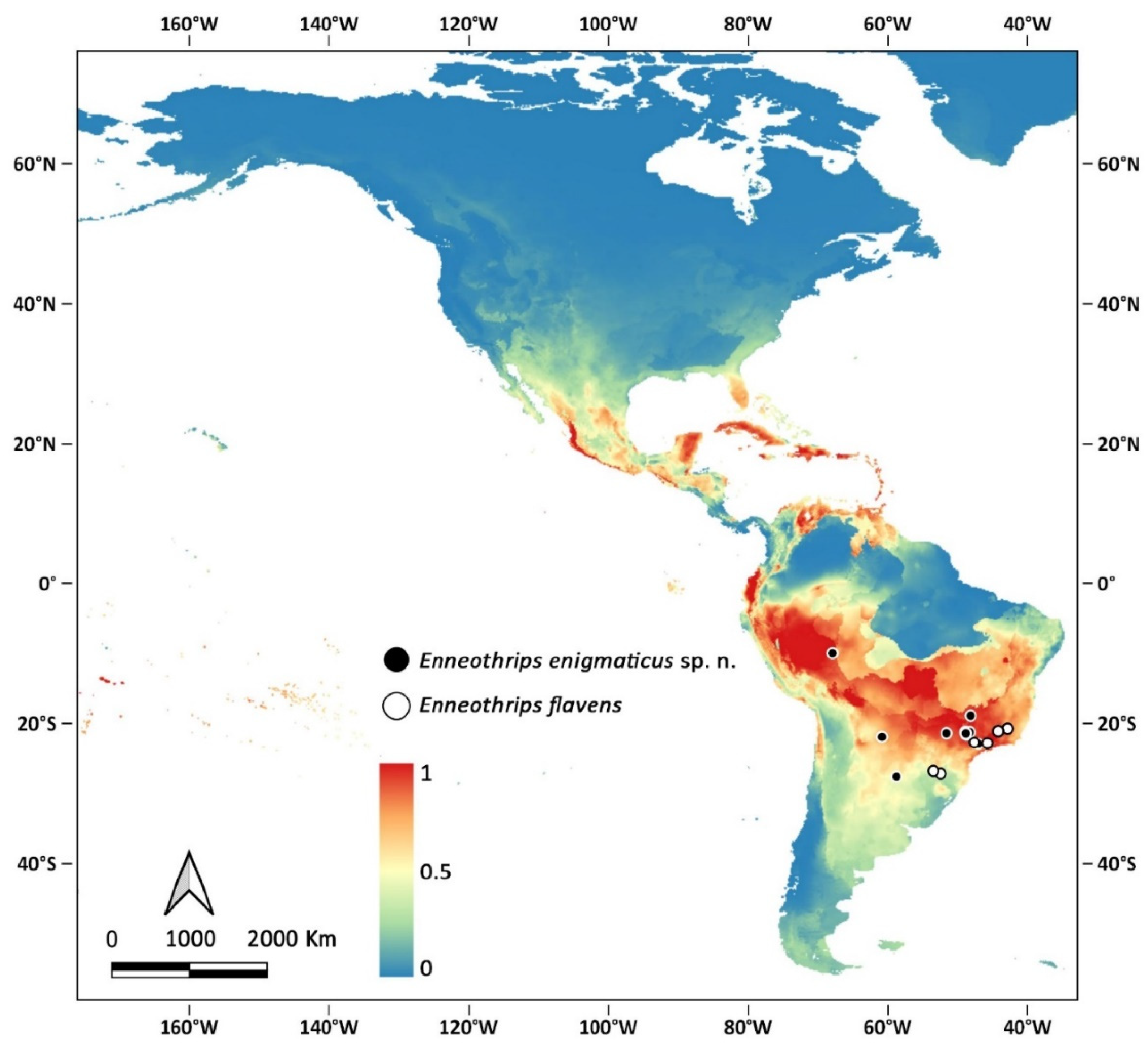
Enneothrips enigmaticus sp. n. and E. flavens are the only representatives of the genus occurring south of 20° S, and at present the only confirmed records are in South America, especially near the Tropic of Capricorn. Modelling, however, indicates that the species may find suitable conditions in other areas of the continent, especially in tropical countries (Figure 6).
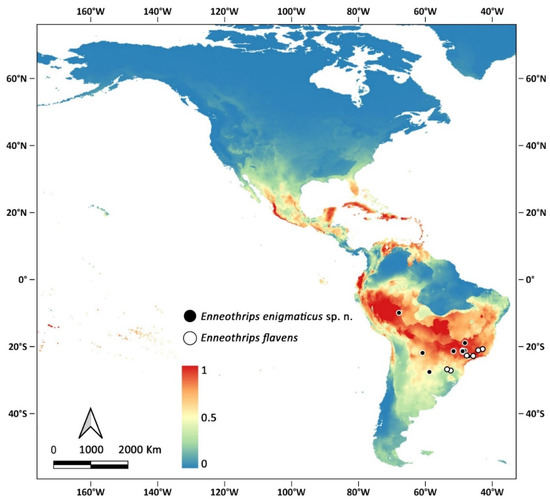
Figure 6.
Confirmed records of Enneothrips spp. (circular markings) and modelling distribution of Enneothrips enigmaticus sp. n. in the Americas.
4. Discussion
The differences reported above are based on comparisons of more than 100 specimens of Enneothrips from peanuts either collected or examined from collections and almost 30 individuals identified here as E. flavens, including the species holotype. Thus, they constitute robust evidence that the name historically applied to the key peanut pest has been misidentified. Males of both Enneothrips enigmaticus sp. n. and E. flavens exhibit an internal pore plate between sternites II and III, a character shared with other thrips genera from the Neotropics, namely, Ameranathrips, Apterothrips, Baileyothrips, Charassothrips, Desartathrips, Enneothrips, Pseudothrips, Psydrothrips, and Xerothrips [18]. The easily found males of E. enigmaticus sp. n. might open possibilities to study their behavior and the significance of such pore plate character. It is also possible to develop studies aiming to understand the role of the reduced wings in E. enigmaticus sp. n. males, a character found only in this species in the genus.
Immatures of peanut thrips have been found only from that crop, although a few adults have been collected from other plants [6]. In our surveys, we found larvae of E. enigmaticus sp. n. on peanuts, thus confirming the host association with the A. hypogaea (Figure 3K). A good number of adults have also been collected from Arachis pintoi¸ a possible alternative host in the same plant genus that could indicate Arachis spp. as hosts of E. enigmaticus sp. n. As for E. flavens, no specimens that match its holotype morphology have been collected from peanuts. On the other hand, several specimens of this thrips species were collected from at least four plant species, one of which (Campomanesia guazumifolia) (Figure 3L) with immatures. The large number of adult specimens from several plants and the larvae found in one of the plant species suggests that, unlike E. enigmaticus sp. n., E. flavens is probably oligophagous. That could explain the accidental collection of the species holotype from “Indian tea” plants, unrelated to A. hypogaea.
The genetic distance between E. flavens and E. enigmaticus sp. n. individuals is higher than that between other thrips species and supports the presence of two Enneothrips species [19,20,21].
Based on the resemblance and close distribution, it is feasible to indicate that E. enigmaticus sp. n. and E. flavens are two sister species. A possibility to explain the origin of the two species is an ancestor in South America whose populations were separated by a functional character. While E. enigmaticus sp. n. specialized breeding on Arachis, E. flavens became an oligophagous species. Arachis hypogaea, the main host of E. enigmaticus sp. n. is native to South America, most probably to the area around Southern Bolivia, Northern Argentina, and Western Paraguay [22,23], but also reaching Peru, Chile, and Western Brazil. As host-specific, that might be the area of origin of the thrips species as well. Specimens of E. flavens have been collected from Western Santa Catarina state, close to the border of Brazil with Paraguay and Argentina, reinforcing the idea of a shared natural occurrence (Figure 3), also evidenced by the modelling distribution of the new species and the confirmed records of E. flavens. Another possibility is that a lineage of Enneothrips invaded Central and Southern South America and was subsequently divided by vicariance. The western population would have specialized in Arachis, giving raise to E. enigmaticus sp. n., which was subsequently transported further into Southeastern Brazil with its host on cultivated peanuts. These or other explanations can be explored in a phylogeographic analysis.
5. Conclusions
Based on morphological, biological, and molecular evidence, we conclude that the peanut thrips, Enneothrips enigmaticus sp. n., was previously misidentified as Enneothrips flavens and is here described as a new species.
Author Contributions
Conceptualization, É.F.B.L. and A.S.C.; Methodology, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Software, Á.R.S.d.A. and F.N.; Validation, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Formal Analysis, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Investigation, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Resources, É.F.B.L., M.D.M. and A.S.C.; Data Curation, É.F.B.L. and A.S.C.; Writing—Original Draft Preparation, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Writing—Review & Editing, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Visualization, É.F.B.L., Á.R.S.d.A., F.N., M.D.M. and A.S.C.; Supervision, É.F.B.L.; Project Administration, É.F.B.L. and M.D.M.; Funding Acquisition, É.F.B.L. and M.D.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), through process number 2020/12581-1, and by Universidade Federal do Piauí (UFPI), through the concession of a scientific initiation scholarship to the second author (PI 2855-2017).
Institutional Review Board Statement
Not applicable.
Acknowledgments
To Robert Zuparko, Rachel Díaz-Bastin and Chris Grinter for allowing access to the Thysanoptera collection of the California Academy of Sciences, San Francisco, CA, USA; to Cheryle O’Donnell, for allowing access to the USNM’s Thysanoptera Collection in Beltsville, MD, USA and for kindly providing the facilities used in this work to produce SEM images of the new species; to Roberto A. Zucchi, for allowing access to the Thysanoptera collection of Escola Superior de Agricultura “Luiz de Queiroz”, University of São Paulo, Piracicaba, Brazil; to María Inés Zamar (UNJu, Argentina) and Everth Ebratt (INCA, Colombia), for confirming information on Enneothrips from Argentina and Colombia, respectively; to Rodrigo Souza (Embrapa Acre), José Ricardo Lima (UNESP) and Osmar Arias (UNA, Paraguay) for providing specimens used in this research and/or helping during field work; to Elisa Miyasato (IFSudesteMG, Brazil) for assistance in field works; to Laurence Mound (CSIRO) for his valuable comments on an early version of this work. Collections carried out by É.F.B.L. were authorized by the Sistema de Autorização e Informação em Biodiversidade (SISBIO), Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio), Ministry of Environment, through License # 54104.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Nakano, O. Entomologia Econômica; Octávio Nakano: Piracicaba, Brazil, 2011. [Google Scholar]
- Lima, E.F.B. Thysanoptera. Catálogo Taxonômico da Fauna do Brasil. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/316 (accessed on 16 December 2021).
- Quintanilla, R.H. Trips, Características Morfológicas y Biológicas: Especies de Mayor Importancia Agrícola; Hemisferio Sur: Buenos Aires, Argentina, 1980. [Google Scholar]
- Gallego de Sureda, A.E. El trips del mani: Enneothrips (Enneothripiella) flavens. Rev. Fac. Agron. Univ. Nac. Plata 1969, 44, 167–172. [Google Scholar]
- Moulton, D. Thysanoptera from Minas Geraes, Brazil (second paper). Rev. Entomol. 1941, 12, 314–322. [Google Scholar]
- Lima, M.G.A.; Martinelli, N.M.; Monteiro, R.C. Plantas hospedeiras de tripes no período da entressafra do amendoim. Rev. Agr. 2000, 75, 129–135. [Google Scholar]
- Mound, L.A.; Marullo, R. The thrips of Central and South America: An introduction (Insecta: Thysanoptera). Mem. Entomol. Int. 1996, 6, 1–487. [Google Scholar]
- Lima, E.F.B.; O’Donnell, C.A.; Sousa, Y.L.C.; Zamar, M.I. Identification of second instar larvae of Panchaetothripinae (Thysanoptera, Thripidae) in Brazil and Argentina. Zootaxa 2021, 5047, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.T.P.; Moore, W.; Melchior, L.; Worobey, M. DNA Extraction from Dry Museum Beetles without Conferring External Morphological Damage. PLoS ONE 2007, 2, e272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Lopez, J.V.; Yuhki, N.; Masuda, R.; Modi, W.; O’Brien, S.J. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Mol. Evol. 1994, 39, 174–190. [Google Scholar] [PubMed]
- Correa, A.S.; Vinson, C.C.; Braga, L.S.; Guedes, R.N.C.; Oliveira, L.O. Ancient origin and recent range expansion of the maize weevil Sitophilus zeamais, and its genealogical relationship to the rice weevil S. Oryzae. Bull. Entomol. Res. 2017, 107, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrAIC.pl. Program Distributed by the Author. Evolutionary Biology Centre, Uppsala University. Available online: https://github.com/nylander/MrAIC (accessed on 22 September 2021).
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 15 September 2021).
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- De Borbón, C.M. Desertathrips chuquiraga gen. et sp.n. (Thysanoptera, Thripidae) from Argentina. Zootaxa 2008, 1751, 25–34. [Google Scholar] [CrossRef]
- Glover, R.H.; Collins, D.W.; Walsh, K.; Boonham, N. Assessment of loci for DNA barcoding in the genus Thrips (Thysanoptera: Thripidae). Mol. Ecol. Resour. 2010, 10, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, R.; Ashfaq, M.; Rasool, A.; Hebert, P.D. DNA barcode analysis of thrips (Thysanoptera) diversity in Pakistan reveals cryptic species complexes. PLoS ONE 2016, 11, e0146014. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Singha, D.; Kumar, V.; Pakrashi, A.; Kundu, S.; Chandra, K.; Patnaik, S.; Tyagi, K. DNA barcoding of selected Scirtothrips species (Thysanoptera) from India. Mitochondrial DNA B Resour. 2019, 4, 2710–2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriele, M.; Chalup, L.; Robledo, G.; Seijo, G. Genetic and geographic origin of domesticated peanut as evidenced by 5S rDNA and chloroplast DNA sequences. Plant Syst. Evol. 2012, 298, 1151–1165. [Google Scholar]
- Anurudh, K.S.; Nigam, S.N. Arachis gene pools and genetic improvement in groundnut. In Gene Pool Diversity and Crop Improvement; Rajpal, V.R., Rao, S.M., Raina, S.N., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 17–77. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).