
Insect Feeding on Sorghum bicolor Pollen and Hymenoptera Attraction to Aphid-Produced Honeydew

 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Study 1: Sweet Sorghum and Pollen Feeding
2.1.1. Plant Material
2.1.2. Insect Identification and Plant Morphological Measurements
2.1.3. Analysis
2.2. Study 2: Aphid Honeydew and Hymenoptera Diversity
2.2.1. Sorghum
2.2.2. Johnsongrass
2.2.3. Analyses
3. Results
3.1. Study 1
3.1.1. Insects Observed Collecting or Consuming Pollen
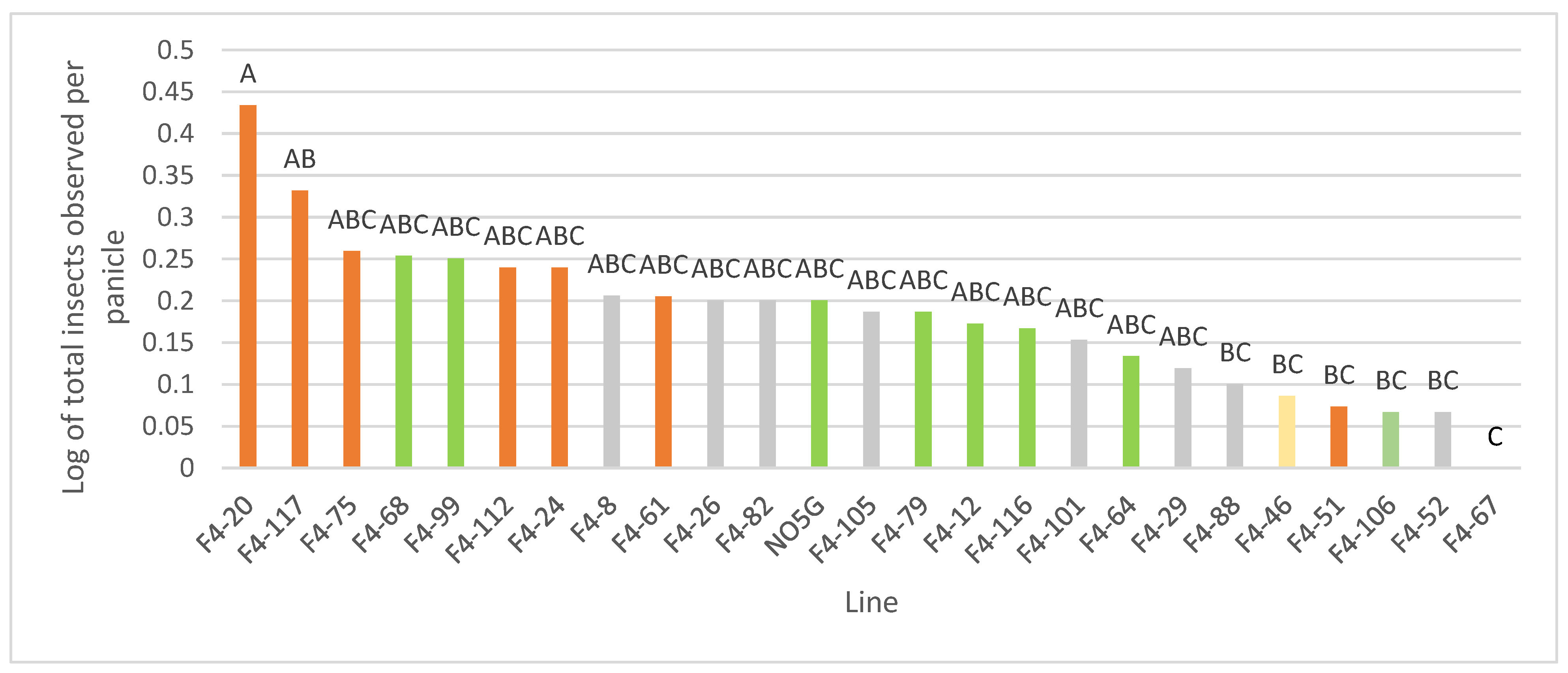
3.1.2. Distribution of Insects and Morphological Traits
3.1.3. Probability of Insect Observations by Date
3.1.4. Differences in F4 Morphological Traits
3.2. Study 2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- McGregor, S.E. Insect Pollination of Cultivated Crop Plants; Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1976. [Google Scholar]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Altierr, M.; Schmidt, L. Cover crops affect insect and spider populations in apple orchards. Calif. Agric. 1986, 40, 15–17. [Google Scholar]
- Andow, D.A. Vegetational diversity arthropod population. Annu. Rev. Entomol. 1991, 36, 561–566. [Google Scholar] [CrossRef]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Saunders, M.E. Insect pollinators collect pollen from wind-pollinated plants: Implications for pollination ecology and sustainable agriculture. Insect Conserv. Divers. 2018, 11, 13–31. [Google Scholar] [CrossRef]
- Decourtye, A.; Mader, E.; Desneux, N. Landscape enhancement of floral resources for honey bees in agro-ecosystems. Apidologie 2010, 41, 264–277. [Google Scholar] [CrossRef]
- Joseph, V.S.; Harris-Shultz, K.; Jespersen, D. Evidence of pollinators foraging on centipedegrass inflorescences. Insects 2020, 11, 795. [Google Scholar] [CrossRef]
- Immelman, K.; Eardley, C. Gathering of grass pollen by solitary bees (Halictidae, Lipotriches) in South Africa. Zoosystematics Evol. 2000, 76, 263–268. [Google Scholar] [CrossRef]
- Siede, R.; Eickhoff, B.; Freyer, C.; Windpassinger, S.; Büchler, R. The bioenergy crop Sorghum bicolor is a relevant pollen source for honey bees (Apis mellifera). GCB Bioenergy 2021, 13, 1149–1161. [Google Scholar] [CrossRef]
- Schmidt, M.; Bothma, G. Risk Assessment for transgenic sorghum in Africa: Crop-to-crop gene flow in Sorghum bicolor (L.) Moench. Crop Sci. 2006, 46, 790–798. [Google Scholar] [CrossRef]
- Checkoff, S. USDA Report Boosts Sorghum Acreage to 7.3 million, Most Since 2015. Available online: https://www.sorghumcheckoff.com/press-releases/usda-report-boost-sorghum-acreage-to-7-3-million-most-since-2015/ (accessed on 12 September 2022).
- Bowling, R.D.; Brewer, M.J.; Kerns, D.L.; Gordy, J.; Seiter, N.; Elliott, N.E.; Buntin, G.D.; Way, M.O.; Royer, T.A.; Biles, S. Sugarcane aphid (Hemiptera: Aphididae): A new pest on sorghum in North America. J. Integr. Pest Manag. 2016, 7, 1–13. [Google Scholar] [CrossRef]
- Fischer, M.K.; Völkl, W.; Hoffmann, K.H. Honeydew production and honeydew sugar composition of polyphagous black bean aphid, Aphis fabae (Hemiptera: Aphididae) on various host plants and implications for ant-attendance. Eur. J. Entomol. 2005, 102, 155–160. [Google Scholar] [CrossRef]
- Sabri, A.; Vandermoten, S.; Leroy, P.D.; Haubruge, E.; Hance, T.; Thonart, P.; De Pauw, E.; Francis, F. Proteomic investigation of aphid honeydew reveals an unexpected diversity of proteins. PLoS ONE 2013, 8, e74656. [Google Scholar] [CrossRef] [PubMed]
- Wäckers, F.L.; Van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a food source for natural enemies: Making the best of a bad meal? Biol. Control 2008, 45, 176–184. [Google Scholar] [CrossRef]
- Armstrong, J.S.; Rooney, W.L.; Peterson, G.C.; Villenueva, R.T.; Brewer, M.J.; Sekula-Ortiz, D. Sugarcane aphid (Hemiptera: Aphididae): Host range and sorghum resistance including cross-resistance from greenbug sources. J. Econ. Entomol. 2015, 108, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, R.T. Copious Amounts of Honeydew Produced by Sugarcane Aphids Attracts Bees and Other Insects: Do We Need to Be Worried. Available online: https://kentuckypestnews.wordpress.com/2017/09/12/copious-amounts-of-honeydew-produced-by-sugarcane-aphids-attracts-bees-and-other-insects-do-we-need-to-be-worried/ (accessed on 12 September 2022).
- Gorz, H.J.; Haskins, F.A.; Johnson, B.E. Registration of 15 germplasm lines of grain sorghum and sweet sorghum. Agron. Publ. 1990, 30, 231. [Google Scholar] [CrossRef]
- Harris-Shultz, K.; Punnuri, S.; Knoll, J.E.; Ni, X.; Wang, H. The sorghum epicuticular wax locus bloomless2 reduces plant damage in p898012 caused by the sugarcane aphid. Agrosystems Geosci. Environ. 2020, 3, e20008. [Google Scholar] [CrossRef]
- Uchimiya, M.; Knoll, J.E. Accumulation of carboxylate and aromatic fluorophores by a pest-resistant sweet sorghum [Sorghum Bicolor (L.) Moench] Genotype. ACS Omega 2019, 4, 20519–20529. [Google Scholar] [CrossRef]
- Haar, P.J.; Bowling, R.; Gardner, W.A.; Buntin, G.D. Epizootics of the entomopathogenic fungus Lecanicillium lecanii (Hypocreales: Clavicipitaceae) in sugarcane aphid (Hemiptera: Aphididae) populations infesting grain sorghum in Georgia and Texas1. J. Entomol. Sci. 2018, 53, 104–106. [Google Scholar] [CrossRef]
- Yeom, D.-J.; Kim, J.H. Comparative evaluation of species diversity indices in the natural deciduous forest of Mt. Jeombong. Forest Sci. Technol. 2011, 7, 68–74. [Google Scholar] [CrossRef]
- Standifer, L.N. A comparison of the protein quality of pollens for growth-stimulation of the hypopharyngeal glands and longevity of honey bees, Apis mellifera L.(Hymenoptera: Apidae). Insectes Soc. 1967, 14, 415–425. [Google Scholar] [CrossRef]
- Shen, L. Nutritional and chemical properties of sorghum, rapeseed, and sunflower pollens (Brassica napus, Helianthus annuus, Apis mellifera). Master’s Thesis, University of Arizona, Tucson, AZ, USA, 1992. [Google Scholar]
- Nunes-Silva, P.; Cordeiro, G.D.; Obregon, D.; Neto, J.F.L.; Thompson, F.C.; Viana, B.F.; Freitas, B.M.; Kevan, P.G. Pollenivory in larval and adult flower flies: Pollen availability and visitation rate by Toxomerus politus Say (Diptera: Syrphidae) on sorghum Sorghum bicolor (L.) MOENCH (Poaceae). Stud Dipterol. 2010, 17, 177–185. [Google Scholar]
- Richardson, C.H. A contribution to the life history of the cornfeeding syrphus fly (Mesogramma polita Say). J. Econ. Entomol. 1915, 8, 338–342. [Google Scholar] [CrossRef]
- Briceño, R.D.; Schüch, W. Reproductive biology and behavior of Doru taeniatum (Forficulidae). Rev. Biol. Trop. 1988, 36, 437–440. [Google Scholar]
- Chisausky, J.L.; Soley, N.M.; Kassim, L.; Bryan, C.J.; Miranda, G.F.G.; Gage, K.L.; Sipes, S.D. Syrphidae of southern Illinois: Diversity, floral associations, and preliminary assessment of their efficacy as pollinators. Biodivers. Data J. 2020, 8, e57331. [Google Scholar] [CrossRef]
- Van Huis, A. Integrated Pest Management in the Small Farmer’s Maize Crop in Nicaragua; Wageningen University & Research: Wageningen, The Netherlands, 1981. [Google Scholar]
- Vicens, N.; Bosch, J. Weather-dependent pollinator activity in an apple orchard, with special reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ. Entomol. 2000, 29, 413–420. [Google Scholar] [CrossRef]
- Brooking, I.R. Male sterility in Sorghum bicolor (L.) Moench induced by low night temperature. II. Genotypic differences in susceptibility. Aust. J. Plant Physiol. 1979, 6, 143–147. [Google Scholar]
- Klecka, J.; Hadrava, J.; Biella, P.; Akter, A. Flower visitation by hoverflies (Diptera: Syrphidae) in a temperate plant-pollinator network. PeerJ 2018, 6, e6025. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C. Lethal and sublethal synergistic effects of a new systemic pesticide, Flupyradifurone (Sivanto®), on honeybees. Proc. R. Soc. B 2019, 286, 20190433. [Google Scholar] [CrossRef]
- Chakrabarti, P.; Carlson, E.A.; Lucas, H.M.; Melathopoulos, A.P.; Sagili, R.R. Field rates of SivantoTM (Flupyradifurone) and Transform® (Sulfoxaflor) increase oxidative stress and induce apoptosis in honey bees (Apis mellifera L.). PLoS ONE 2020, 15, e0233033. [Google Scholar] [CrossRef] [PubMed]
- Mundy-Heisz, K.A.; Prosser, R.S.; Raine, N.E. Acute oral toxicity and risks of exposure to the neonicotinoid thiamethoxam, and other classes of systemic insecticide, for the common eastern bumblebee (Bombus impatiens). bioRxiv 2020. [Google Scholar] [CrossRef]
- Linguadoca, A.; Rizzi, C.; Villa, S.; Brown, M.J.F. Sulfoxaflor and nutritional deficiency synergistically reduce survival and fecundity in bumblebees. Sci. Total Environ. 2021, 795, 148680. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, G.; Pereira-Peixoto, M.-H.; Borth, J.; Lotz, S.; Wintermantel, D.; Allan, M.J.; Dean, R.; Schwarz, J.M.; Knauer, A.; Albrecht, M. Fungicide and insecticide exposure adversely impacts bumblebees and pollination services under semi-field conditions. Environ. Int. 2021, 157, 106813. [Google Scholar] [CrossRef] [PubMed]
- Bayer Sivanto Prime Insecticide: Sorghum. Available online: https://www.cropscience.bayer.us/-/media/Bayer-CropScience/Country-United-States-Internet/Documents/Products/Insecticides/Sivanto/Sivanto-Prime-Sorghum-Technical-Bulletin.ashx (accessed on 12 September 2022).
- Corteva Transform WG-Superior Insect Control in Cotton and Sorghum. Available online: https://www.corteva.us/content/dam/dpagco/corteva/na/us/en/products/files/TransformWG_CottonandSorghum_FS.pdf (accessed on 12 September 2022).
- Anderson, A.; McCormack, S.; Helden, A.; Sheridan, H.; Kinsella, A.; Purvis, G. The potential of parasitoid hymenoptera as bioindicators of arthropod diversity in agricultural grasslands. J. Appl. Ecol. 2011, 48, 382–390. [Google Scholar] [CrossRef]
- Roulston, T.H.; Smith, S.A.; Brewster, A.L. A Comparison of pan trap and intensive net sampling techniques for documenting a bee (Hymenoptera: Apiformes) fauna. J. Kansas Entomol. Soc. 2007, 80, 179–181. [Google Scholar] [CrossRef]
- Pak, D.; Iverson, A.L.; Ennis, K.K.; Gonthier, D.J.; Vandermeer, J.H. Parasitoid wasps benefit from shade tree size and landscape complexity in mexican coffee agroecosystems. Agric. Ecosyst. Environ. 2015, 206, 21–32. [Google Scholar] [CrossRef]
- Smith, R.G.; Gross, K.L.; Robertson, G.P. Effects of crop diversity on agroecosystem function: Crop yield response. Ecosystems 2008, 11, 355–366. [Google Scholar] [CrossRef]
- Barel, J.M.; Kuyper, T.W.; de Boer, W.; Douma, J.C.; De Deyn, G.B. Legacy effects of diversity in space and time driven by winter cover crop biomass and nitrogen concentration. J. Appl. Ecol. 2018, 55, 299–310. [Google Scholar] [CrossRef]
- Calderone, N.W. Insect pollinated crops, insect pollinators and us agriculture: Trend analysis of aggregate data for the period 1992–2009. PLoS ONE 2012, 7, e37235. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Family | Organism | Common Name | Total No. Observed |
|---|---|---|---|
| Syrphidae | Toxomerus politus | Maize calligrapher | 83 |
| Apidae | Apis mellifera | Honeybee | 43 |
| Forficulidae | Doru taeniatum | Earwig | 23 |
| Apidae | Bombus sp. | Bumblebee | 2 |
| Apidae y | Xylocopa micans | Carpenter bee | 1 |
| Sorghum | |||||
|---|---|---|---|---|---|
| With Aphids | Without Aphids | ||||
| Superfamily | Family | # of Morphospecies | # of Individuals | # of Morphospecies | # of Individuals |
| Apoidea | |||||
| ** Halictidae | 12 | 618 | 6 | 48 | |
| Apidae | 9 | 10 | 1 | 1 | |
| Andrenidae | 1 | 5 | 1 | 1 | |
| * Sphecidae | 19 | 41 | 4 | 6 | |
| Megachilidae | 0 | 0 | 0 | 0 | |
| Pompiloidea | |||||
| Pompilidae | 8 | 34 | 5 | 37 | |
| Mutillidae | 1 | 4 | 0 | 0 | |
| Chrysidoidea | |||||
| Chrysididae | 1 | 1 | 0 | 0 | |
| Bethylidae | 1 | 5 | 2 | 8 | |
| Dryinidae | 1 | 1 | 0 | 0 | |
| Tiphioidea | |||||
| Tiphiidae | 1 | 7 | 1 | 3 | |
| Sierolomorphidae | 1 | 1 | 0 | 0 | |
| Ichneumonoidea | |||||
| Ichnuemonidae | 2 | 2 | 1 | 1 | |
| * Brachonidae | 7 | 17 | 2 | 3 | |
| Chalcidoidea | |||||
| Aphelinidae | 2 | 5 | 1 | 1 | |
| Chalcidae | 3 | 7 | 0 | 0 | |
| ** Encyrtidae | 6 | 89 | 5 | 26 | |
| Eupelmidae | 1 | 1 | 1 | 1 | |
| Eurytomidae | 2 | 3 | 0 | 0 | |
| ** Mymaridae | 4 | 48 | 4 | 9 | |
| Pteromalidae | 7 | 26 | 1 | 50 | |
| Torymidae | 1 | 6 | 1 | 3 | |
| Eulophidae | 2 | 3 | 0 | 0 | |
| Perilampidae | 1 | 1 | 1 | 1 | |
| Cynipoidea | |||||
| Figitidae | 4 | 17 | 1 | 5 | |
| Diaprioidea | |||||
| * Diapriidae | 2 | 15 | 1 | 1 | |
| Ceraphronoidea | |||||
| Ceraphronidae | 1 | 2 | 1 | 1 | |
| Platygastroidea | |||||
| Platygastridae | 3 | 13 | 2 | 11 | |
| ** Scelionidae | 12 | 376 | 8 | 179 | |
| Formicoidea | |||||
| ** Formicidae | 3 | 528 | 3 | 85 | |
| Evanioidea | |||||
| Evaniidae | 0 | 0 | 0 | 0 | |
| Unknown | 3 | 6 | 3 | 3 | |
| TOTALS | 121 | 1892 | 56 | 484 | |
| Johnsongrass | |||||
|---|---|---|---|---|---|
| With Aphids | Without Aphids | ||||
| Superfamily | Family | # of Morphospecies | # of Individuals | # of Morphospecies | # of Individuals |
| Apoidea | |||||
| ** Halictidae | 13 | 78 | 3 | 3 | |
| Apidae | 2 | 2 | 0 | 0 | |
| Andrenidae | 3 | 3 | 0 | 0 | |
| * Sphecidae | 6 | 10 | 0 | 0 | |
| Megachilidae | 1 | 1 | 0 | 0 | |
| Pompiloidea | |||||
| * Pompilidae | 9 | 16 | 2 | 1 | |
| Mutillidae | 2 | 16 | 0 | 0 | |
| Chrysidoidea | |||||
| Chrysididae | 1 | 1 | 0 | 0 | |
| Bethylidae | 4 | 9 | 1 | 7 | |
| Dryinidae | 0 | 0 | 0 | 0 | |
| Tiphioidea | |||||
| Tiphiidae | 1 | 4 | 0 | 0 | |
| Sierolomorphidae | 0 | 0 | 1 | 1 | |
| Ichneumonoidea | |||||
| Ichnuemonidae | 6 | 9 | 0 | 0 | |
| * Brachonidae | 6 | 12 | 2 | 2 | |
| Chalcidoidea | |||||
| Aphelinidae | 1 | 1 | 0 | 0 | |
| Chalcidae | 3 | 4 | 0 | 0 | |
| ** Encyrtidae | 6 | 31 | 0 | 0 | |
| Eupelmidae | 2 | 2 | 0 | 0 | |
| Eurytomidae | 1 | 3 | 0 | 0 | |
| ** Mymaridae | 6 | 36 | 3 | 4 | |
| ** Pteromalidae | 3 | 7 | 2 | 2 | |
| Torymidae | 1 | 2 | 0 | 0 | |
| Eulophidae | 0 | 0 | 0 | 0 | |
| Perilampidae | 0 | 0 | 0 | 0 | |
| Cynipoidea | |||||
| ** Figitidae | 3 | 22 | 1 | 1 | |
| Diaprioidea | |||||
| Diapriidae | 3 | 6 | 1 | 1 | |
| Ceraphronoidea | |||||
| Ceraphronidae | 2 | 33 | 0 | 0 | |
| Platygastroidea | |||||
| Platygastridae | 2 | 19 | 0 | 0 | |
| ** Scelionidae | 13 | 365 | 3 | 23 | |
| Formicoidea | |||||
| ** Formicidae | 11 | 789 | 5 | 28 | |
| Evanioidea | |||||
| Evaniidae | 1 | 1 | 0 | 0 | |
| Unknown | 4 | 8 | 1 | 1 | |
| TOTALS | 116 | 1490 | 25 | 74 | |
| Sorghum | Johnsongrass | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Pan | Sticky | Pan | Sticky | ||||||
| Superfamily | Family | Types | # | Types | # | Types | # | Types | # |
| Apoidea | |||||||||
| Halictidae | 12 | 638 | 5 | 28 | 9 | 68 | 6 | 13 | |
| Apidae | 9 | 11 | 0 | 0 | 2 | 2 | 0 | 0 | |
| Andrenidae | 1 | 6 | 0 | 0 | 3 | 3 | 0 | 0 | |
| Sphecidae | 19 | 42 | 3 | 5 | 5 | 10 | 0 | 0 | |
| Megachilidae | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | |
| Pompiloidea | |||||||||
| Pompilidae | 9 | 64 | 1 | 7 | 12 | 18 | 0 | 0 | |
| Mutillidae | 1 | 2 | 1 | 2 | 4 | 12 | 1 | 4 | |
| Chrysidoidea | |||||||||
| Chrysididae | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | |
| Bethylidae | 1 | 3 | 2 | 10 | 3 | 5 | 2 | 11 | |
| Dryinidae | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Tiphioidea | |||||||||
| Tiphiidae | 1 | 7 | 1 | 3 | 1 | 1 | 1 | 3 | |
| Sierolomorphidae | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | |
| Ichneumonoidea | |||||||||
| Ichnuemonidae | 1 | 1 | 2 | 2 | 5 | 7 | 2 | 2 | |
| Brachonidae | 4 | 7 | 5 | 19 | 3 | 3 | 3 | 11 | |
| Chalcidoidea | |||||||||
| Aphelinidae | 1 | 2 | 1 | 4 | 1 | 1 | 0 | 0 | |
| Chalcidae | 2 | 6 | 1 | 1 | 0 | 0 | 3 | 4 | |
| Encyrtidae | 4 | 9 | 6 | 107 | 1 | 4 | 6 | 28 | |
| Eupelmidae | 1 | 1 | 1 | 1 | 2 | 3 | 0 | 0 | |
| Eurytomidae | 1 | 1 | 2 | 2 | 0 | 0 | 1 | 3 | |
| Mymaridae | 1 | 2 | 4 | 55 | 0 | 0 | 6 | 40 | |
| Pteromalidae | 7 | 8 | 6 | 68 | 2 | 2 | 4 | 7 | |
| Torymidae | 0 | 0 | 1 | 10 | 0 | 0 | 1 | 2 | |
| Eulophidae | 2 | 2 | 2 | 1 | 0 | 0 | 0 | 0 | |
| Perilampidae | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 0 | |
| Cynipoidea | |||||||||
| Figitidae | 3 | 11 | 3 | 11 | 3 | 7 | 2 | 16 | |
| Diaprioidea | |||||||||
| Diapriidae | 2 | 12 | 2 | 4 | 3 | 5 | 2 | 2 | |
| Ceraphronoidea | |||||||||
| Ceraphronidae | 1 | 1 | 1 | 2 | 2 | 8 | 1 | 25 | |
| Platygastroidea | |||||||||
| Platygastridae | 0 | 0 | 3 | 24 | 1 | 1 | 2 | 18 | |
| Scelionidae | 10 | 89 | 11 | 466 | 12 | 76 | 11 | 312 | |
| Formicoidea | |||||||||
| Formicidae | 3 | 613 | 0 | 0 | 11 | 801 | 6 | 16 | |
| Evanioidea | |||||||||
| Evaniidae | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | |
| Unknown | 5 | ||||||||
| Unidentified | 1 | 3 | 3 | 6 | 1 | 4 | 3 | ||
| Total Families | 26 | 24 | 24 | 20 | |||||
| TOTAL | 100 | 1544 | 68 | 840 | 89 | 1044 | 65 | 523 | |
| Shannon-Weiner Diversity Index (Sorghum) | ||||
|---|---|---|---|---|
| Pan | Sticky | |||
| With Aphids | Control | With Aphids | Control | |
| 31 July 2019 | 1.914 | 0.910 | 2.403 | 1.991 |
| 17 August 2019 | 2.037 | 2.239 | 2.123 | 1.120 |
| 2 September 2019 | 2.147 | 1.526 | 2.100 | 0.674 |
| 9 September 2019 | 2.179 | 1.826 | 2.029 | 1.750 |
| 16 September 2019 | 2.252 | 1.229 | 2.513 | 2.594 |
| 27 September 2019 | 2.617 | 2.579 | 2.587 | 2.332 |
| Simpson Diversity Index (Sorghum) | ||||
| Pan | Sticky | |||
| With Aphids | Control | With Aphids | Control | |
| 31 July 2019 | 0.749 | 1.000 | 0.884 | 0.810 |
| 17 August 2019 | 0.780 | 0.871 | 0.732 | 0.612 |
| 2 September 2019 | 0.794 | 0.751 | 0.836 | 0.475 |
| 9 September 2019 | 0.767 | 0.772 | 0.739 | 0.810 |
| 16 September 2019 | 0.841 | 0.638 | 0.875 | 0.931 |
| 27 September 2019 | 0.840 | 0.953 | 0.868 | 0.890 |
| Shannon-Weiner Diversity Index (Johnsongrass) | ||||
| Pan | Sticky | |||
| With Aphids | Control | With Aphids | Control | |
| 2 August 2019 | 2.509 | 1.300 | 1.888 | 1.851 |
| 26 August 2019 | 2.259 | 1.600 | 2.082 | 1.709 |
| 6 September 2019 | 2.177 | N/A | 2.376 | N/A |
| 15 September 2019 | 2.039 | N/A | 2.695 | N/A |
| 23 September 2019 | 2.387 | N/A | 2.587 | N/A |
| Simpson Diversity Index (Johnsongrass) | ||||
| Pan | Sticky | |||
| With Aphids | Control | With Aphids | Control | |
| 2 August 2019 | 0.843 | 0.891 | 0.633 | 0.842 |
| 26 August 2019 | 0.827 | 0.875 | 0.749 | 0.838 |
| 6 September 2019 | 0.808 | N/A | 0.897 | N/A |
| 15 September 2019 | 0.781 | N/A | 0.903 | N/A |
| 23 September 2019 | 0.819 | N/A | 0.883 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris-Shultz, K.R.; Armstrong, J.S.; Caballero, M.; Hoback, W.W.; Knoll, J.E. Insect Feeding on Sorghum bicolor Pollen and Hymenoptera Attraction to Aphid-Produced Honeydew. Insects 2022, 13, 1152. https://doi.org/10.3390/insects13121152
Harris-Shultz KR, Armstrong JS, Caballero M, Hoback WW, Knoll JE. Insect Feeding on Sorghum bicolor Pollen and Hymenoptera Attraction to Aphid-Produced Honeydew. Insects. 2022; 13(12):1152. https://doi.org/10.3390/insects13121152
Chicago/Turabian StyleHarris-Shultz, Karen R., John Scott Armstrong, Michael Caballero, William Wyatt Hoback, and Joseph E. Knoll. 2022. "Insect Feeding on Sorghum bicolor Pollen and Hymenoptera Attraction to Aphid-Produced Honeydew" Insects 13, no. 12: 1152. https://doi.org/10.3390/insects13121152
APA StyleHarris-Shultz, K. R., Armstrong, J. S., Caballero, M., Hoback, W. W., & Knoll, J. E. (2022). Insect Feeding on Sorghum bicolor Pollen and Hymenoptera Attraction to Aphid-Produced Honeydew. Insects, 13(12), 1152. https://doi.org/10.3390/insects13121152