
Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Curculionidae) and Its Fungal Symbiont Ambrosiella roeperi Associated with Arecanut Kernel Decay in Karnataka, India

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
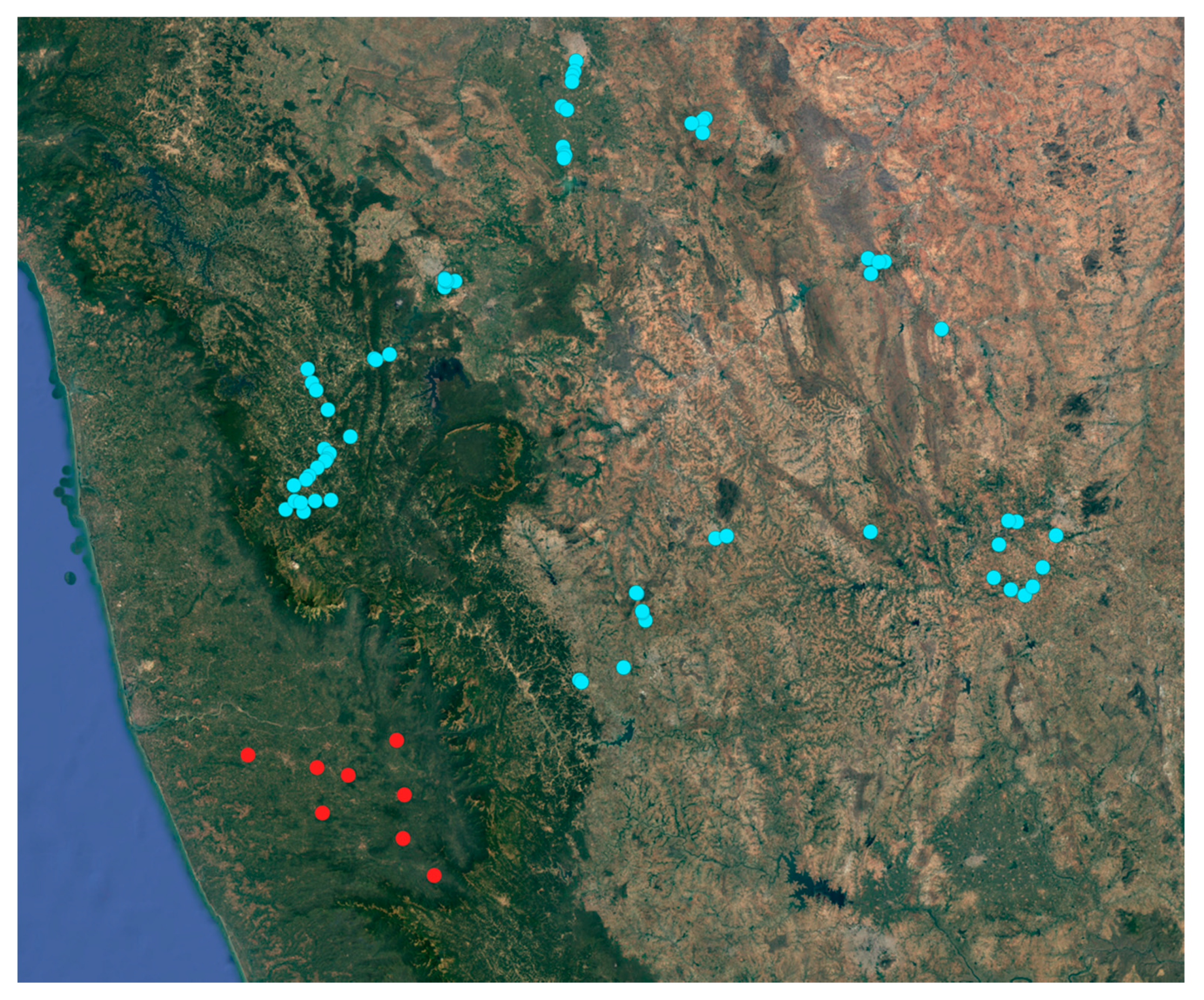
2.1. Sample Collection, Symptoms Documentation and Pest Distribution
2.2. Pest Identification
2.3. Symbiont Fungus Isolation and Characterization
3. Results
3.1. Ambrosia Beetle Identification
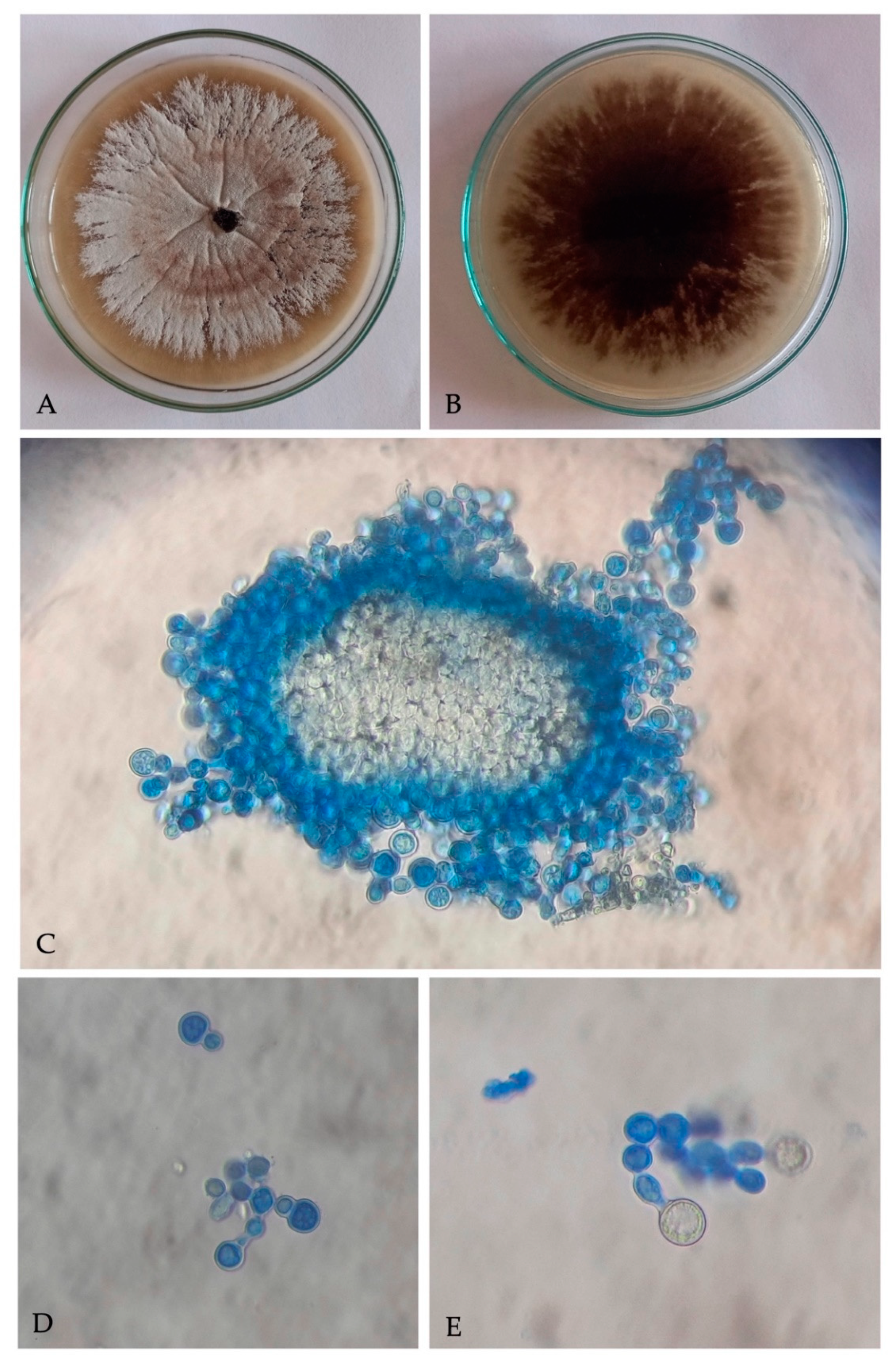
3.2. Symbiont Fungus Identification
3.3. Infestation Symptoms, Damage and Incidence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bright, D.E.; (Department of Agricultural Biology Colorado State University, Fort Collins, CO, USA). Personal communication, 2021.
- Dole, S.A.; Jordal, B.H.; Cognato, A.I. Polyphyly of Xylosandrus Reitter inferred from nuclear and mitochondrial genes (Coleoptera: Curculionidae: Scolytinae). Mol. Phylogenetics Evol. 2010, 54, 773–782. [Google Scholar] [CrossRef]
- Storer, C.; Payton, A.; McDaniel, S.; Jordal, B.; Hulcr, J. Cryptic genetic variation in an inbreeding and cosmopolitan pest, Xylosandrus crassiusculus, revealed using dd RAD seq. Ecol. Evol. 2017, 7, 10974–10986. [Google Scholar] [CrossRef] [Green Version]
- EPPO Database. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/alert_list (accessed on 10 September 2021).
- Ranger, C.M.; Reding, M.E.; Schultz, P.B.; Oliver, J.B.; Frank, S.D.; Addesso, K.M.; Chong, J.H.; Sampson, B.; Werle, C.; Gill, S.; et al. Biology, ecology, and management of non-native ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) in ornamental plant nurseries. J. Integr. Pest Manag. 2016, 7, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Beeson, C.F.C. The Ecology and Control of the Forest Insects of India and the Neighbouring Countries; Forest Research Institute: Dehra Dun, India, 1941; 1007p. [Google Scholar]
- Prathapan, K.D.; Hiremath, S.R. Post-flood outbreak of Xylosandrus crassiusculus and Diuncus corpulentus (Coleoptera: Curculionidae: Scolytinae: Xyleborini) on tree spices in Kerala. J. Spices Aromat. Crops 2018, 27, 161–166. [Google Scholar]
- Oregon State University. Available online: https://ir.library.oregonstate.edu/concern/conference_proceedings_or_journals/0v838255b (accessed on 21 November 2021).
- Ruzzier, E.; Prazaru, S.C.; Faccoli, M.; Duso, C. Xylosandrus germanus (Blandford, 1894) on Grapevines in Italy with a Compilation of World Scolytine Weevils Developing on Vitaceae. Insects 2021, 12, 869. [Google Scholar] [CrossRef]
- Schedl, K.E. Scolytidae und Platypodidae Afrikans. Band 2. Familie Scolytidae. Rev. De Entomol. De Mocamb. 1963, 5, 1–594. [Google Scholar]
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Michael, E.R. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, e0131496. [Google Scholar]
- Ranger, C.M.; Reding, M.E.; Addesso, C.; Ginzel, M.; Rassati, D. Semiochemical-mediated host selection by Xylosandrus spp. ambrosia beetles (Coleoptera: Curculionidae) attacking horticultural tree crops: A review of basic and applied science. Can. Entomol. 2021, 153, 103–120. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Ranger, C.M.; Rassati, D. Ambrosia beetle response to ethanol concentration and host tree species. J. Appl. Entomol. 2021, 145, 800–809. [Google Scholar] [CrossRef]
- Biedermann, P.H.W.; Vega, F.E. Ecology and evolution of insect-fungus mutualisms. Annu. Rev. Entomol. 2020, 65, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.C. Ecology and evolution of mycophagous bark beetles and their fungal partners. In Insect–Fungal Associations; Vega, F.E., Blackwell, M., Eds.; Oxford University Press: New York, NY, USA, 2005; pp. 257–291. [Google Scholar]
- Brader, L. Etude de la Relation Entre le Scolyte des Rameaux du Cafeier, Xyleborus Compactus Eichh. (X. morstatti Hag.), et sa Plante-Hote; Wageningen University: Wageningen, The Netherlands, 1964; Volume 64, pp. 1–109. [Google Scholar]
- Gebhardt, H.; Weiss, M.; Oberwinkler, F. Dryadomyces amasae: A nutritional fungus associated with ambrosia beetles of the genus Amasa (Coleoptera: Curculionidae, Scolytinae). Mycol. Res. 2005, 109, 687–696. [Google Scholar] [CrossRef]
- Kinuura, H. Symbiotic fungi associated with ambrosia beetles. Jpn. Agric. Res. Q. 1995, 29, 57–63. [Google Scholar]
- Hulcr, J.; Dunn, R.R. The sudden emergence of pathogenicity in insect-fungus symbioses threatens naive forest ecosystems. Proc. R. Soc. B Biol. Sci. 2011, 278, 2866–2873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, T.C.; McNew, D.; Chase, M.; Fraedrich, S.W.; Reed, S.E. Ambrosiella roeperi sp. nov. is the mycangial symbiont of the granulate ambrosia beetle, Xylosandrus crassiusculus. Mycologia 2014, 106, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Gugliuzzo, A.; Biedermann, P.H.W.; Carrillo, D.; Castrillo, L.A.; Egonyu, J.P.; Gallego, D.; Haddi, K.; Hulcr, J.; Jactel, H.; Kajimura, H.; et al. Recent advances toward the sustainable management of invasive Xylosandrus ambrosia beetles. J. Pest Sci. 2021, 94, 615–637. [Google Scholar] [CrossRef]
- Thube, S.H.; Mohan, C.; Pandian, R.T.P.; Saneera, E.K.; Sannagoudra, H.M.; Hegde, V.; Chowdappa, P. First record of the invasive neotropical ambrosia beetle Euplatypus parallelus (Fabricius, 1801) (Coleoptera: Curculionidae: Platypodinae) infesting arecanut in Karnataka, India. Coleopt. Bull. 2018, 72, 713–717. [Google Scholar] [CrossRef]
- Bhat, R.; Sujatha, S.; Balasimha, D. Impact of drip fertigation on productivity of arecanut (Areca catechu L.). Agric. Water Manag. 2007, 90, 101–111. [Google Scholar] [CrossRef]
- Arjungi, K.N. Areca nut: A review. Arzneim.-Forsch. 1976, 26, 951–957. [Google Scholar]
- Smith, S.M.; Beaver, R.A.; Cognato, A.I.; Hulcr, J.; Redford, A.J. Southeast Asian Ambrosia Beetle ID. USDA APHIS Identification Technology Program (ITP) and Michigan State University. Fort Collins, CO, USA. Available online: https://idtools.org/id/wbb/sea-ambrosia/ (accessed on 10 September 2021).
- University of Florida. Available online: https://journals.flvc.org/edis/article/view/119233 (accessed on 10 September 2021).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; Volume 1. [Google Scholar]
- Francke-Grosmann, H. Ectosymbiosis in wood-inhabiting insects. In Symbiosis; Henry, S.M., Ed.; Academic Press: New York, NY, USA, 1967; Volume 11, pp. 142–206. [Google Scholar]
- Pandian, R.T.P.; Bhat, A.I.; Biju, C.N.; Sasi, S. Development of diagnostic assays for rapid and sensitive detection of Phytophthora infecting major spices and plantation crops. J. Spices Aromat. Crops 2018, 27, 119–130. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for Phylogenetic. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 1–315. [Google Scholar]
- Kühnholz, S.; Borden, J.H.; Uzunovic, A. Secondary ambrosia beetles in apparently healthy trees: Adaptations, potential causes and suggested research. Integr. Pest Manag. Rev. 2001, 6, 209–219. [Google Scholar] [CrossRef]
- Chong, J.; Reid, L.; Williamson, M. Distribution, host plants, and damage of the black twig borer, Xylosandrus compactus (Eichhoff), in South Carolina. J. Agric. Urban Entomol. 2009, 26, 199–208. [Google Scholar] [CrossRef]
- Reding, M.E.; Ranger, C.M. Attraction of invasive ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) to ethanol-treated tree bolts. J. Econ. Entomol. 2020, 113, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.L. The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph; Brigham Young University: Provo, UT, USA, 1982; Volume 6. [Google Scholar]
- Carlton, C.; Bayless, V. A case of Cnestus mutilatus (Blandford) (Curculionidae: Scolytinae: Xyleborini) females damaging plastic fuel storage containers in Louisiana, USA. Coleopt. Bull. 2011, 65, 290–291. [Google Scholar] [CrossRef]
- Rajan, A.; Kurup, J.G.; Abraham, T.E. Biosoftening of arecanut fiber for value added products. Biochem. Eng. J. 2005, 25, 237–242. [Google Scholar] [CrossRef]
- Balanagouda, P.; Sridhara, S.; Shil, S.; Hegde, V.; Naik, M.K.; Narayanswamy, H.; Balsundram, K. Assessment of the spatial distribution and risk associated with fruit rot disease in Areca catechu L. J. Fungi 2021, 7, 797. [Google Scholar] [CrossRef]
- NIPHM. Available online: https://niphm.gov.in/IPMPackages/Arecanut.pdf (accessed on 10 September 2021).
- Devasahayam, S.; Nair, C.R. Insect pest management in arecanut. Indian Farming 1982, 32, 44–46. [Google Scholar]
- Rajashekharappa, M.T. Impact of Crop Losses Due to Pests and Diseases on Cost Structure of Arecanut Cultivation—An Economic Analysis in Traditional Malnad Region of Karnataka’. Master’s Thesis, University of Agricultural Sciences, Bengaluru, India, 2001. [Google Scholar]
- Williams, G.M.; Ginzel, M.D. Spatial and climatic factors influence ambrosia beetle (Coleoptera: Curculionidae) abundance in intensively managed plantations of eastern black walnut. Environ. Entomol. 2020, 49, 49–58. [Google Scholar] [CrossRef]
- Urvois, T.; Auger-Rozenberg, M.A.; Roques, A.; Rossi, J.P.; Kerdelhue, C. Climate change impact on the potential geographical distribution of two invading Xylosandrus ambrosia beetles. Sci. Rep. 2021, 11, 1339. [Google Scholar] [CrossRef] [PubMed]
- Thube, S.H.; Pandian, R.T.P.; Bhavishya, E.K.; Mohan, C.; Nagaraja, N.R. Major storage insect pests of arecanut Areca catechu L.—A Survey. J. Entomol. Zool. Stud. 2017, 5, 1471–1475. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
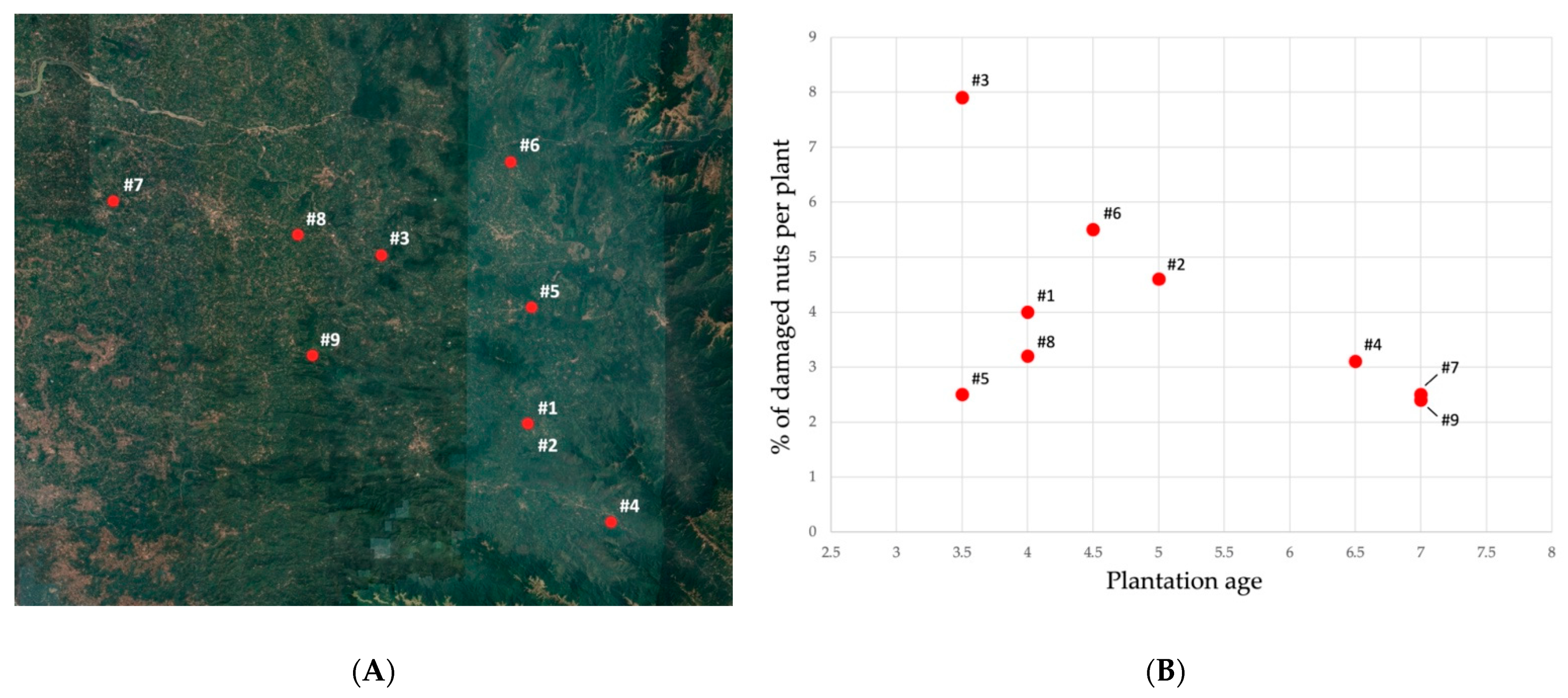
| Location | Palms Age (Years) | n° Infested Palms | n° Damaged Nuts/Total Nuts on Infested Palms | % of Damaged Nuts on Infested Palms | % of Damaged Nuts |
|---|---|---|---|---|---|
| 12°34.487′ 75°29.412′ | 4 | 7 | 11/272 | 4 | 0.05 |
| 12°34.502′ 75°29.392′ | 5 | 12 | 18/384 | 4.6 | 0.08 |
| 12°43.563′ 75°21.334′ | 3.5 | 27 | 68/856 | 7.9 | 0.31 |
| 12°29.201′ 75°33.987′ | 6.5 | 2 | 3/95 | 3.1 | 0.01 |
| 12°40.758′ 75°29.604′ | 3.5 | 11 | 8/320 | 2.5 | 0.04 |
| 12°48.570′ 75°28.458′ | 4.5 | 2 | 6/109 | 5.5 | 0.04 |
| 12°46.472′ 75°06.575′ | 7 | 12 | 14/555 | 2.5 | 0.06 |
| 12°44.664′ 75°16.736′ | 4 | 5 | 6/185 | 3.2 | 0.03 |
| 12°38.160′ 75°17.535′ | 7 | 6 | 6/244 | 2.4 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thube, S.H.; Pandian, T.P.; Bhavishya, A.; Babu, M.; Josephrajkumar, A.; Chaithra, M.; Hegde, V.; Ruzzier, E. Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Curculionidae) and Its Fungal Symbiont Ambrosiella roeperi Associated with Arecanut Kernel Decay in Karnataka, India. Insects 2022, 13, 67. https://doi.org/10.3390/insects13010067
Thube SH, Pandian TP, Bhavishya A, Babu M, Josephrajkumar A, Chaithra M, Hegde V, Ruzzier E. Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Curculionidae) and Its Fungal Symbiont Ambrosiella roeperi Associated with Arecanut Kernel Decay in Karnataka, India. Insects. 2022; 13(1):67. https://doi.org/10.3390/insects13010067
Chicago/Turabian StyleThube, Shivaji Hausrao, Thava Prakasa Pandian, Anthara Bhavishya, Merin Babu, Arulappan Josephrajkumar, Muddumadiah Chaithra, Vinayaka Hegde, and Enrico Ruzzier. 2022. "Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Curculionidae) and Its Fungal Symbiont Ambrosiella roeperi Associated with Arecanut Kernel Decay in Karnataka, India" Insects 13, no. 1: 67. https://doi.org/10.3390/insects13010067
APA StyleThube, S. H., Pandian, T. P., Bhavishya, A., Babu, M., Josephrajkumar, A., Chaithra, M., Hegde, V., & Ruzzier, E. (2022). Xylosandrus crassiusculus (Motschulsky) (Coleoptera: Curculionidae) and Its Fungal Symbiont Ambrosiella roeperi Associated with Arecanut Kernel Decay in Karnataka, India. Insects, 13(1), 67. https://doi.org/10.3390/insects13010067