
Ephemeroptera (Mayflies) Assemblages and Environmental Variation along Three Streams Located in the Dry-Hot Valleys of Baima Snow Mountain, Yunnan, Southwest China

 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sampling and Lab Analysis
2.2. Data Analysis
3. Results
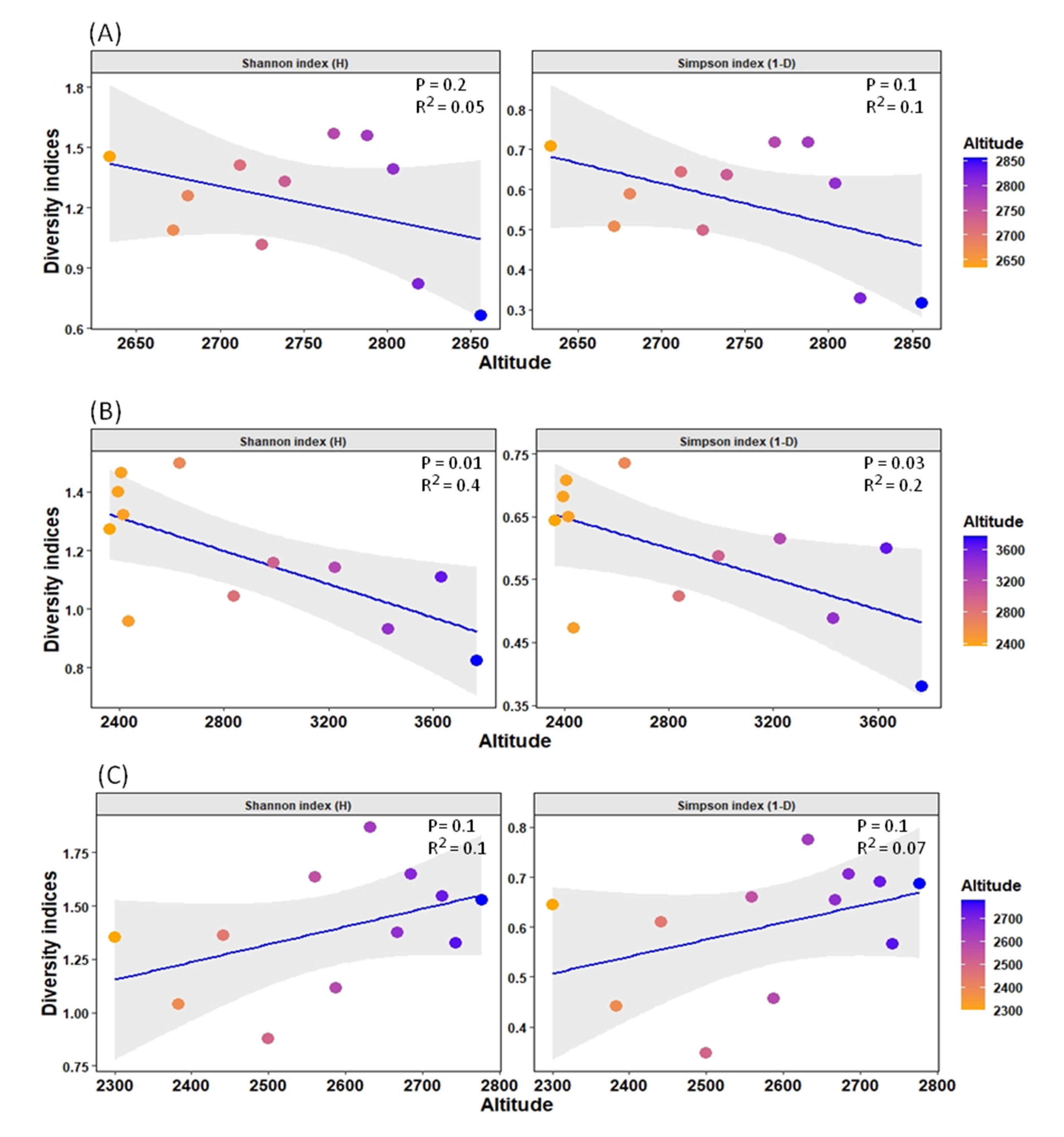
3.1. Environmental Variation across the Three Streams in BSM
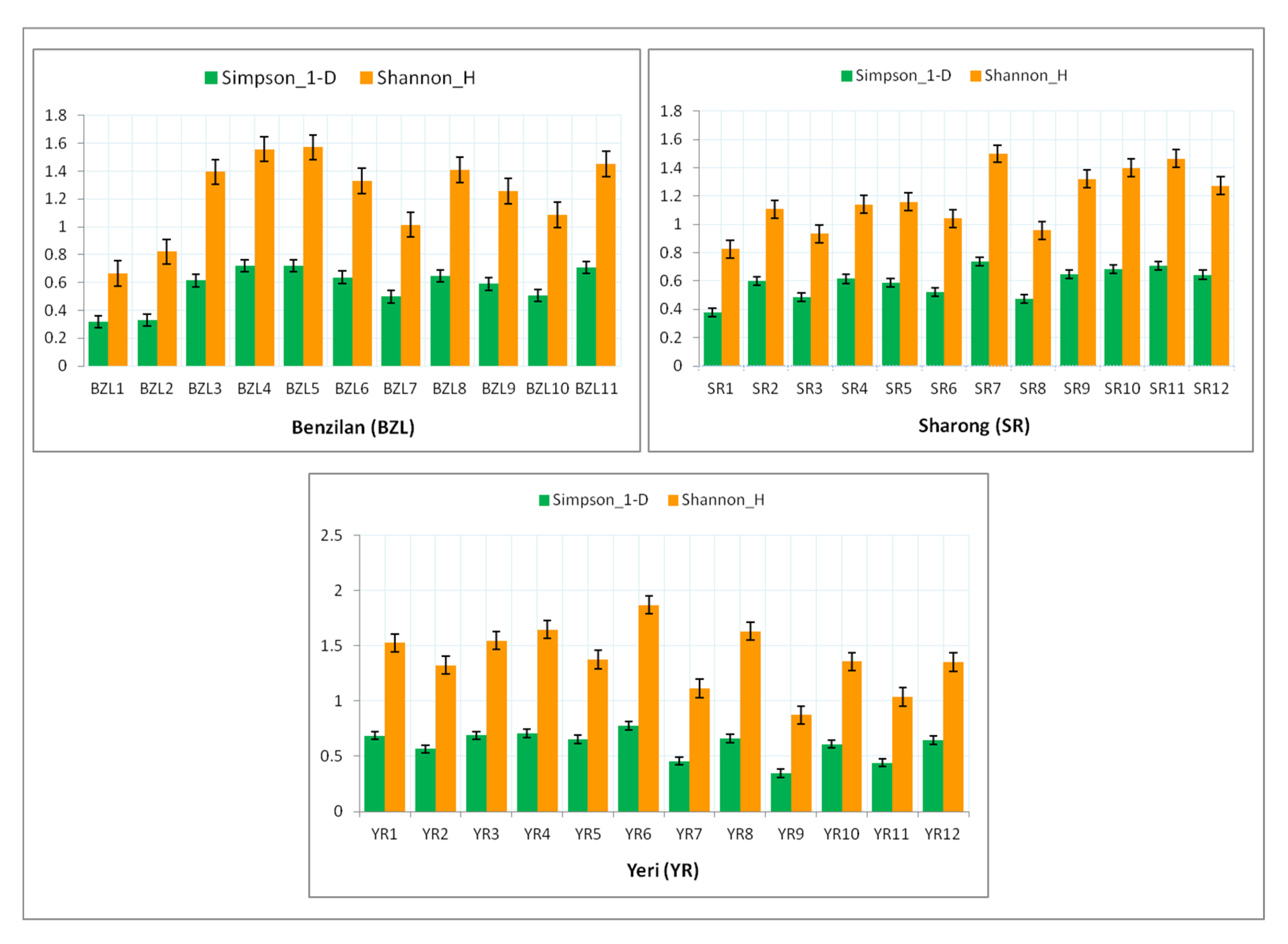
3.2. Distribution, Structure, and Composition of Mayfly Assemblages
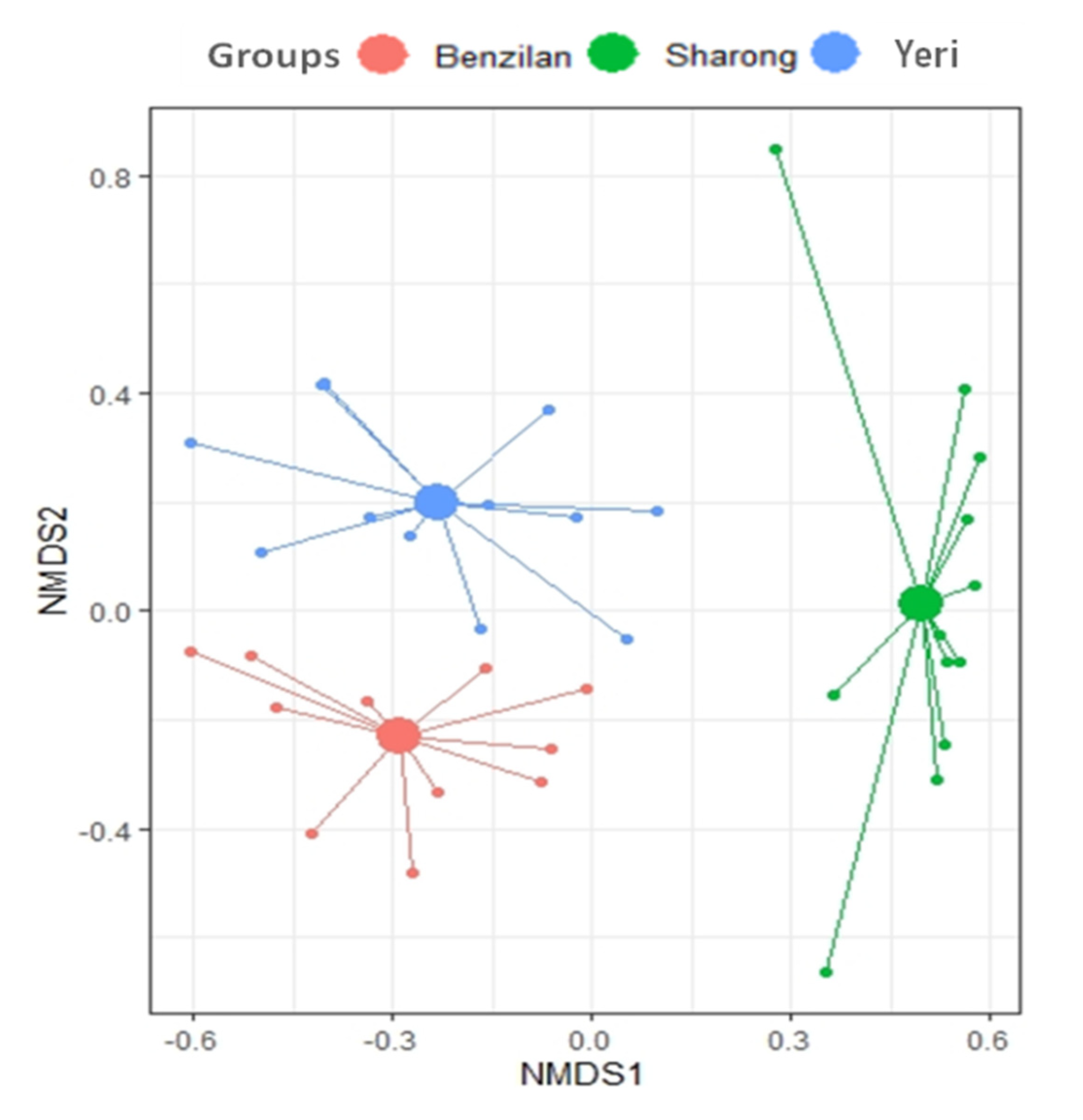
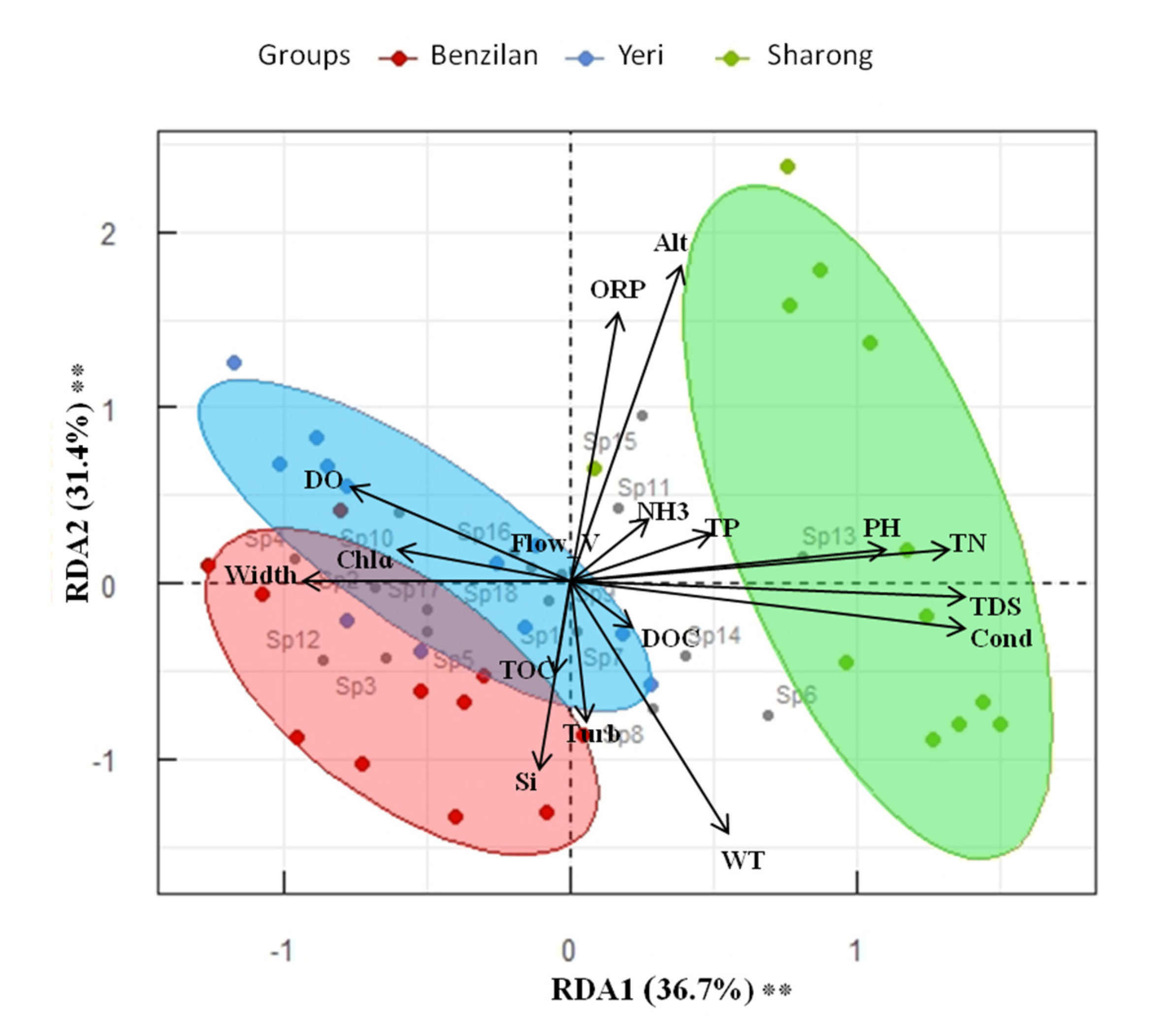
3.3. Distribution Pattern and Relationship between Ephemeropterans and Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rogora, M.; Frate, L.; Carranza, M.L.; Freppaz, M.; Stanisci, A.; Bertani, I.; Bottarin, R.; Brambilla, A.; Canullo, R.; Carbognani, M.; et al. Assessment of climate change effects on mountain ecosystems through a cross-site analysis in the Alps and Apennines. Sci. Total. Environ. 2018, 624, 1429–1442. [Google Scholar] [CrossRef]
- Arimoro, F.O.; Muller, W.J. Mayfly (Insecta: Ephemeroptera) community structure as an indicator of the ecological status of a stream in the Niger Delta area of Nigeria. Environ. Monit. Assess. 2009, 166, 581–594. [Google Scholar] [CrossRef]
- Sartori, M.; Brittain, J.E. Order Ephemeroptera. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology, 4th ed.; Thorp, J., Rodgers, D.C., Eds.; Academic Press: New York, NY, USA, 2015; pp. 873–891. [Google Scholar] [CrossRef]
- Alhejoj, I.; Salameh, E.; Bandel, K. Mayflies (order ephemeroptera): An effective indicator of water bodies conditions in Jordan. Int. J. Sci. Res. Environ. Sci. 2014, 2, 361–370. [Google Scholar] [CrossRef][Green Version]
- Baptista, D.F.; Buss, D.F.; Dias, L.G.; Nessimian, J.; Da Silva, E.R.; Neto, A.H.D.M.; De Carvalho, S.N.; de Oliveira, M.; Andrade, L. Functional feeding groups of Brazilian Ephemeroptera nymphs: Ultrastructure of mouthparts. Ann. Limnol. Int. J. Limnol. 2006, 42, 87–96. [Google Scholar] [CrossRef]
- Brittain, J.E. Biology of Mayflies. Annu. Rev. Èntomol. 1982, 27, 119–147. [Google Scholar] [CrossRef]
- Landa, V.; Soldán, T. The possibilities of mayfly faunistics to indicate environmental changes of large areas. In Overview and Strategies of Ephemeroptera and Plecoptera; Alba-Tercedor, J., Sánchez-Ortega, A., Eds.; Sandhill Crane Press: Gainesville, FL, USA, 1991; pp. 559–565. [Google Scholar]
- Bae, Y.J. Current research and methods in aquatic entomology with special reference to mayfly studies. In 92 Symposium Proceedings of the Korean Society of Limnology; 1992; pp. 9–32. [Google Scholar]
- Park, S.; Bae, Y.; Yoon, I. Revision of the aetidac (Ephcmeroptera) of Korea (1) Historical Review, Acentrella Berigtsson and Balcticlla Uéno. Entomol. Res. Bull. 1996, 22, 55–66. [Google Scholar]
- Vilenica, M.; Ivković, M.; Sartori, M.; Mihaljević, Z. Mayfly emergence along an oligotrophic Dinaric karst hydrosystem: Spatial and temporal patterns, and species–environment relationship. Aquat. Ecol. 2017, 51, 417–433. [Google Scholar] [CrossRef]
- Ramulifho, P.; Rivers-Moore, N.; Dallas, H.; Foord, S. A conceptual framework towards more holistic freshwater conservation planning through incorporation of stream connectivity and thermal vulnerability. J. Hydrol. 2018, 556, 173–181. [Google Scholar] [CrossRef]
- Bauernfeind, E.; Moog, O. Mayflies (Insecta: Ephemeroptera) and the assessment of ecological integrity: A methodological approach. Assess. Ecol. Integr. Run. Waters 2000, 423, 71–83. [Google Scholar] [CrossRef]
- Popielarz, P.A.; Neal, Z.P. The Niche as a Theoretical Tool. Annu. Rev. Sociol. 2007, 33, 65–84. [Google Scholar] [CrossRef]
- Heino, J. Does dispersal ability affect the relative importance of environmental control and spatial structuring of littoral macroinvertebrate communities? Oecologia 2012, 171, 971–980. [Google Scholar] [CrossRef]
- Axelsson, E.P.; Hjältén, J.; Leroy, C.J.; Whitham, T.G.; Julkunen-Tiitto, R.; Wennström, A. Leaf litter from insect-resistant transgenic trees causes changes in aquatic insect community composition. J. Appl. Ecol. 2011, 48, 1472–1479. [Google Scholar] [CrossRef]
- Dolný, A.; Harabiš, F.; Bárta, D.; Lhota, S.; Drozd, P. Aquatic insects indicate terrestrial habitat degradation: Changes in taxonomical structure and functional diversity of dragonflies in tropical rainforest of East Kalimantan. Trop. Zool. 2012, 25, 141–157. [Google Scholar] [CrossRef]
- Ligeiro, R.; Hughes, R.M.; Kaufmann, P.R.; Macedo, D.R.; Firmiano, K.R.; Ferreira, W.R.; Oliveira, D.; Melo, A.S.; Callisto, M. Defining quantitative stream disturbance gradients and the additive role of habitat variation to explain macroinvertebrate taxa richness. Ecol. Indic. 2013, 25, 45–57. [Google Scholar] [CrossRef]
- Souza, H.M.D.L.; Cabette, H.S.R.; Juen, L. Baetidae (Insecta, Ephemeroptera) em córregos do cerrado matogrossense sob diferentes níveis de preservação ambiental. Iheringia Série Zool. 2011, 101, 181–190. [Google Scholar] [CrossRef]
- Monteiro, C.D.S., Jr.; Couceiro, S.R.M.; Hamada, N.; Juen, L. Effect of vegetation removal for road building on richness and composition of Odonata communities in Amazonia, Brazil. Int. J. Odonatol. 2013, 16, 135–144. [Google Scholar] [CrossRef]
- Moog, O. Fauna Aquatica Austriaca, Edition 2002, Wassserwirtschaftskataster, Bundesministerium für Land und Forstwirtschaft; Umwelt und Wasserwirtschaft: Vienna, Austria, 2002. [Google Scholar]
- Allan, J.D.; Castillo, M.M. Stream Ecology: Structure and Function of Running Waters; Chapman–Hall: New York, NY, USA, 2007. [Google Scholar]
- Choudhary, J.; Singh, S.N. Physico-Chemical and Biological Parameters of the Three Rural Ponds of Sasaram of Bihar. Int. J. Appl. Sci. Biotechnol. 2014, 2, 206–210. [Google Scholar] [CrossRef]
- Karr, J.R. Assessment of Biotic Integrity Using Fish Communities. Fisheries 1981, 6, 21–27. [Google Scholar] [CrossRef]
- Primack, R.B.; Rodrigues, E. Biologia da Conservação; Planta: Lodrina, Brazil, 2001. [Google Scholar]
- Shimano, Y.; Salles, F.F.; Juen, L. Study of the mayfly order Ephemeroptera (Insecta) in Brazil: A scienciometric review. Rev. Bras. Èntomol. 2013, 57, 359–364. [Google Scholar] [CrossRef]
- Shimano, Y.; Juen, L.; Salles, F.F.; Nogueira, D.; Cabette, H.S.R. Environmental and spatial processes determining Ephemeroptera (Insecta) structures in tropical streams. Ann. Limnol. Int. J. Limnol. 2013, 49, 31–41. [Google Scholar] [CrossRef]
- Fischer, A.; Blaschke, M.; Bässler, C. Altitudinal gradients in biodiversity research: The state of the art and future perspectives under climate change aspects. Wald. Online 2011, 11, 35–47. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, Q.; Wen, Z.; Xia, L.; Zhang, Q.; Zhou, H. What drives the species richness patterns of non-volant small mammals along a subtropical elevational gradient? Ecography 2012, 36, 185–196. [Google Scholar] [CrossRef]
- Wen, Z.; Wu, Y.; Du, Y.; Xia, L.; Ge, D.; Yang, Q.; Chen, L. Seasonal Change of Species Diversity Patterns of Non-volant Small Mammals along Three Subtropical Elevational Gradients. Biotropica 2014, 46, 479–488. [Google Scholar] [CrossRef]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1994; pp. 1–570. [Google Scholar]
- Jackson, J.K.; Dudgeon, D. Tropical Asian Streams: Zoobenthos, Ecology and Conservation. David Dudgeon. J. N. Am. Benthol. Soc. 1999, 18, 427–428. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 26 August 2021).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.5-1. 2018. Available online: http://CRAN.R-project.org/package=vegan (accessed on 26 August 2021).
- Bertuzzo, E.; Carrara, F.; Mari, L.; Altermatt, F.; Rodriguez-Iturbe, I.; Rinaldo, A. Geomorphic controls on elevational gradients of species richness. Proc. Natl. Acad. Sci. USA 2016, 113, 1737–1742. [Google Scholar] [CrossRef]
- Ruggiero, A.; Hawkins, B.A. Why do mountains support so many species of birds? Ecography 2008, 31, 306–315. [Google Scholar] [CrossRef]
- Huan-Cheng, M.; McConchie, J.A. The dry-hot valleys and forestation in southwest China. J. For. Res. 2001, 12, 35–39. [Google Scholar] [CrossRef]
- Svitok, M. Structure and spatial variability of mayfly (Ephemeroptera) communities in the upper Hron River basin. Biologia 2006, 61, 547–554. [Google Scholar] [CrossRef][Green Version]
- Kłonowska-Olejnik, M.; Skalski, T. The effect of environmental factors on the mayfly communities of headwater streams in the Pieniny Mountains (West Carpathians). Biologia 2014, 69, 498–507. [Google Scholar] [CrossRef]
- Chapman, D.V. (Ed.) Water Quality Assessments: A Guide to the Use of Biota, Sediments and Water in Environmental Monitoring; CRC Press: Boca Raton, FL, USA, 1996. [Google Scholar]
- Hrovat, M.; Urbanič, G.; Sivec, I. Community structure and distribution of Ephemeroptera and Plecoptera larvae in lowland karst rivers in Slovenia. Aquat. Insects 2009, 31, 343–357. [Google Scholar] [CrossRef]
- Vilenica, M.; Previšić, A.; Ivković, M.; Popijač, A.; Vučković, I.; Kučinić, M.; Kerovec, M.; Gattolliat, J.-L.; Sartori, M.; Mihaljević, Z. Mayfly (Insecta: Ephemeroptera) assemblages of a regulated perennial Mediterranean river system in the Western Balkans. Biologia 2016, 71, 1038–1048. [Google Scholar] [CrossRef]
- Rueda, J.; Camacho, A.; Mezquita, F.; Hernández, R.; Roca, J.R. Effect of episodic and regular sewage discharges on the water chemistry and macroinvertebrate fauna of a Mediterranean stream. Water Air Soil Pollut. 2002, 140, 425–444. [Google Scholar] [CrossRef]
- Buss, D.F.; Salles, F.F. Using Baetidae Species as Biological Indicators of Environmental Degradation in a Brazilian River Basin. Environ. Monit. Assess. 2006, 130, 365–372. [Google Scholar] [CrossRef]
- Olson, J.R.; Hawkins, C.P. Effects of total dissolved solids on growth and mortality predict distributions of stream macroinvertebrates. Freshw. Biol. 2017, 62, 779–791. [Google Scholar] [CrossRef]
- Vilenica, M.; Stanković, V.M.; Sartori, M.; Kučinić, M.; Mihaljević, Z. Environmental factors affecting mayfly assemblages in tufa-depositing habitats of the Dinaric Karst. Knowl. Manag. Aquat. Ecosyst. 2017, 418, 14. [Google Scholar] [CrossRef][Green Version]
- Haidekker, A.; Hering, D. Relationship between benthic insects (Ephemeroptera, Plecoptera, Coleoptera, Trichoptera) and temperature in small and medium-sized streams in Germany: A multivariate study. Aquat. Ecol. 2007, 42, 463–481. [Google Scholar] [CrossRef]
- Pond, G.J. Patterns of Ephemeroptera taxa loss in Appalachian headwater streams (Kentucky, USA). Hydrobiologia 2010, 641, 185–201. [Google Scholar] [CrossRef]
- Pond, G.J.; Passmore, M.E.; Borsuk, F.A.; Reynolds, L.; Rose, C.J. Downstream effects of mountaintop coal mining: Comparing biological conditions using family- and genus-level macroinvertebrate bioassessment tools. J. N. Am. Benthol. Soc. 2008, 27, 717–737. [Google Scholar] [CrossRef]
- Ramulifho, P.A.; Foord, S.H.; Rivers-Moore, N.A. The role of hydro-environmental factors in Mayfly (Ephemeroptera, Insecta) community structure: Identifying threshold responses. Ecol. Evol. 2020, 10, 6919–6928. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y.; Kim, P.-J.; Hwang, S.-J.; Lee, J.-K.; Lee, S.-W.; Park, C.-H.; Moon, J.-S.; Kong, D.-S. Korean stream types based on benthic macroinvertebrate communities according to stream size and altitude. J. Freshw. Ecol. 2017, 32, 741–759. [Google Scholar] [CrossRef]
- Helson, J.E.; Williams, D.D.; Turner, D. Larval Chironomid Community Organization in Four Tropical Rivers: Human Impacts and Longitudinal Zonation. Hydrobiologia 2006, 559, 413–431. [Google Scholar] [CrossRef]
- Jacobus, L.M.; Macadam, C.R.; Sartori, M. Mayflies (Ephemeroptera) and Their Contributions to Ecosystem Services. Insects 2019, 10, 170. [Google Scholar] [CrossRef]
- Jacobsen, D. Low oxygen pressure as a driving factor for the altitudinal decline in taxon richness of stream macroinvertebrates. Oecologia 2007, 154, 795–807. [Google Scholar] [CrossRef]
- Verberk, W.C.; Durance, I.; Vaughan, I.; Ormerod, S.J. Field and laboratory studies reveal interacting effects of stream oxygenation and warming on aquatic ectotherms. Glob. Chang. Biol. 2016, 22, 1769–1778. [Google Scholar] [CrossRef]
- Pringle, C.M.; Ramírez, A. Use of both benthic and drift sampling techniques to assess tropical stream invertebrate communities along an altitudinal gradient, Costa Rica. Freshw. Biol. 1998, 39, 359–373. [Google Scholar] [CrossRef]
- Tate, C.; Heiny, J. The ordination of benthic invertebrate communities in the South Platte River Basin in relation to environmental factors. Freshw. Biol. 1995, 33, 439–454. [Google Scholar] [CrossRef]
- Timm, H. Ephemeroptera and Plecoptera Larvae as Environmental Indicators in Running Waters of Estonia. Ephemeroptera and Plecoptera: Biology-ecology-systematics. In Proceedings of the 8th International Conference Ephemeroptera, Lausanne, Switzerland, 4–20 August 1995; Mauron+Tinguely & Lacht SA: Fribourg, Switzerland, 1995; pp. 247–253. [Google Scholar]
- Menetrey, N.; Oertli, B.; Sartori, M.; Wagner, A.; Lachavanne, J.B. Eutrophication: Are mayflies (Ephemeroptera) good bioindicators for ponds? Hydrobiologia 2007, 597, 125–135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | Benzilan | Sharong | Yeri | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Mean ± Std | Max | Min | Mean ± Std | Max | Min | Mean ± Std | |
| Alt (m) ns | 2856 | 2634 | 2743.27 ± 75.8 | 3769 | 2364 | 2878.5 ± 520.95 | 2778 | 2300 | 2583.75 ± 151.21 |
| Cond (µs/cm) *** | 305.0 | 84.1 | 159.064 ± 99.6 | 287.3 | 172 | 254.958 ± 36.83 | 170.0 | 139.4 | 151.392 ± 10.21 |
| TDS *** | 247.6 | 75.40 | 134.963 ± 80.3 | 250.9 | 167.7 | 219.158 ± 23.66 | 142.35 | 126.10 | 133.466 ± 5.07 |
| WT (°C) ns | 14.6 | 10.2 | 12.082 ± 1.44 | 15.6 | 7.5 | 12.000 ± 2.87 | 13.3 | 10.1 | 11.200 ± 1.25 |
| DO (mg/L) ns | 8.39 | 6.98 | 7.530 ± 0.40 | 7.78 | 6.94 | 7.398 ± 0.31 | 8.20 | 7.27 | 7.665 ± 0.29 |
| pH *** | 8.86 | 7.91 | 8.437 ± 0.27 | 8.96 | 8.62 | 8.863 ± 0.08 | 8.91 | 8.11 | 8.678 ± 0.21 |
| ORP ns | 132.2 | 75.3 | 97.727 ± 21.11 | 138.1 | 62.1 | 102.058 ± 27.87 | 115.2 | 36.6 | 85.642 ± 25.08 |
| TN (mg/L) *** | 0.15 | 0.09 | 0.118 ± 0.02 | 0.39 | 0.16 | 0.260 ± 0.06 | 0.22 | 0.18 | 0.194 ± 0.01 |
| NH3 (mg/L) ns | 0.019 | 0.004 | 0.008 ± 0.00 | 0.018 | 0.005 | 0.009 ± 0.00 | 0.019 | 0.03 | 0.008 ± 0.00 |
| TP (mg/L) ns | 5.006 | 3.754 | 4.256 ± 0.55 | 4.335 | 2.719 | 3.548 ± 0.57 | 3.512 | 3.033 | 3.173 ± 0.15 |
| Si (mg/L) *** | 0.034 | 0.016 | 0.022 ± 00.00 | 0.118 | 0.014 | 0.030 ± 0.02 | 0.026 | 0.013 | 0.015 ± 0.00 |
| DOC (mg/L) * | 1.58 | 0.48 | 1.103 ± 0.35 | 2.02 | 0.71 | 0.975 ± 0.35 | 1.08 | 0.58 | 0.780 ± 0.14 |
| Chla ns | 0.078 | 0.031 | 0.493 ± 0.01 | 0.189 | 0.000 | 0.031 ± 0.05 | 1.183 | 0.009 | 0.124 ± 0.33 |
| TOC (mg/L) *** | 3.060 | 0.943 | 1.373 ± 0.57 | 2.31 | 0.54 | 0.875 ± 0.52 | 0.765 | 0.485 | 0.582 ± 0.07 |
| Width (m) *** | 4.5 | 1.8 | 2.982 ± 0.84 | 2 | 2 | 2.00 ± 0.00 | 5.0 | 2.5 | 3.708 ± 0.75 |
| Flow-v (m/s) ns | 1.00 | 0.32 | 0.574 ± 0.18 | 0.80 | 0.44 | 0.558 ± 0.10 | 0.74 | 0.24 | 0.471 ± 0.15 |
| Turb (NTU) ns | 2.8 | 0.5 | 1.573 ± 0.66 | 5.3 | 0.0 | 1.200 ± 1.70 | 1.8 | 0.0 | 0.983 ± 0.67 |
| Families | Taxa | Benzilan | Sharong | Yeri | Total |
|---|---|---|---|---|---|
| Neoephemeridae | Potamanthellus edmundsi | 1 | 0 | 1 | 2 |
| Ephemerellidae | Ephemerlla sp. | 2 | 0 | 16 | 45 |
| Ephemerellidae | Serratella sp. | 36 | 0 | 11 | 47 |
| Ephemerellidae | Drunella sp. | 190 | 0 | 209 | 399 |
| Baetidae | Baetis sp. | 2821 | 1098 | 1806 | 5725 |
| Baetidae | Nigrobaetis (Takobia) sp. | 330 | 282 | 174 | 786 |
| Baetidae | Baetiella marginata | 701 | 548 | 232 | 1481 |
| Baetidae | Baetiella spathae | 174 | 43 | 75 | 292 |
| Siphlonuridae | Ameletus sp. | 3 | 1 | 0 | 4 |
| Leptophlebiidae | Habrophlebiodes sp | 7 | 1 | 306 | 314 |
| Caenidae | Caenis sp. | 0 | 1 | 0 | 1 |
| Heptageniidae | Iron sp. | 206 | 14 | 74 | 294 |
| Heptageniidae | Epeorus sp. | 12 | 116 | 59 | 187 |
| Heptageniidae | Ecdyonurus sp. | 21 | 46 | 31 | 98 |
| Heptageniidae | Rhithrogena sp1. | 30 | 820 | 109 | 959 |
| Heptageniidae | Rhithrogena sp2. | 1 | 1 | 4 | 6 |
| Ephemeridae | Ephemera sp. | 8 | 0 | 9 | 17 |
| Ephemeridae | Ephemera wuchowensis | 0 | 0 | 1 | 1 |
| Total | 4570 | 2971 | 3117 | 10,658 |
| Taxon | Average Dissimilarity | Percentage Contribution | Cumulative Percentage |
|---|---|---|---|
| Baetis sp. | 21.01 | 39.21 | 39.21 |
| Baetiella marginata | 8.552 | 15.96 | 55.17 |
| Rhithrogena sp1 | 7.106 | 13.26 | 68.43 |
| Nigrobaetis (Takobia) sp. | 3.762 | 7.021 | 75.45 |
| Habrophlebiodes sp. | 3.121 | 5.825 | 81.28 |
| Drunella sp. | 2.988 | 5.577 | 86.85 |
| Iron sp. | 2.156 | 4.025 | 90.88 |
| Baetiella spathae | 1.873 | 3.495 | 94.37 |
| Epeorus sp. | 1.198 | 2.236 | 96.61 |
| Ecdyonurus sp. | 0.707 | 1.321 | 97.93 |
| Ephemerlla sp. | 0.420 | 0.784 | 98.71 |
| Serratella sp. | 0.375 | 0.700 | 99.42 |
| Ephemera sp. | 0.147 | 0.274 | 99.69 |
| Rhithrogena sp2 | 0.055 | 0.103 | 99.79 |
| Ameletus sp. | 0.032 | 0.061 | 99.92 |
| Potamanthellus edmundsi | 0.025 | 0.048 | 99.97 |
| Ephemera wuchowensis | 0.009 | 0.016 | 99.98 |
| Caenis sp. | 0.008 | 0.015 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, M.; Li, X.; Tan, L.; Fornacca, D.; Li, Y.; Cili, N.; Tian, Z.; Yang, L.; Deng, X.; Liu, S.; et al. Ephemeroptera (Mayflies) Assemblages and Environmental Variation along Three Streams Located in the Dry-Hot Valleys of Baima Snow Mountain, Yunnan, Southwest China. Insects 2021, 12, 775. https://doi.org/10.3390/insects12090775
Farooq M, Li X, Tan L, Fornacca D, Li Y, Cili N, Tian Z, Yang L, Deng X, Liu S, et al. Ephemeroptera (Mayflies) Assemblages and Environmental Variation along Three Streams Located in the Dry-Hot Valleys of Baima Snow Mountain, Yunnan, Southwest China. Insects. 2021; 12(9):775. https://doi.org/10.3390/insects12090775
Chicago/Turabian StyleFarooq, Muhammad, Xianfu Li, Lu Tan, Davide Fornacca, Yanpeng Li, Nima Cili, Zhen Tian, Lu Yang, Xiaoling Deng, Shuoran Liu, and et al. 2021. "Ephemeroptera (Mayflies) Assemblages and Environmental Variation along Three Streams Located in the Dry-Hot Valleys of Baima Snow Mountain, Yunnan, Southwest China" Insects 12, no. 9: 775. https://doi.org/10.3390/insects12090775
APA StyleFarooq, M., Li, X., Tan, L., Fornacca, D., Li, Y., Cili, N., Tian, Z., Yang, L., Deng, X., Liu, S., & Xiao, W. (2021). Ephemeroptera (Mayflies) Assemblages and Environmental Variation along Three Streams Located in the Dry-Hot Valleys of Baima Snow Mountain, Yunnan, Southwest China. Insects, 12(9), 775. https://doi.org/10.3390/insects12090775