
Nectar Feeding by a Honey Bee’s Hairy Tongue: Morphology, Dynamics, and Energy-Saving Strategies



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
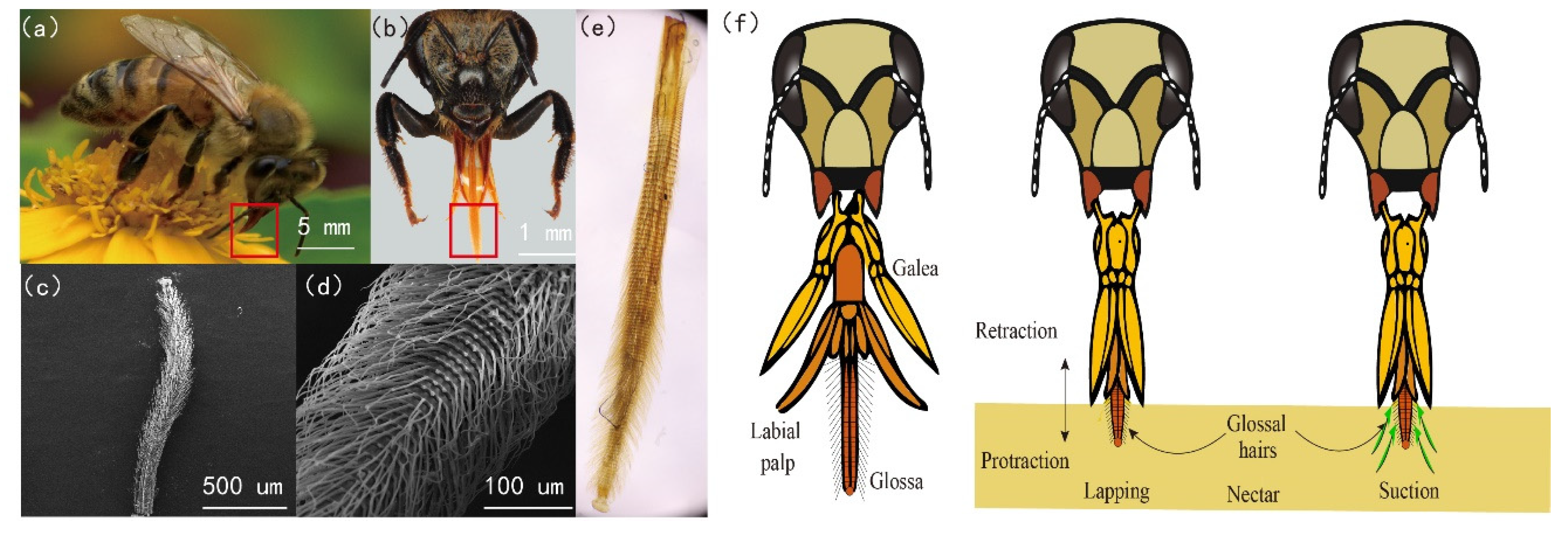
2. Honey Bee Mouth Parts Morphology
3. Feeding Behaviour of a Honey Bee Glossa
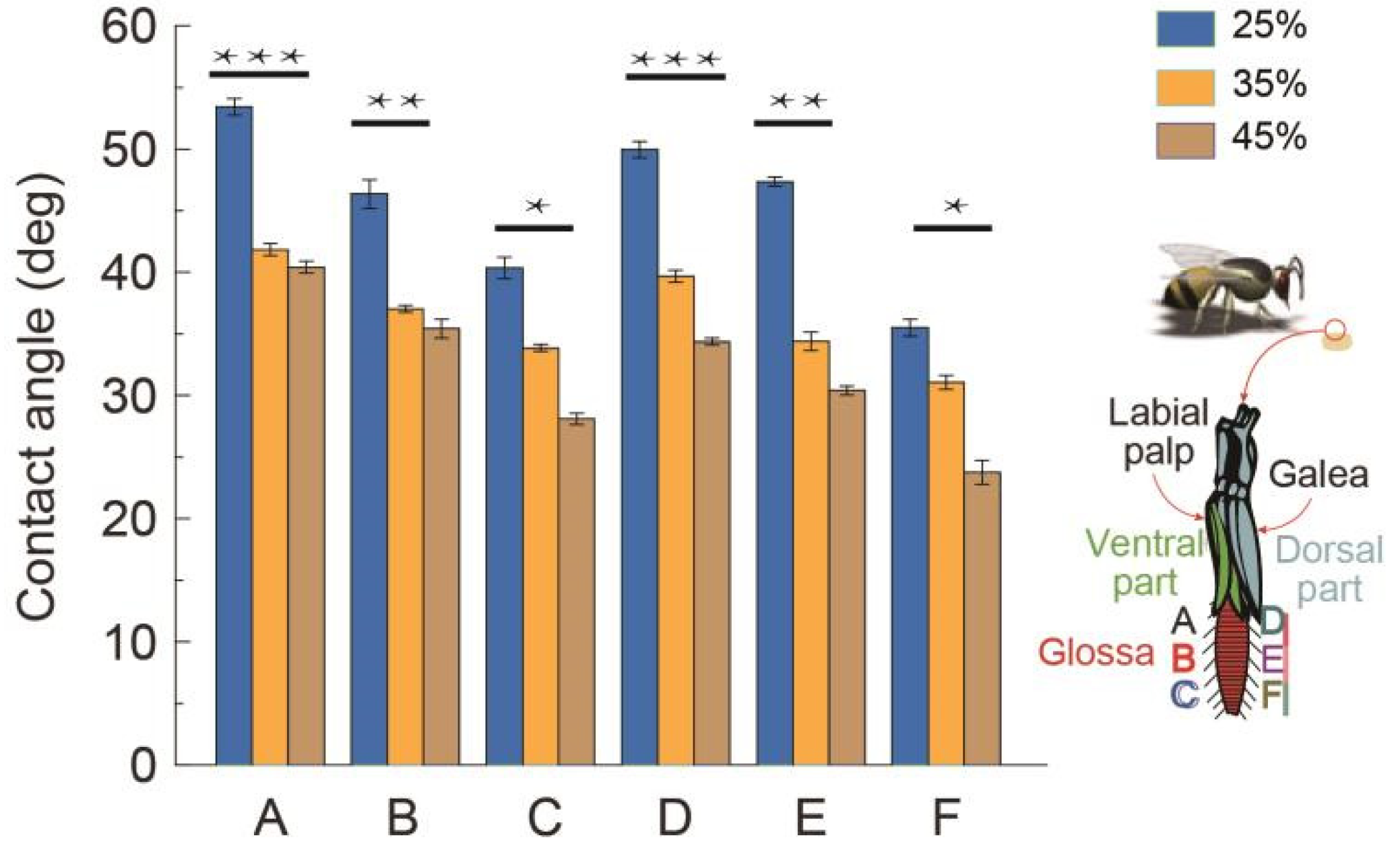
3.1. Section-Wise Wettability of the Glossa
3.2. Facultative Feeding Modes in Honey Bees
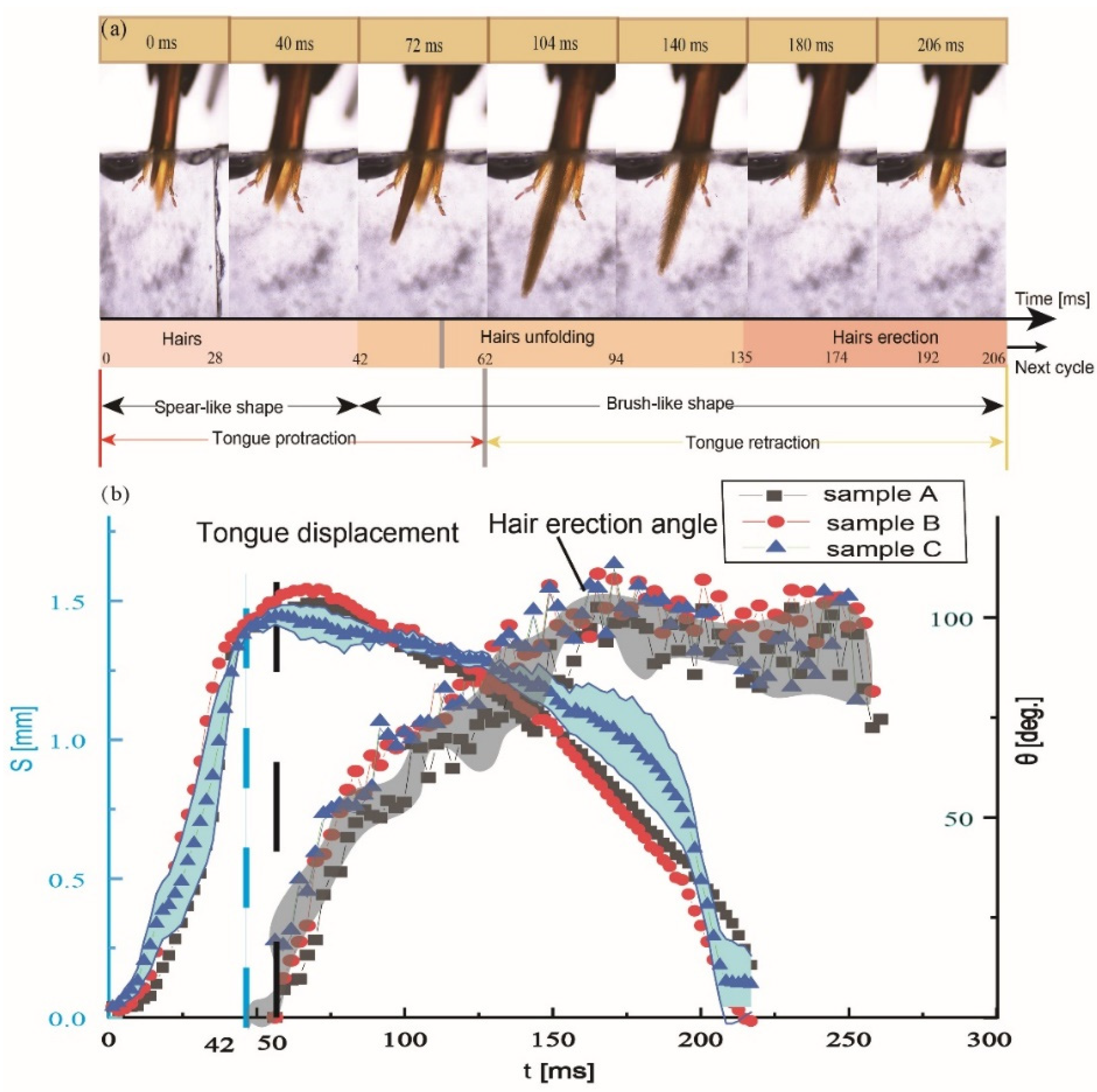
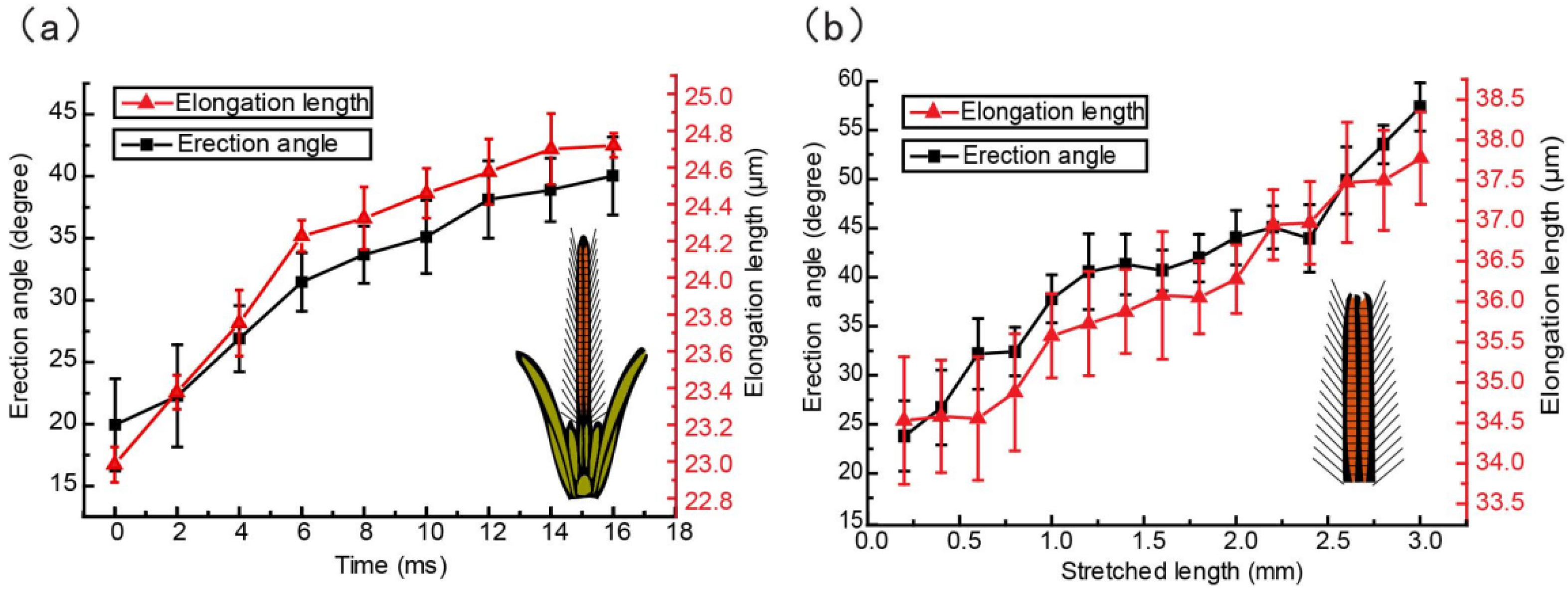
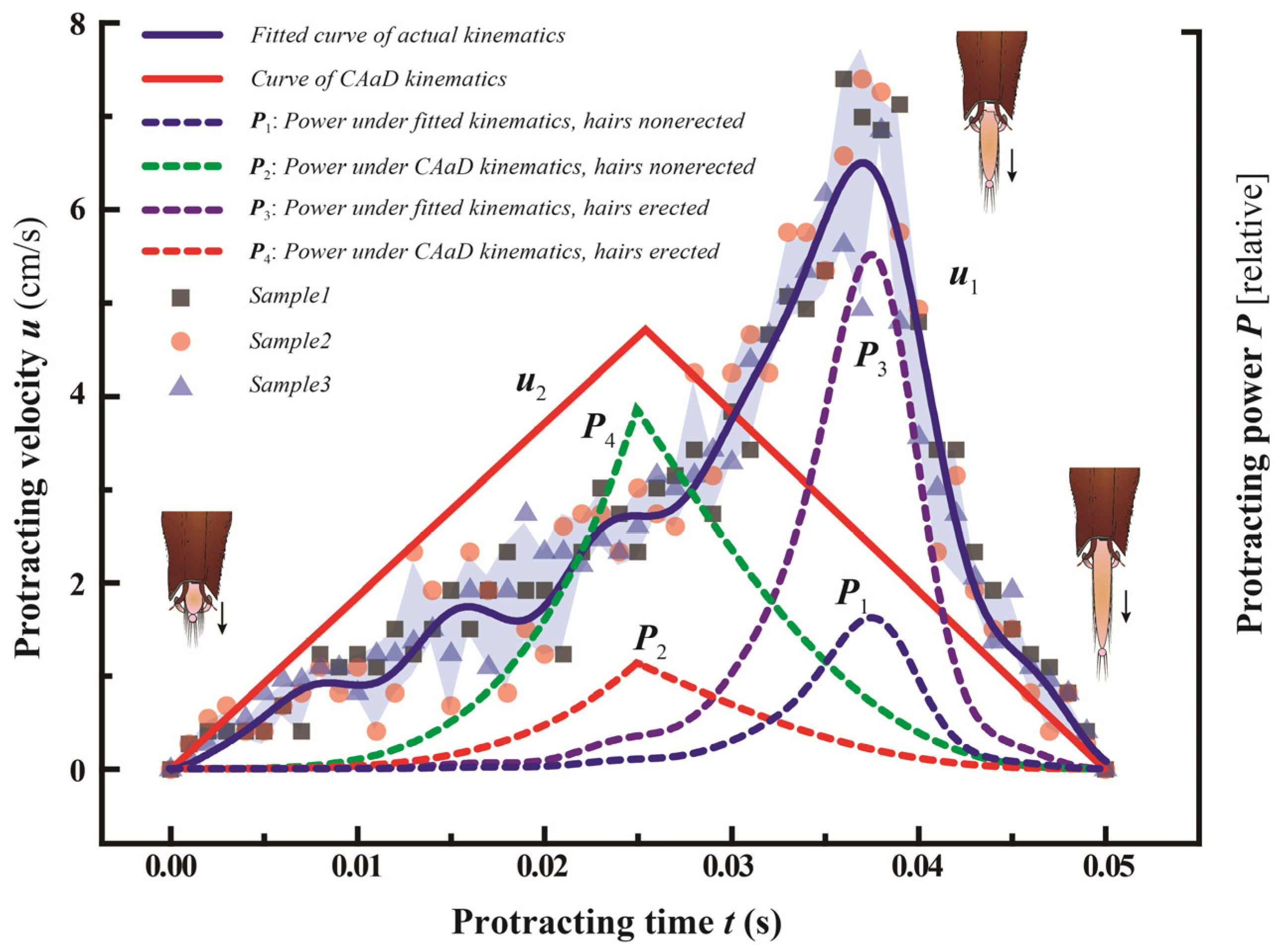
3.3. Kinematics of the Glossa and Glossal Hairs
3.4. Coordinated Movements of the Abdomen While Dipping Nectar
4. Energy Saving Strategy
4.1. Modelling for Energy Saving
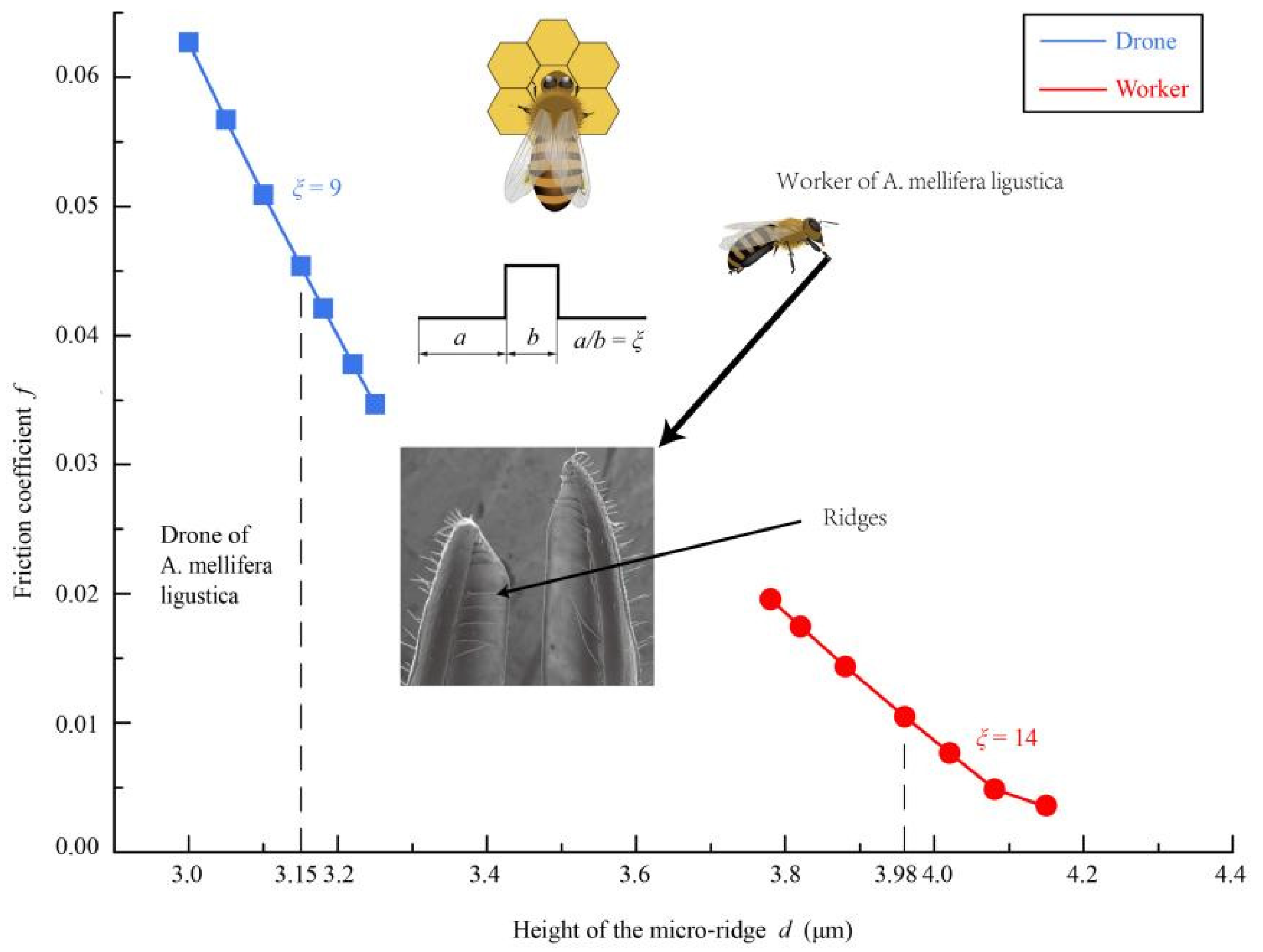
4.2. Effects of Galea Ridges on Drag Reduction
5. Functional Compensation by Regulating Dipping Frequency
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cook, J.M.; Rasplus, J.-Y. Mutualists with attitude: Coevolving fig wasps and figs. Trends Ecol. Evol. 2003, 18, 241–248. [Google Scholar] [CrossRef]
- Zhang, H.; Song, J.; Zhao, H.; Li, M.; Han, W. Predicting the Distribution of the Invasive Species Leptocybe invasa: Combining MaxEnt and Geodetector Models. Insects 2021, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Elberling, H.; Olesen, J.M. The structure of a high latitude plant-flower visitor system: The dominance of flies. Ecography 1999, 22, 314–323. [Google Scholar] [CrossRef]
- Ghazanfar, M.; Malik, M.F.; Hussain, M.; Iqbal, R.; Younas, M. Butterflies and their contribution in ecosystem: A review. J. Entomol. Zool. Stud. 2016, 4, 115–118. [Google Scholar]
- Cunningham, J.P.; Moore, C.J.; Zalucki, M.; West, S. Learning, odour preference and flower foraging in moths. J. Exp. Biol. 2004, 207, 87–94. [Google Scholar] [CrossRef]
- Krenn, H.; Plant, J.D.; Szucsich, N. Mouthparts of flower-visiting insects. Arthropod Struct. Dev. 2005, 34, 1–40. [Google Scholar] [CrossRef]
- Lechantre, A.; Michez, D.; Damman, P. Collection of nectar by bumblebees: How the physics of fluid demonstrates the prominent role of the tongue’s morphology. Soft Matter 2019, 15, 6392–6399. [Google Scholar] [CrossRef] [PubMed]
- Lechantre, A.; Draux, A.; Hua, H.-A.B.; Michez, D.; Damman, P.; Brau, F. Essential role of papillae flexibility in nectar capture by bees. Proc. Natl. Acad. Sci. USA 2021, 118, e2025513118. [Google Scholar] [CrossRef]
- Breeze, T.; Bailey, A.; Balcombe, K.; Potts, S. Pollination services in the UK: How important are honeybees? Agric. Ecosyst. Environ. 2011, 142, 137–143. [Google Scholar] [CrossRef]
- Wei, J.; Huo, Z.; Gorb, S.N.; Rico-Guevara, A.; Wu, Z.; Wu, J. Sucking or lapping: Facultative feeding mechanisms in honeybees (Apis mellifera). Biol. Lett. 2020, 16, 20200449. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Daniel, T.L. Mechanical determinants of nectar feeding strategy in hummingbirds: Energetics, tongue morphology, and licking behavior. Oecologia 1983, 60, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, R.E.; Morse, R.A. Anatomy of the Honey Bee; Cornell University Press: Ithaca, NY, USA, 1985. [Google Scholar]
- Nicolson, S.W.; Nepi, M.; Pacini, E. Nectaries and Nectar; Springer: Berlin/Heidelberg, Germany, 2007; Volume 4. [Google Scholar]
- Burrows, M.; Sutton, G. Interacting Gears Synchronize Propulsive Leg Movements in a Jumping Insect. Science 2013, 341, 1254–1256. [Google Scholar] [CrossRef]
- Zhao, J.; Meng, F.; Yan, S.; Wu, J.; Liang, Y.; Zhang, Y. Abdominal pumping involvement in the liquid feeding of honeybee. J. Insect Physiol. 2019, 112, 109–116. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, J.; Yan, S. Observations and temporal model of a honeybee’s hairy tongue in microfluid transport. J. Appl. Phys. 2015, 118, 194701. [Google Scholar] [CrossRef]
- Kumar, G.; Prabhu, K.N. Review of non-reactive and reactive wetting of liquids on surfaces. Adv. Colloid Interface Sci. 2007, 133, 61–89. [Google Scholar] [CrossRef]
- Lehnert, M.S.; Bennett, A.; Reiter, K.; Gerard, P.D.; Wei, Q.-H.; Byler, M.; Yan, H.; Lee, W.-K. Mouthpart conduit sizes of fluid-feeding insects determine the ability to feed from pores. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162026. [Google Scholar] [CrossRef] [PubMed]
- Rico-Guevara, A.; Rubega, M.A.; Hurme, K.J.; Dudley, R. Shifting Paradigms in the Mechanics of Nectar Extraction and Hummingbird Bill Morphology. Integr. Org. Biol. 2019, 1, oby006. [Google Scholar] [CrossRef] [PubMed]
- Pyke, G.; Waser, N.M. The Production of Dilute Nectars by Hummingbird and Honeyeater Flowers. Biotropica 1981, 13, 260. [Google Scholar] [CrossRef]
- Wu, J.; Zhu, R.; Yan, S.; Yang, Y. Erection pattern and section-wise wettability of honeybee glossal hairs in nectar feeding. J. Exp. Biol. 2015, 218, 664–667. [Google Scholar] [CrossRef][Green Version]
- Nicolson, S.W.; De Veer, L.; Köhler, A.; Pirk, C. Honeybees prefer warmer nectar and less viscous nectar, regardless of sugar concentration. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131597. [Google Scholar] [CrossRef]
- Seeley, T.D. Social foraging by honeybees: How colonies allocate foragers among patches of flowers. Behav. Ecol. Sociobiol. 1986, 19, 343–354. [Google Scholar] [CrossRef]
- Kakutani, T.; Inoue, T.; Kato, M. Nectar secretion pattern of the dish-shaped flower, Cayratia japonica (Vitaceae), and nectar utilization patterns by insect visitors. Popul. Ecol. 1989, 31, 381–400. [Google Scholar] [CrossRef]
- Hunt, G.J.; Page, R.E.; Fondrk, M.K.; Dullum, C.J.; Hunt, G.J.; Page, R.E., Jr.; Fondrk, M.K.; Dullum, C.J. Major quantitative trait loci affecting honey bee foraging behavior. Genetics 1996, 141, 1537–1545. [Google Scholar] [CrossRef]
- Wu, J.; Yang, H.; Yan, S. Energy saving strategies of honeybees in dipping nectar. Sci. Rep. 2015, 5, 15002. [Google Scholar] [CrossRef]
- Zhu, R.; Lv, H.; Liu, T.; Yang, Y.; Wu, J.; Yan, S. Feeding Kinematics and Nectar Intake of the Honey Bee Tongue. J. Insect Behav. 2016, 29, 325–339. [Google Scholar] [CrossRef]
- Finlayson, L.H.; Lowenstein, O. The structure and function of abdominal stretch receptors in insects. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1958, 148, 433–449. [Google Scholar] [CrossRef]
- Camp, A.L.; Roberts, T.J.; Brainerd, E.L. Swimming muscles power suction feeding in largemouth bass. Proc. Natl. Acad. Sci. USA 2015, 112, 8690–8695. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Bush, J.W.M. Natural drinking strategies. J. Fluid Mech. 2012, 705, 7–25. [Google Scholar] [CrossRef]
- Hou, Q.; Yang, X.; Cheng, J.; Wang, S.; Duan, D.; Xiao, J.; Li, W. Optimization of Performance Parameters and Mechanism of Bionic Texture on Friction Surface. Coatings 2020, 10, 171. [Google Scholar] [CrossRef]
- Li, C.; Wu, J.-N.; Yang, Y.-Q.; Zhu, R.-G.; Yan, S.-Z. Drag reduction effects facilitated by microridges inside the mouthparts of honeybee workers and drones. J. Theor. Biol. 2016, 389, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R. Die Sehfelder und Ommatidien-Divergenzwinkel der drei Kasten der Honigbiene (Apis mellifica). Verh. Zool. Gesell. 1980, 359–373. [Google Scholar]
- Hrassnigg, N.; Crailsheim, K. Differences in drone and worker physiology in honeybees (Apis mellifera). Apidologie 2005, 36, 255–277. [Google Scholar] [CrossRef]
- Tautz, J. The Buzz about Bees: Biology of a Superorganism; Springer Science & Business Media: Berlin, Germany, 2008. [Google Scholar]
- Schmickl, T.; Crailsheim, K. How honeybees (Apis mellifera L.) change their broodcare behaviour in response to non-foraging conditions and poor pollen conditions. Behav. Ecol. Sociobiol. 2002, 51, 415–425. [Google Scholar]
- Schmickl, T.; Crailsheim, K. Inner nest homeostasis in a changing environment with special emphasis on honey bee brood nursing and pollen supply. Apidologie 2004, 35, 249–263. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World; JHU Press: Baltimore, MD, USA, 2000; Volume 1. [Google Scholar]
- Wu, J.; Chen, Y.; Li, C.; Lehnert, M.S.; Yang, Y.; Yan, S. A quick tongue: Older honey bees dip nectar faster to compensate for mouthpart structure deterioration. J. Exp. Biol. 2019, 222, jeb212191. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Revue Sci. Tech. Office Int. Epizoot. 2008, 27, 499–510. [Google Scholar]
- Brown, M.J.F.; Paxton, R. The conservation of bees: A global perspective. Apidologie 2009, 40, 410–416. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wu, Z.; Zhao, J.; Wu, J. Nectar Feeding by a Honey Bee’s Hairy Tongue: Morphology, Dynamics, and Energy-Saving Strategies. Insects 2021, 12, 762. https://doi.org/10.3390/insects12090762
Wang H, Wu Z, Zhao J, Wu J. Nectar Feeding by a Honey Bee’s Hairy Tongue: Morphology, Dynamics, and Energy-Saving Strategies. Insects. 2021; 12(9):762. https://doi.org/10.3390/insects12090762
Chicago/Turabian StyleWang, Hao, Zhigang Wu, Jieliang Zhao, and Jianing Wu. 2021. "Nectar Feeding by a Honey Bee’s Hairy Tongue: Morphology, Dynamics, and Energy-Saving Strategies" Insects 12, no. 9: 762. https://doi.org/10.3390/insects12090762
APA StyleWang, H., Wu, Z., Zhao, J., & Wu, J. (2021). Nectar Feeding by a Honey Bee’s Hairy Tongue: Morphology, Dynamics, and Energy-Saving Strategies. Insects, 12(9), 762. https://doi.org/10.3390/insects12090762