
Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing

 , ,
, , 
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. S. frugiperda Sample Collection and Genomic DNA Extraction
2.2. TaqMan Genotyping for SfABCC2mut
2.3. Targeted DNA Amplification and Sequencing
2.4. Genotype Assignment for SfABCC2mut Using a Targeted-Sequencing k-mer-Based Approach
2.5. Alignment, Variant Calling and Variant Analysis
3. Results
3.1. Genotyping Results Using Taqman
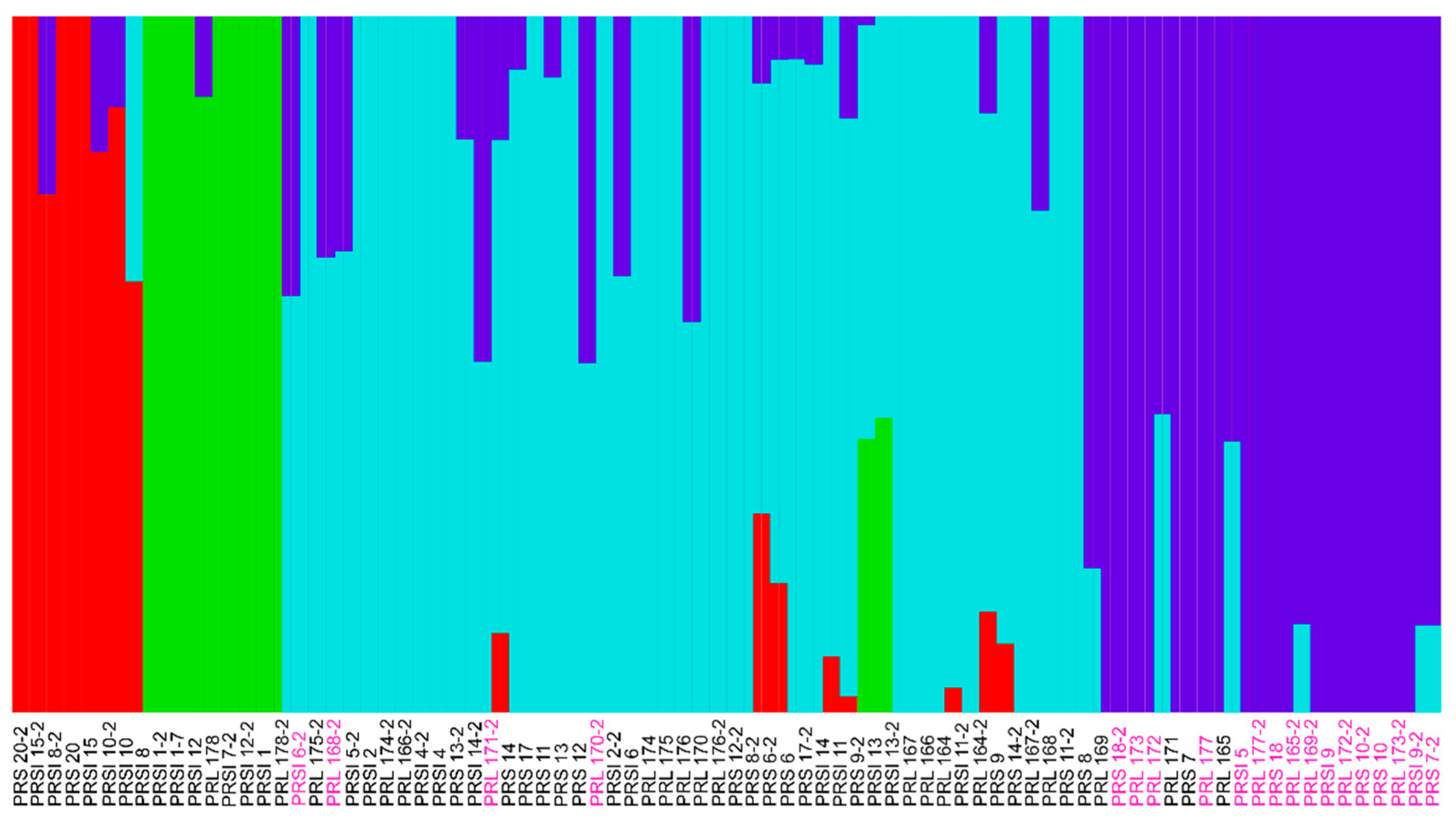
3.2. Genotyping Results Using Targeted Sequencing
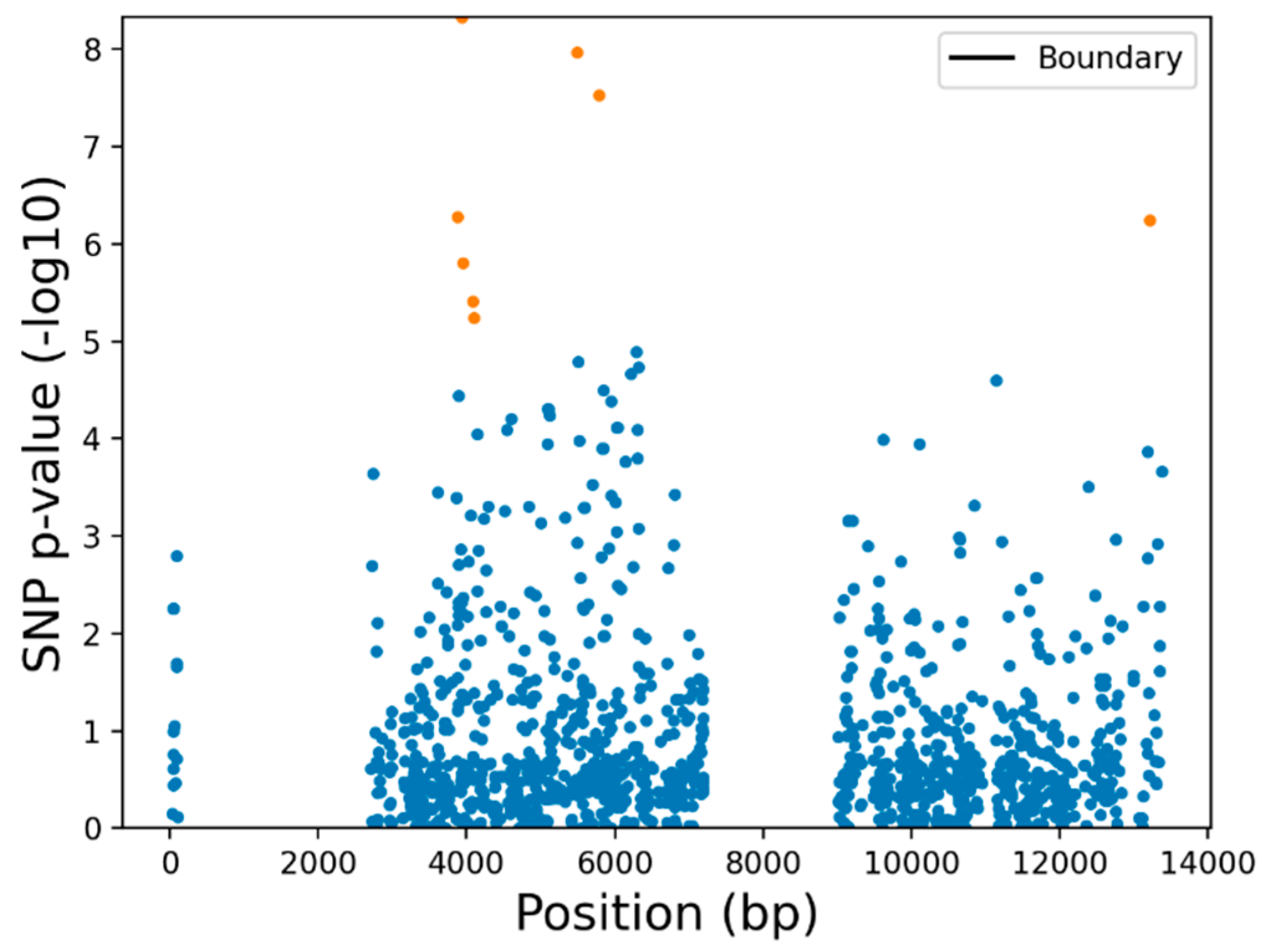
3.3. Variant Analysis Pipeline and Genotyping
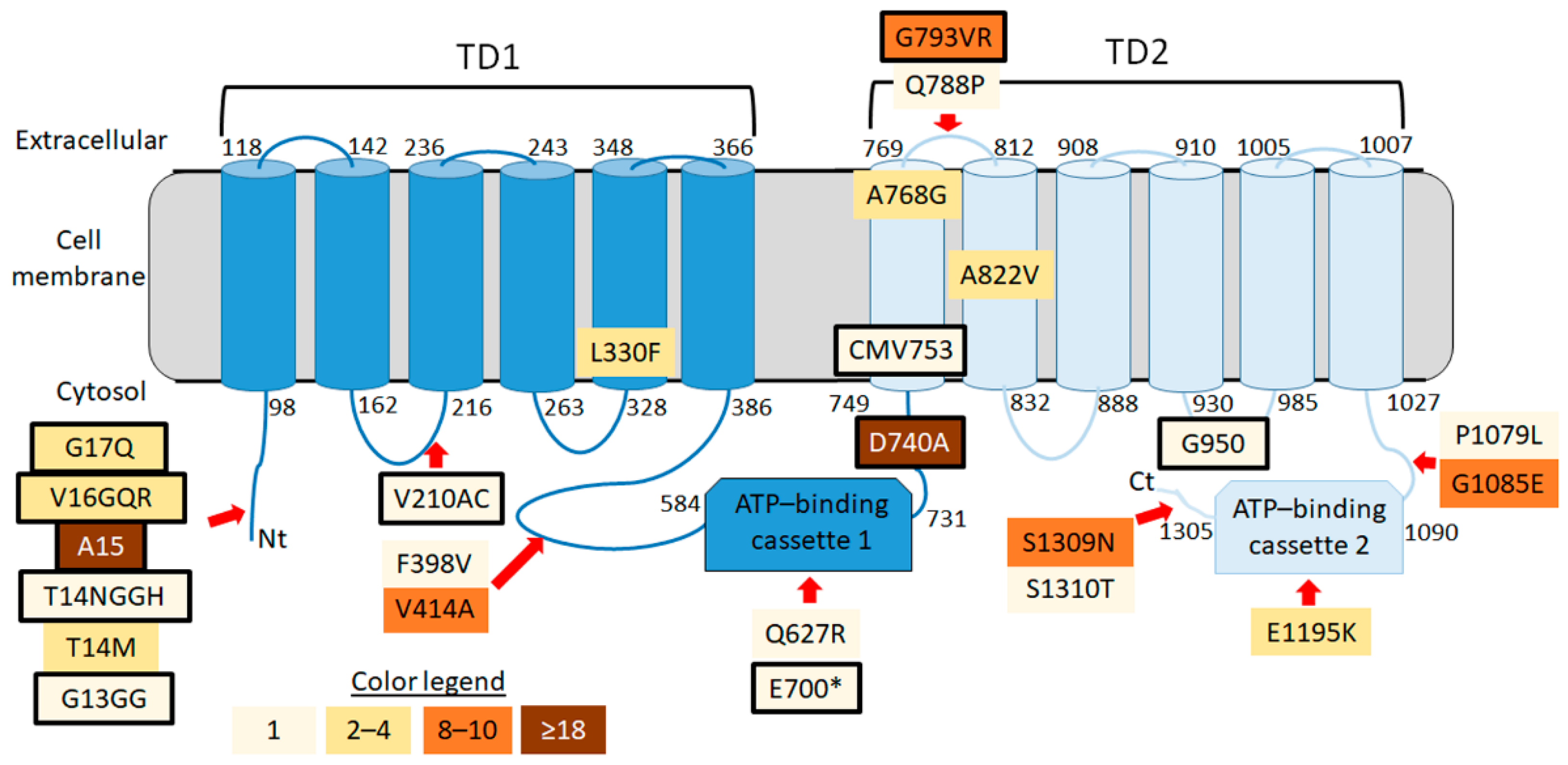
3.4. SfABCC2 Sequence Variation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.; Fleischer, S.; Jairam, S.; Meagher, R.; Nagoshi, R. Multigenerational migration of fall armyworm, a pest insect. Ecosphere 2019, 10, e02919. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Fleischer, S.; Meagher, R.L.; Hay-Roe, M.; Khan, A.; Murua, M.G.; Silvie, P.; Vergara, C.; Westbrook, J. Fall armyworm migration across the Lesser Antilles and the potential for genetic exchanges between North and South American populations. PLoS ONE 2017, 12, e0171743. [Google Scholar] [CrossRef] [Green Version]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Qureshi, J.A.; Meagher, R.L., Jr.; Reisig, D.D.; Head, G.P.; Andow, D.A.; Ni, X.; Kerns, D.; Buntin, G.D.; Niu, Y.; et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: Single gene versus pyramided Bt maize. PLoS ONE 2014, 9, e112958. [Google Scholar] [CrossRef] [Green Version]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Protect. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Chandrasena, D.I.; Signorini, A.M.; Abratti, G.; Storer, N.P.; Olaciregui, M.L.; Alves, A.P.; Pilcher, C.D. Characterization of field-evolved resistance to Bacillus thuringiensis-derived Cry1F delta-endotoxin in Spodoptera frugiperda populations from Argentina. Pest Manag. Sci. 2018, 74, 746–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; Hasler, J.; Meagher, R.; Nagoshi, R.; Hietala, L.; Huang, F.; Narva, K.; Jurat-Fuentes, J.L. Mechanism and DNA-based detection of field-evolved resistance to transgenic Bt corn in fall armyworm (Spodoptera frugiperda). Sci. Rep. 2017, 7, 10877. [Google Scholar] [CrossRef] [Green Version]
- Flagel, L.; Lee, Y.W.; Wanjugi, H.; Swarup, S.; Brown, A.; Wang, J.; Kraft, E.; Greenplate, J.; Simmons, J.; Adams, N.; et al. Mutational disruption of the ABCC2 gene in fall armyworm, Spodoptera frugiperda, confers resistance to the Cry1Fa and Cry1A.105 insecticidal proteins. Sci. Rep. 2018, 8, 7255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boaventura, D.; Ulrich, J.; Lueke, B.; Bolzan, A.; Okuma, D.; Gutbrod, O.; Geibel, S.; Zeng, Q.; Dourado, P.M.; Martinelli, S.; et al. Molecular characterization of Cry1F resistance in fall armyworm, Spodoptera frugiperda from Brazil. Insect Biochem. Mol. Biol. 2020, 116, 103280. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, M.; Wang, L.; Wang, H.; Xia, Z.; Yang, Y.; Bravo, A.; Soberón, M.; Xiao, Y.; Liu, K. SfABCC2 transporter extracellular loops 2 and 4 are responsible for the Cry1Fa insecticidal specificity against Spodoptera frugiperda. Insect Biochem. Mol. Biol. 2021, 135, 103608. [Google Scholar] [CrossRef]
- Jin, M.-H.; Tao, J.-H.; Li, Q.; Cheng, Y.; Sun, X.-X.; Wu, K.-M.; Xiao, Y.-T. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2020, in press. [Google Scholar] [CrossRef]
- Abdelgaffar, H.; Perera, O.P.; Jurat-Fuentes, J.L. ABC transporter mutations in Cry1F-resistant fall armyworm (Spodoptera frugiperda) do not result in altered susceptibility to selected small molecule pesticides. Pest Manag. Sci. 2021, 77, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Vélez, A.M.; Spencer, T.A.; Alves, A.P.; Moellenbeck, D.; Meagher, R.L.; Chirakkal, H.; Siegfried, B.D. Inheritance of Cry1F resistance, cross-resistance and frequency of resistant alleles in Spodoptera frugiperda (Lepidoptera: Noctuidae). Bull. Entomol. Res. 2013, 103, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakka, S.R.; Knight, V.R.; Jurat-Fuentes, J.L. Fitness costs associated with field-evolved resistance to Bt maize in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Dumont, T.; Teo, Z.L.; Pope, B.J.; Hammet, F.; Mahmoodi, M.; Tsimiklis, H.; Sabbaghian, N.; Tischkowitz, M.; Foulkes, W.D.; Giles, G.G.; et al. Hi-Plex for high-throughput mutation screening: Application to the breast cancer susceptibility gene PALB2. BMC Med. Genom. 2013, 6, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen-Dumont, T.; Pope, B.J.; Hammet, F.; Southey, M.C.; Park, D.J. A high-plex PCR approach for massively parallel sequencing. BioTechniques 2013, 55, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Storer, N.P.; Kubiszak, M.E.; Ed King, J.; Thompson, G.D.; Santos, A.C. Status of resistance to Bt maize in Spodoptera frugiperda: Lessons from Puerto Rico. J. Invertebr. Pathol. 2012, 110, 294–300. [Google Scholar] [CrossRef]
- Meagher, R.L. Trapping fall armyworm (Lepidoptera: Noctuidae) adults in traps baited with pheromone and a synthetic floral volatile compound. Fla. Entomol. 2001, 84, 288–292. [Google Scholar] [CrossRef]
- Capinera, J.L.; University of Florida. Fall Armyworm. Publication Number EENY-98. 2017. Available online: http://entnemdept.ufl.edu/creatures/field/fall_armyworm.htm#host (accessed on 20 May 2021).
- Blanco, C.A.; Portilla, M.; Jurat-Fuentes, J.L.; Sanchez, J.F.; Viteri, D.; Vega-Aquino, P.; Teran-Vargas, A.P.; Azuara-Dominguez, A.; Lopez, J.D.J.; Arias, R.; et al. Susceptibility of isofamilies of Spodoptera frugiperda (Lepidoptera: Noctuidae) to Cry1Ac and Cry1Fa proteins of Bacillus thuringiensis. Southwest. Entomol. 2010, 35, 409–415. [Google Scholar] [CrossRef]
- Gervais, A.L.; Marques, M.; Gaudreau, L. PCRTiler: Automated design of tiled and specific PCR primer pairs. Nucleic Acids Res. 2010, 38, W308–W312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushnell, B. BBMap (BBMap Short Read Aligner, and Other Bioinformatic Tools). Available online: https://jgi.doe.gov/data-and-tools/bbtools/ (accessed on 20 June 2019).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequencing Data [Online]. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 December 2017).
- Gimenez, S.; Abdelgaffar, H.; Goff, G.L.; Hilliou, F.; Blanco, C.A.; Hänniger, S.; Bretaudeau, A.; Legeai, F.; Nègre, N.; Jurat-Fuentes, J.L.; et al. Adaptation by copy number variation increases insecticide resistance in the fall armyworm. Commun. Biol. 2020, 3, 664. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Slater, G.S.; Birney, E. Automated generation of heuristics for biological sequence comparison. BMC Bioinform. 2005, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 370–3071. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (Austin) 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Delaneau, O.; Marchini, J.; Zagury, J.F. A linear complexity phasing method for thousands of genomes. Nat. Methods 2011, 9, 179–181. [Google Scholar] [CrossRef]
- Cheng, T.; Wu, J.; Wu, Y.; Chilukuri, R.V.; Huang, L.; Yamamoto, K.; Feng, L.; Li, W.; Chen, Z.; Guo, H.; et al. Genomic adaptation to polyphagy and insecticides in a major East Asian noctuid pest. Nat. Ecol. Evol. 2017, 1, 1747–1756. [Google Scholar] [CrossRef]
- Raj, A.; Stephens, M.; Pritchard, J.K. fastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Qureshi, J.A.; Ni, X.; Head, G.P.; Price, P.A.; Meagher, R.L.; Kerns, D.; Levy, R.; Yang, X.; Huang, F. F2 screen for resistance to Bacillus thuringiensis Cry2Ab2-maize in field populations of Spodoptera frugiperda (Lepidoptera: Noctuidae) from the southern United States. J. Invertebr. Pathol. 2016, 138, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Bernardi, D.; Ribeiro, R.D.S.; do Nascimento, A.R.; Dos Santos, A.C.; Omoto, C. Frequency of Cry1F resistance alleles in Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Pest Manag. Sci. 2016, 72, 2295–2302. [Google Scholar] [CrossRef] [PubMed]
- Yainna, S.; Nègre, N.; Silvie, P.J.; Brévault, T.; Tay, W.T.; Gordon, K.; dAlençon, E.; Walsh, T.; Nam, K. Geographic monitoring of insecticide resistance mutations in native and invasive populations of the fall armyworm. Insects 2021, 12, 468. [Google Scholar] [CrossRef] [PubMed]
- Viteri, D.M.; Linares, A.M.; Cabrera, I.; Sarmiento, L. Presence of corn earworm and fall armyworm (Lepidoptera: Noctuidae) populations in sweet corn and their susceptibility to insecticides in Puerto Rico. Fla. Entomol. 2019, 102, 451–454. [Google Scholar] [CrossRef] [Green Version]
- Schlum, K.A.; Lamour, K.; de Bortoli, C.P.; Banerjee, R.; Meagher, R.; Pereira, E.; Murua, M.G.; Sword, G.A.; Tessnow, A.E.; Viteri Dillon, D.; et al. Whole genome comparisons reveal panmixia among fall armyworm (Spodoptera frugiperda) from diverse locations. BMC Genom. 2021, 22, 179. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Chandrasena, D.; Difonzo, C.; Conner, J.; Head, G.; Berman, K.; Wise, J. Susceptibility of fall armyworms (Spodoptera frugiperda j.e.) from mexico and puerto rico to Bt proteins. Insects 2020, 11, 831. [Google Scholar] [CrossRef] [PubMed]
- Vélez, A.M.; Spencer, T.A.; Alves, A.P.; Crespo, A.L.B.; Siegfried, B.D. Fitness costs of Cry1F resistance in fall armyworm, Spodoptera frugiperda. J. Appl. Entomol. 2013, 138, 315–325. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Taqman | K-mer |
|---|---|---|
| PRL_164 | SS | SS |
| PRL_165 | rr | Sr |
| PRL_166 | SS | SS |
| PRL_167 | SS | SS |
| PRL_168 | Sr | Sr |
| PRL_169 | rr | Sr |
| PRL_170 | Sr | Sr |
| PRL_171 | Sr | Sr |
| PRL_172 | rr | rr |
| PRL_173 | rr | rr |
| PRL_174 | SS | SS |
| PRL_175 | SS | SS |
| PRL_176 | SS | SS |
| PRL_177 | rr | rr |
| PRL_178 | SS | SS |
| PRS_6 | SS | SS |
| PRS_7 | Sr | Sr |
| PRS_8 | SS | SS |
| PRS_9 | Sr | SS |
| PRS_10 | rr | rr |
| PRS_11 | SS | SS |
| PRS_12 | Sr | SS |
| PRS_13 | SS | SS |
| PRS_14 | SS | SS |
| PRS_17 | Sr | SS |
| PRS_18 | SS | rr |
| PRS_20 | SS | SS |
| PRSI_1 | SS | SS |
| PRSI_2 | SS | SS |
| PRSI_4 | SS | SS |
| PRSI_5 | SS | Sr |
| PRSI_6 | rr | Sr |
| PRSI_7 | rr | SS |
| PRSI_8 | SS | SS |
| PRSI_9 | SS | rr |
| PRSI_10 | rr | SS |
| PRSI_11 | SS | SS |
| PRSI_12 | SS | SS |
| PRSI_13 | SS | SS |
| PRSI_14 | SS | SS |
| PRSI_15 | SS | SS |
| Position | Class | Mutation | Aa Change | #Samples |
|---|---|---|---|---|
| 38 | SHIFT | ggc/ggTGGTc | G13GG | 1 |
| 40 | SHIFT | acg/aATGGTGGTCATcg | T14NGGH * | 1 |
| 41 | MISS | aCg/aTg | T14M | 4 |
| 42 | SHIFT | gct/ | A15 * | 20 |
| 45 | SHIFT | gtg/gGTCAACG | V16GQR * | 4 |
| 48 | SHIFT | ggc/CAggc | G17Q * | 4 |
| 3421 | SHIFT | gtc/gCATGtc | V210AC * | 1 |
| 4077 | MISS | Ctc/Ttc | L330F | 2 |
| 4357 | MISS | Ttt/Gtt | F398V | 1 |
| 4540 | MISS | gTg/gCg | V414A | 9 |
| 6020 | MISS | cAa/cGa | Q627R | 1 |
| 6915 | NON | Gaa/Taa | E700 * | 1 |
| 7034 | SHIFT | gat/gCGat | D740A * | 18 |
| 7070 | SHIFT | tgcatggtg/ | CMV753 | 1 |
| 7120 | MISS | gCt/gGt | A768G | 3 |
| 9098 | MISS | cAa/cCa | Q788P | 1 |
| 9122 | SHIFT | gga/GTAAgga | G793VR | 8 |
| 9335 | MISS | gCc/gTc | A822V | 3 |
| 10491 | SHIFT | gga/ | G950 | 1 |
| 11708 | MISS | cCg/cTg | P1079L | 1 |
| 11726 | MISS | gGa/gAa | G1085E | 10 |
| 12507 | MISS | Gaa/Aaa | E1195K | 2 |
| 12748 | MISS | aGt/aAt | S1209N | 10 |
| 13285 | MISS | Tcc/Acc | S1310T | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schlum, K.; Lamour, K.; Tandy, P.; Emrich, S.J.; de Bortoli, C.P.; Rao, T.; Viteri Dillon, D.M.; Linares-Ramirez, A.M.; Jurat-Fuentes, J.L. Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing. Insects 2021, 12, 618. https://doi.org/10.3390/insects12070618
Schlum K, Lamour K, Tandy P, Emrich SJ, de Bortoli CP, Rao T, Viteri Dillon DM, Linares-Ramirez AM, Jurat-Fuentes JL. Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing. Insects. 2021; 12(7):618. https://doi.org/10.3390/insects12070618
Chicago/Turabian StyleSchlum, Katrina, Kurt Lamour, Peter Tandy, Scott J. Emrich, Caroline Placidi de Bortoli, Tejas Rao, Diego M. Viteri Dillon, Angela M. Linares-Ramirez, and Juan Luis Jurat-Fuentes. 2021. "Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing" Insects 12, no. 7: 618. https://doi.org/10.3390/insects12070618
APA StyleSchlum, K., Lamour, K., Tandy, P., Emrich, S. J., de Bortoli, C. P., Rao, T., Viteri Dillon, D. M., Linares-Ramirez, A. M., & Jurat-Fuentes, J. L. (2021). Genetic Screening to Identify Candidate Resistance Alleles to Cry1F Corn in Fall Armyworm Using Targeted Sequencing. Insects, 12(7), 618. https://doi.org/10.3390/insects12070618