
Diversity, Host Plants and Potential Distribution of Edible Saturniid Caterpillars in Kenya

 , ,
, ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection and Preparation
2.3. Morphological Identification
2.4. Molecular Identification
2.4.1. Tissue Preparation, DNA Extraction and Quantification
2.4.2. PCR for Insect Samples
2.4.3. PCR for Plant Samples
2.4.4. Agarose Gel Electrophoresis, PCR Product Purification and Sequencing
2.4.5. Sequence Analysis
2.5. Distribution Modelling
2.5.1. Occurrence Data
2.5.2. Environmental Variables
2.5.3. Model Calibration and Accuracy Assessment
3. Results
3.1. Morphological Identification of Edible Saturniids
3.1.1. Bunaea alcinoe
3.1.2. Cirina forda
3.1.3. Gonimbrasia cocaulti
3.1.4. Gonimbrasia krucki
3.1.5. Gonimbrasia belina
3.1.6. Gonimbrasia zambesina
3.1.7. Gynanisa nigra
3.2. Molecular Identification
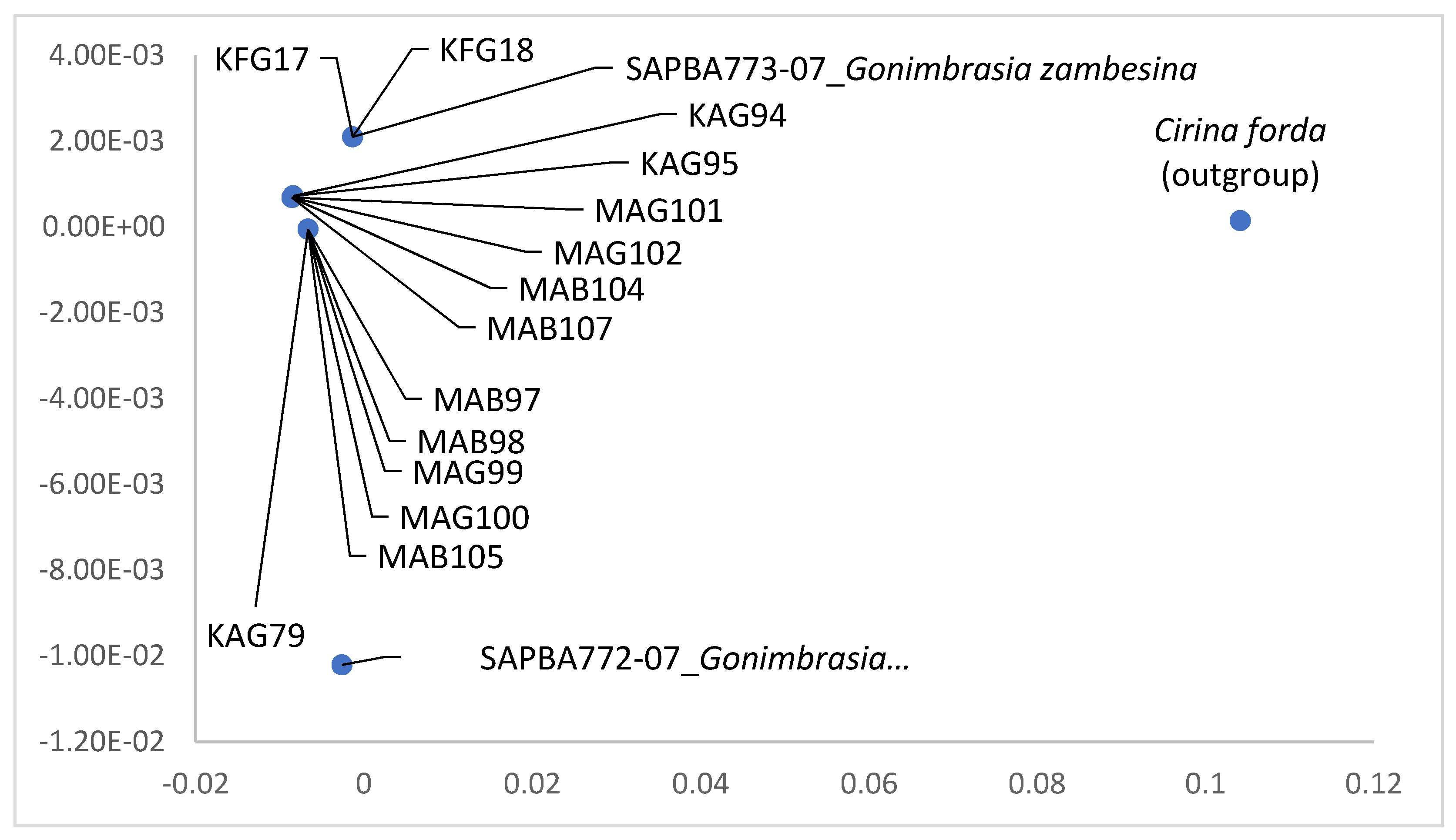
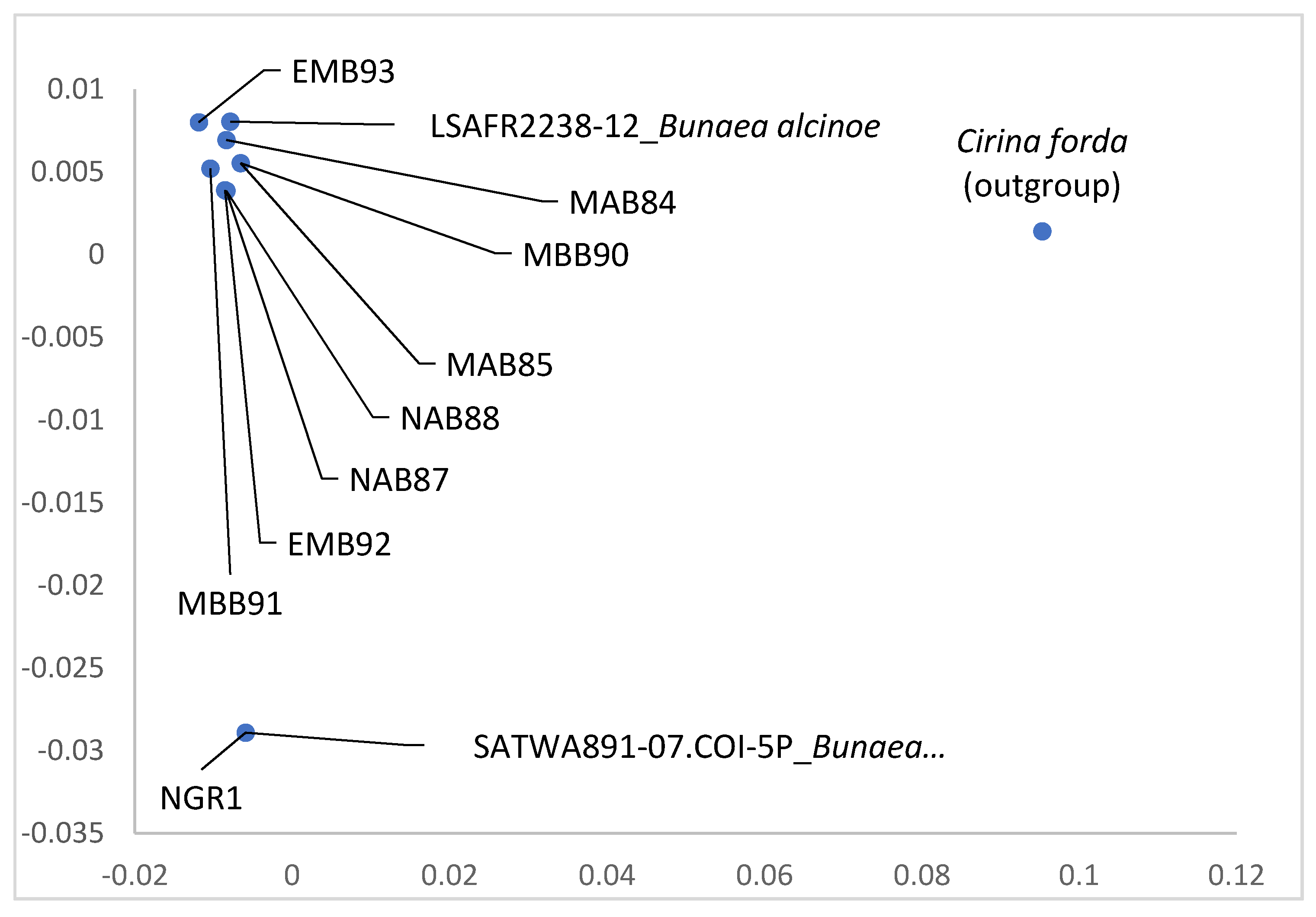
Molecular Differences among the Color Forms of Gonimbrasia zambesina and Bunaea alcinoe
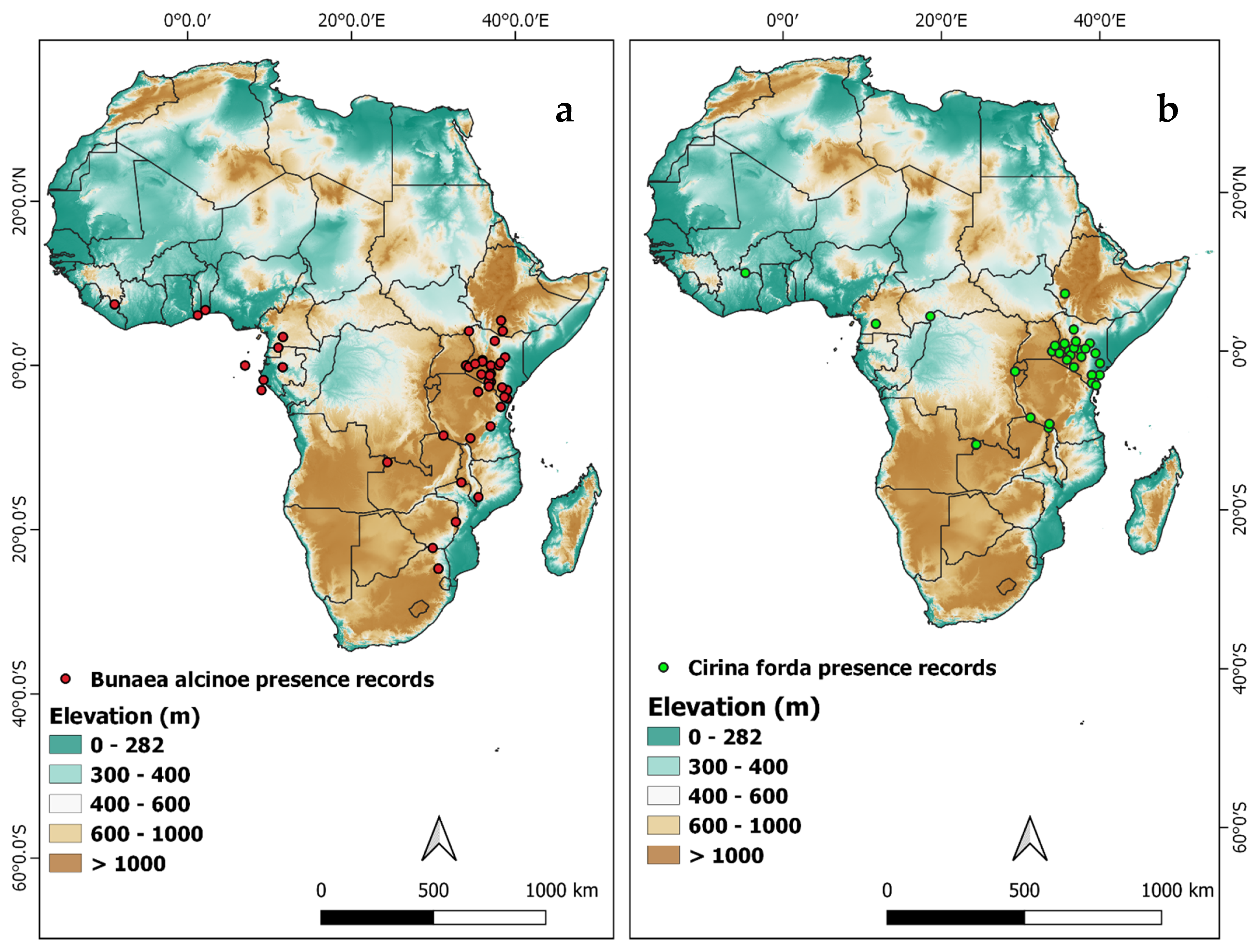
3.3. Distribution and Seasonality of Edible Saturniids in Kenya
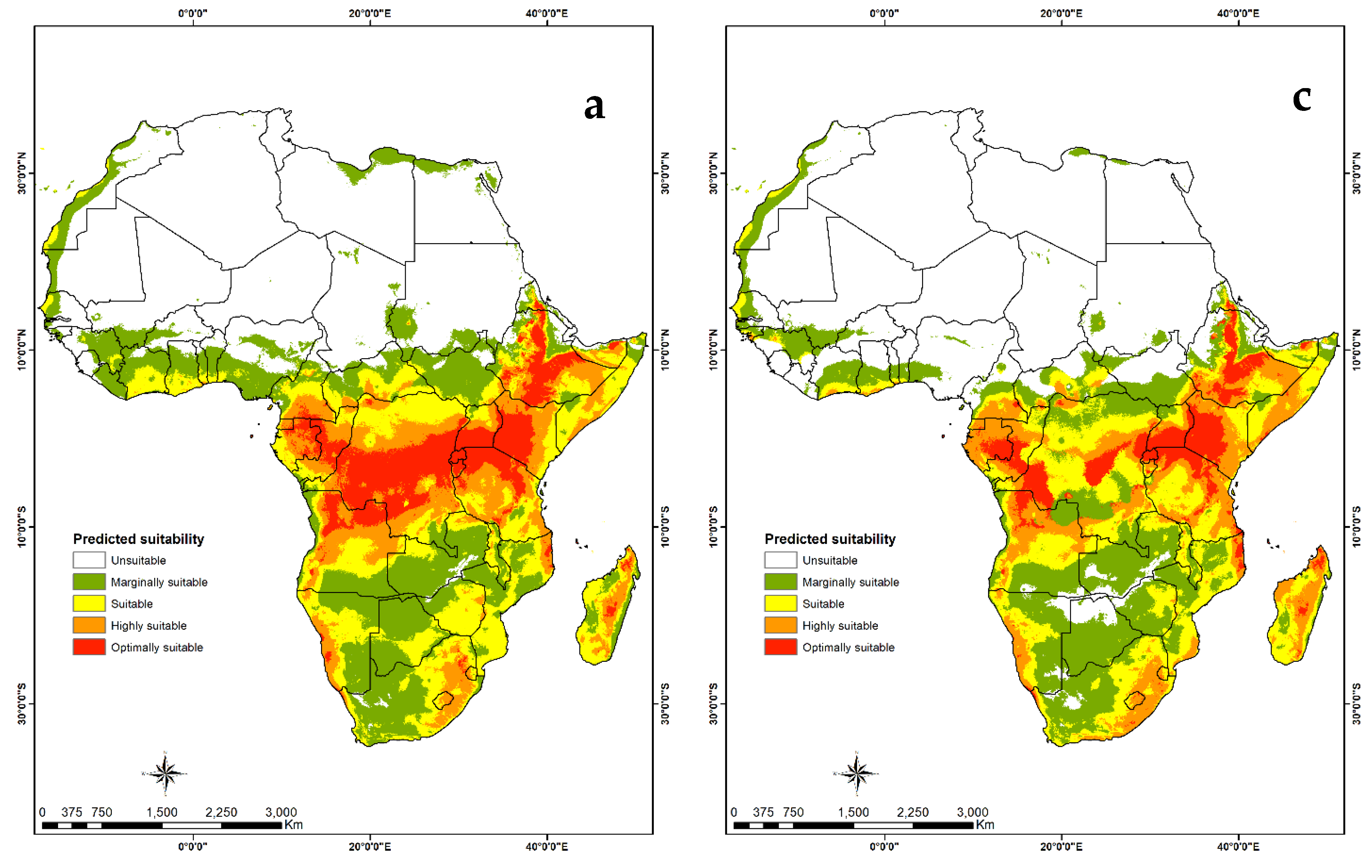
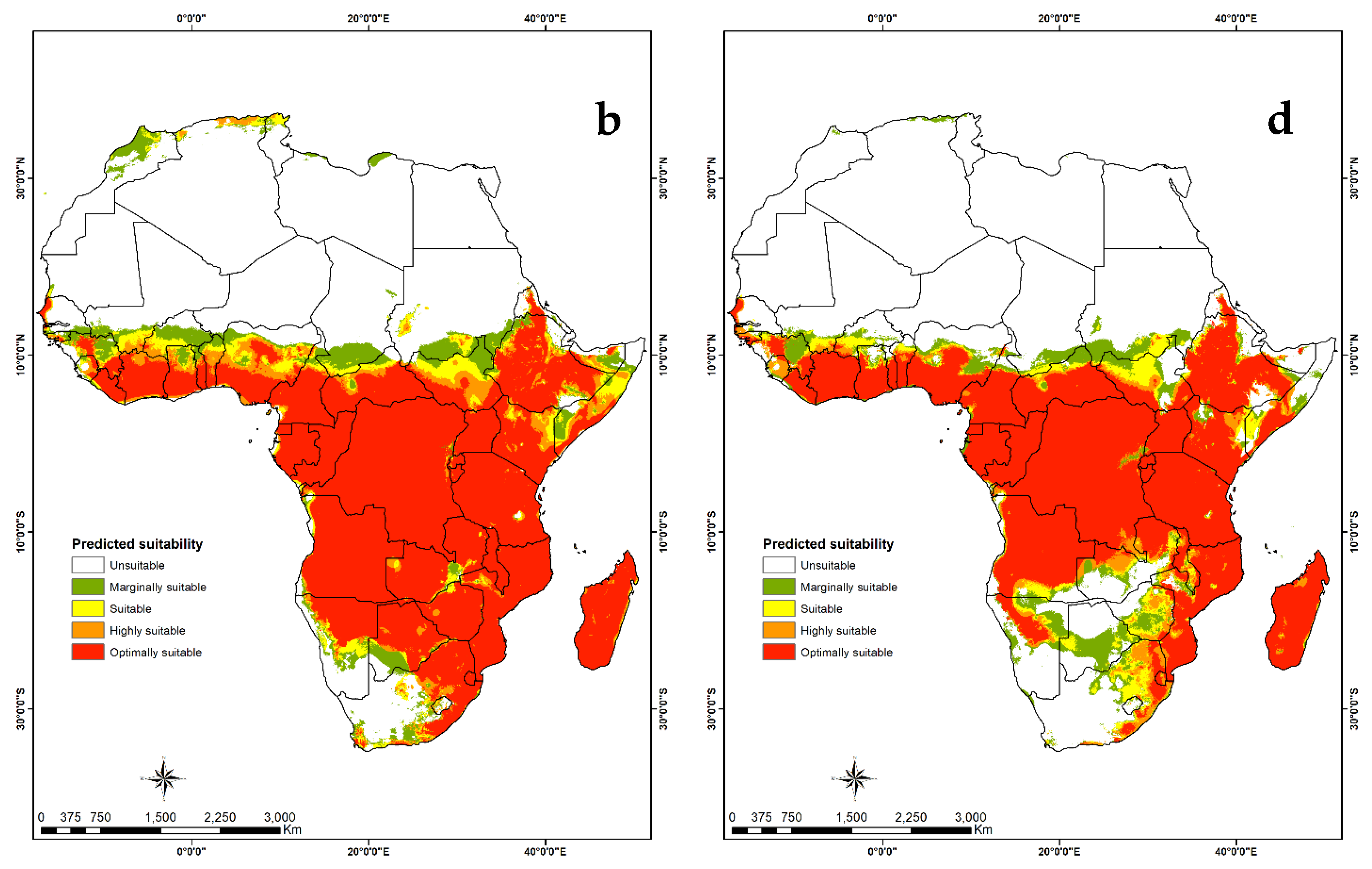
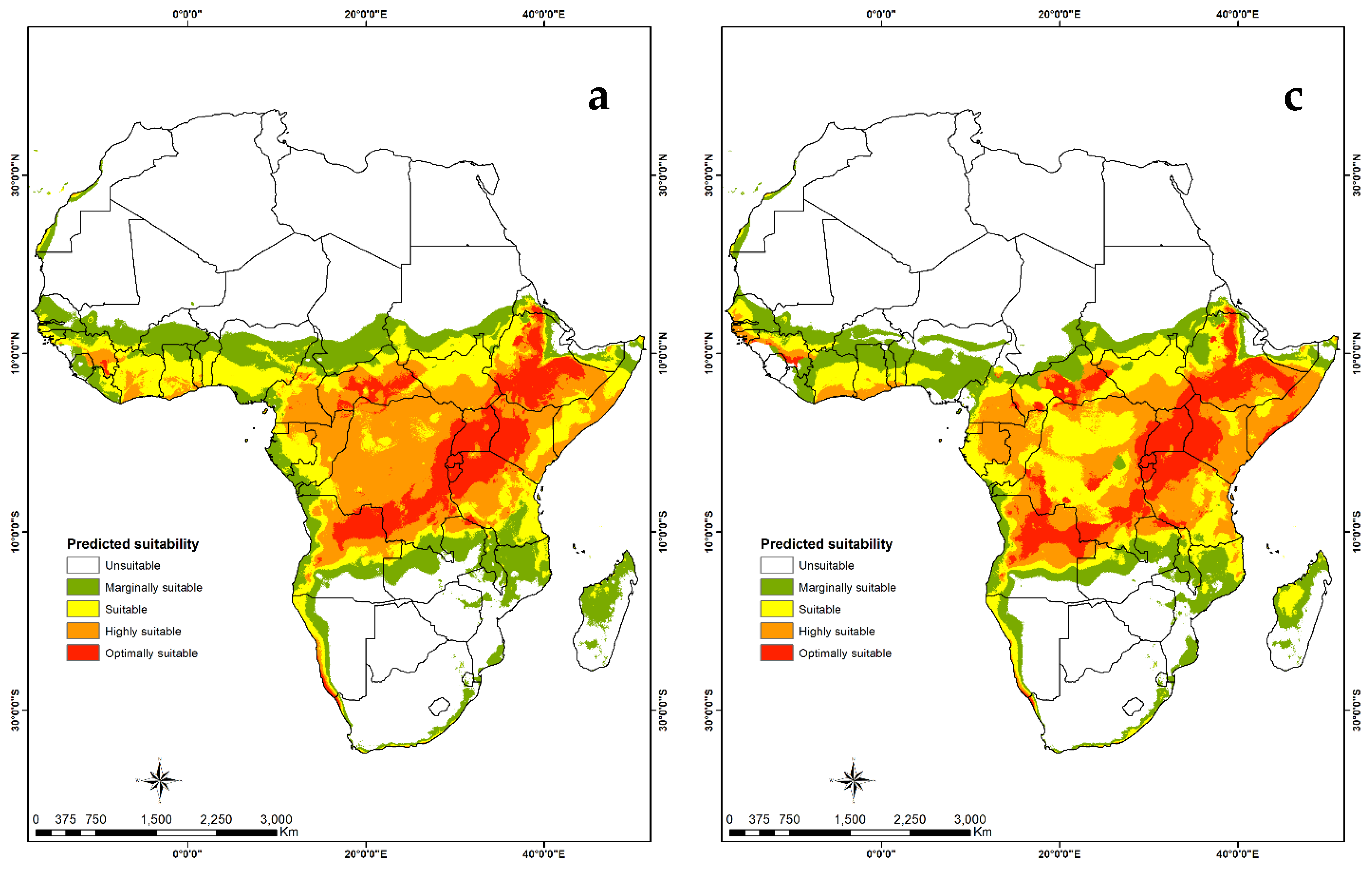
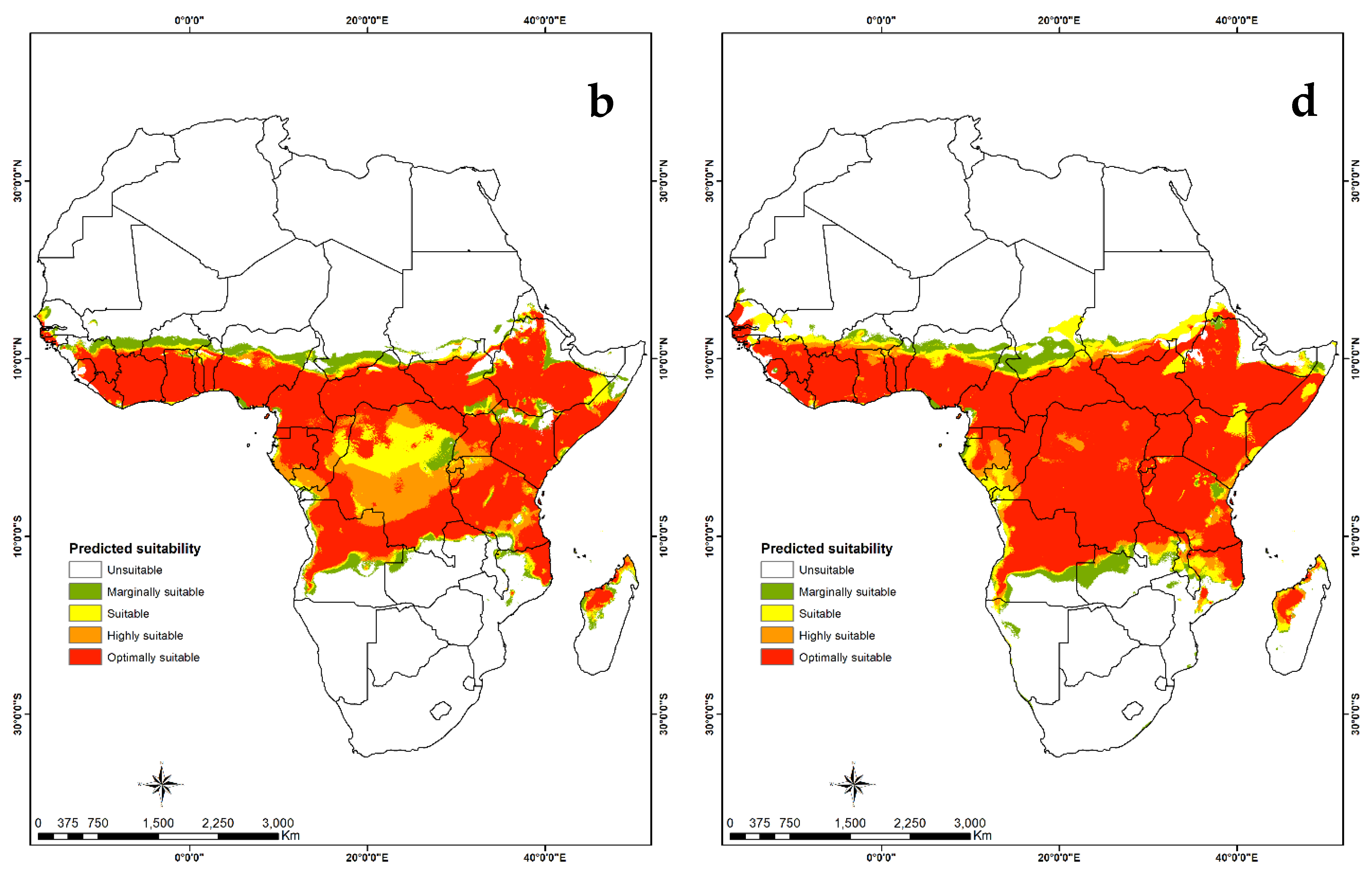
3.4. Habitat Suitability and Probability Distribution
3.4.1. Area under Curve (AUC) Values
3.4.2. Visualization of Habitat Suitability under Current and Future Climatic Conditions
3.5. Host Plants of Edible Saturniids in Kenya
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects. Future Prospects for Food and Feed Security; FAO Forestry Paper 171; FAO: Rome, Italy, 2013. [Google Scholar]
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Di Mattia, C.; Battista, N.; Sacchetti, G.; Serafini, M. Antioxidant activities in vitro of water and liposoluble extracts obtained by different species of edible insects and invertebrates. Front. Nutr. 2019, 6, 106. [Google Scholar] [CrossRef] [PubMed]
- Stull, V.J.; Finer, E.; Bergmans, R.S.; Febvre, H.P.; Longhurst, C.; Manter, D.K.; Patz, J.A.; Weir, T.L. Impact of edible cricket consumption on gut microbiota in healthy adults, a double-blind, randomized crossover trial. Sci. Rep. 2018, 8, 10762. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; de Boer, I.J.M. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Itterbeeck, J.; Heetkamp, M.J.W.; van den Brand, H.; van Loon, J.J.A.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed]
- Miglietta, P.; De Leo, F.; Ruberti, M.; Massari, S. Mealworms for food: A water footprint perspective. Water 2015, 7, 6190–6203. [Google Scholar] [CrossRef]
- Dobermann, D.; Michaelson, L.; Field, L.M. The effect of an initial high-quality feeding regime on the survival of Gryllus bimaculatus (black cricket) on bio-waste. J. Insects Food Feed 2019, 5, 117–123. [Google Scholar] [CrossRef]
- Magara, H.J.O.; Tanga, C.M.; Ayieko, M.A.; Hugel, S.; Mohamed, S.A.; Khamis, F.M.; Salifu, D.; Niassy, S.; Sevgan, S.; Fiaboe, K.K.M.; et al. Performance of newly described native edible cricket Scapsipedus icipe (Orthoptera: Gryllidae) on various diets of relevance for farming. J. Econ. Entomol. 2019, 112, 653–664. [Google Scholar] [CrossRef]
- Jongema, Y. List of Edible Insects of the World; Wageningen University: Wageningen, The Netherlands, 2017. [Google Scholar]
- Kelemu, S.; Niassy, S.; Torto, B.; Fiaboe, K.; Affognon, H.; Tonnang, H.; Maniania, N.K.; Ekesi, S. African edible insects for food and feed: Inventory, diversity, commonalities and contribution to food security. J. Insects Food Feed 2015, 1, 103–119. [Google Scholar] [CrossRef]
- Pinhey, E.C.G. The Emperor Moths of Eastern Africa; The East Africa Natural History Society: Nairobi, Kenya, 1956; Volume 23. [Google Scholar]
- Pinhey, E.C.G. Emperor Moths of South and South Central Africa; C. Struik: Cape Town, South Africa, 1972. [Google Scholar]
- Pinhey, E.C.G. Moths of Southern Africa: Descriptions and Colour Illustrations of 1183 Species, 1st ed.; Tafelberg Publishers Ltd.: Cape Town, South Africa, 1975. [Google Scholar]
- Ande, A.T. The influence of local harvesting and processing methods on some nutrient content of Cirina forda Westwood (Lepidoptera: Saturniidae) in Nigeria. Niger. J. Pure Appl. Sci. 2002, 17, 1165–1168. [Google Scholar]
- Badanaro, F.; Amevoin, K.; Lamboni, C.; Kou’santa, A. Edible Cirina forda (Westwood, 1849) (Lepidoptera: Saturniidae) caterpillar among Moba people of the Savannah Region in North Togo: From collector to consumer. Asian J. Appl. Sci. Eng. 2014, 3. [Google Scholar] [CrossRef]
- Latham, P. Edible Caterpillars and Their Food Plants in Bas-Congo Province, Democratic Republic of Congo, 3rd ed.; Paul Latham: Perthshire, UK, 2015; p. 44. [Google Scholar]
- Glew, R.H.; Jackson, D.; Sena, L.; VanderJagt, D.J.; Pastuszyn, A.; Millson, M. Gonimbrasia belina (Lepidoptera: Saturniidae): A nutritional food source rich in protein, fatty acids, and minerals. Am. Entomol. 1999, 45, 250–253. [Google Scholar] [CrossRef][Green Version]
- Makhado, R.; Potgieter, M.; Timberlake, J.; Gumbo, D. A review of the significance of mopane products to rural people’s livelihoods in Southern Africa. Trans. R. Soc. South Afr. 2014, 69, 117–122. [Google Scholar] [CrossRef]
- Baiyegunhi, L.J.S.; Oppong, B.B. Commercialisation of mopane worm (Imbrasia belina) in rural households in Limpopo Province, South Africa. For. Policy Econ. 2016, 62, 141–148. [Google Scholar] [CrossRef]
- Hlongwane, Z.T.; Slotow, R.; Munyai, T.C. Nutritional composition of edible insects consumed in Africa: A systematic review. Nutrients 2020, 12, 2786. [Google Scholar] [CrossRef]
- Scoble, M. The Lepidoptera Form Function, and Diversity; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Oberprieler, R. The Emperor Moths of Namibia; Ekogilde: Pretoria, South Africa, 1995. [Google Scholar]
- Stack, J.; Dorward, A.; Gondo, T.; Frost, P.; Taylor, F.; Kurebgaseka, N. Mopane worm utilisation and rural livelihoods in Southern Africa. In Proceedings of the International Conference on Rural Livelihoods, Forests and Biodiversity, Bonn, Germany, 19–23 May 2003. [Google Scholar]
- Fasoranti, J.O.; Ajiboye, D.O. Some edible insects of Kwara state, Nigeria. Entomol. Soc. Am. 1993, 39, 113–116. [Google Scholar] [CrossRef]
- Defoliart, G.R. Edible insects as minilivestock. Biodivers. Conserv. 1995, 4, 306–321. [Google Scholar] [CrossRef]
- Nyoka, B.I. State of Forest and Tree Genetic Resources in Dry Zone Southern Africa Development Community Countries; Working Paper FGR/41E; Food and Agriculture Organization of the United Nations: Rome, Italy, 2003. [Google Scholar]
- Ditlhogo, M. The Natural History of Imbrasia belina (Westwood) (Lepidoptera: Saturniidae) and Some Factors Affecting Its Abundance in North-Eastern Botswana. Ph.D. Thesis, University of Manitoba, Winnipeg, MB, Canada, 1996. [Google Scholar]
- Ande, A.T.; Fasoranti, J.O. Life history notes for the pallid emperor moth, Cirina forda (Westwood) (Lepidoptera: Saturniidae). J. Lepidoterists Soc. 1997, 51, 269–271. [Google Scholar]
- Balinga, M.; Mapunzu, P.M.; Moussa, J.B.; N’Gasse, G. Contribution Des Insectes De La Forêt À La Sécurité Alimentaire: L’exemple Des Chenilles d’Afrique Centrale; Food and Agriculture Organization of The United Nations: Rome, Italy, 2004; p. 117. (In French) [Google Scholar]
- Moruakgomo, M.B.W. Commercial utilization of Botswana’s veld products the economics of phane: The dimensions of phane trade. In Proceedings of the First Multidisciplinary Symposium on Phane, Gaborone, Botswana, 18 June 1996; The Department of Biological Sciences and the Kalahari Conservation Society: Gaborone, Botswana, 1996; pp. 23–31. [Google Scholar]
- Ande, A.T.; Fasoranti, J.O. Some aspects of the biology, foraging and defensive behaviour of the emperor moth caterpillar, Cirina forda (Westwood). Int. J. Trop. Insect Sci. 1998, 18, 177–181. [Google Scholar] [CrossRef]
- Dwomoh, E.A.; Ahadzie, S.K.; Agene, V.N.; Somuah, G.A.; Ackonor, J.B. Biology and phenology of the shea tree defoliator Cirina forda (Westwood) (Lepidoptera: Saturniidae) in Northern Ghana. Ghana J. Hortic. 2010, 8, 55–64. [Google Scholar]
- Omotoso, O.T. Nutritional quality, functional properties and anti-nutrient compositions of the larva of Cirina forda (Westwood) (Lepidoptera: Saturniidae). J. Zhejiang Univ. Sci. B 2006, 7, 51–55. [Google Scholar] [CrossRef]
- Womeni, H.M.; Linder, M.; Tiencheu, B.; Mbiapo, F.T.; Villeneuve, P.; Fanni, J.; Parmentier, M. Oils of insects and larvae consumed in Africa: Potential sources of polyunsaturated fatty acids. Ol. Corps Gras Lipides 2009, 16, 230–235. [Google Scholar] [CrossRef]
- Adepoju, O.T.; Daboh, O.O. Nutrient composition of Cirina forda (Westwood)-enriched complementary foods. Ann. Nutr. Metab. 2013, 63, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Vantomme, P.; Göhler, D.; N’Deckere-Ziangba, F. Contribution of Forest Insects to Food Security and Forest Conservation: The Example of Caterpillars in Central Africa; Food and Agriculture Organization of The United Nations: Rome, Italy, 2004; p. 4. [Google Scholar]
- Sombroek, W.G.; Braun, H.M.H.; van der Pouw, B.J.A. Exploratory Soil Map and Agro-Climatic Zone Map of Kenya; Ministry of Agriculture: Nairobi, Kenya, 1982; p. 56.
- FAO. Agro-Ecological Zoning Guidelines; FAO Soils Bulletin; Food and Agriculture Organization of the United Nations: Rome, Italy, 1996; p. 78. [Google Scholar]
- Kuhne, L. Butterflies and Moth Diversity of the Kakamega Forest; Selbstverl: Postdam, Germany, 2008. [Google Scholar]
- Goff, R. African Moths. Available online: https://www.africanmoths.com/ (accessed on 26 September 2019).
- Beentje, H.; Adamson, J.; Bhanderi, D. Kenya Trees, Shrubs, and Lianas; National Museums of Kenya: Nairobi, Kenya, 1994. [Google Scholar]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janze, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rantnasingham, S.; Herbert, P.D.N. Bold: The barcode of life data system. Mol. Ecol. Notes 2007, 7, 355. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 1547–1549. [Google Scholar] [CrossRef]
- Dauda, B.E.N.; Mathew, J.T.; Paiko, Y.B.; Ndamitso, M.M. Nutritive and anti-nutritive composition of locust bean tree emperor moth larvae Bunaea alcinoe (Lepidoptera-Saturniidae Stoll 1780) from Gurara local government area, Niger state, Nigeria. J. Sci. Res. Rep. 2014, 3, 1771–1779. [Google Scholar] [CrossRef]
- Paiko, Y.B.; Jacob, J.O.; Salihu, S.O.; Dauda, B.E.N.; Suleiman, M.A.T.; Akanya, H.O. Fatty acid and amino acid profile of emperor moth caterpillar (Cirina forda) in Paikoro local government area of Niger state, Nigeria. Am. J. Biochem. 2014, 4, 29–34. [Google Scholar]
- Mabossy-Mobouna, G.; Bouyer, T.; Latham, P.; Roulon-Doko, P.; Konda Ku Mbuta, A.; Malaisse, F. Preliminary knowledge for breeding edible caterpillars in Congo-Brazzaville. Geo Eco Trop Rev. Int. Géol. Géograph. Écol. Trop. 2016, 40, 145–174. [Google Scholar]
- Agbidye, F.S.; Ofuya, T.I.; Akindele, S.O. Marketability and nutritional qualities of some edible forest insects in Benue state, Nigeria. Pak. J. Nutr. 2009, 8, 917–922. [Google Scholar] [CrossRef]
- Ehounou, G.P.; Ouali-N’goran, S.W.M.; Niassy, S. Assessment of entomophagy in Abidjan (Cote Divoire, West Africa). Afr. J. Food Sci. 2018, 12, 6–14. [Google Scholar] [CrossRef]
- Ngute, A.S.K.; Dongmo, M.A.K.; Effa, J.A.M.; Onguene, E.M.A.; Lontchi, J.F.; Cuni-Sanchez, A. Edible caterpillars in central Cameroon: Host plants, value, harvesting, and availability. For. Trees Livelihoods 2020, 29, 16–33. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Wei, B.; Wang, R.; Hou, K.; Wang, X.; Wu, W. Predicting the current and future cultivation regions of Carthamus tinctorius L. using MaxEnt model under climate change in China. Glob. Ecol. Conserv. 2018, 16, e00477. [Google Scholar] [CrossRef]
- Mbata, K.J.; Chidumayo, E.N.; Lwatula, C.M. Traditional regulation of edible caterpillar exploitation in the Kopa area of Mpika district in northern Zambia. J. Insect Conserv. 2002, 6, 115–130. [Google Scholar] [CrossRef]
- Amadi, E.N.; Ogbalu, O.K.; Barimalaa, I.S.; Pius, M. Microbiology and nutritional composition of an edible larva (Bunaea alcinoe Stoll) of the Niger delta. J. Food Saf. 2005, 25, 193–197. [Google Scholar] [CrossRef]
- Temitope, A.O.; Job, O.O.; Abiodun, A.-F.T.; Dare, A.O. Eco-diversity of edible insects of Nigeria and its impact on food security. J. Biol. Life Sci. 2014, 5, 175–187. [Google Scholar] [CrossRef]
- Kachapulula, P.W.; Akelo, J.; Bandyopathyay, R.; Cotty, P.J. Aflatoxin contamination of dried insects and fish in Zambia. J. Food Prot. 2018, 81, 1508–1518. [Google Scholar] [CrossRef]
- Siozios, S.; Massa, A.; Parr, C.L.; Verspoor, R.L.; Hurst, G.D.D. DNA barcoding reveals incorrect labelling of insects sold as food in the UK. PeerJ 2020, 8. [Google Scholar] [CrossRef]
- Akanbi, M.O. Comparing two saturniid defoliators of Holarrhena floribunda and Ekebergia sengalensis, respectively, with notes on their natural enemies. J. Nat. Hist. 1973, 7, 307–318. [Google Scholar] [CrossRef]
- Thomas, B. Sustainable harvesting and trading of mopane worms (Imbrasia belina) in northern Namibia: An experience from the Uukwaluudhi area. Int. J. Environ. Stud. 2013, 70, 494–502. [Google Scholar] [CrossRef]
- Kwiri, R.; Winini, C.; Muredzi, P.; Tongonya, J.; Gwala, W.; Mujuru, F.; Gwala, S.T. Mopane worm (Gonimbrasia belina) utilisation, a potential source of protein in fortified blended foods in Zimbabwe: A review. Glob. J. Sci. Front. Res. 2014, 14, 1–14. [Google Scholar]
- De Almeida, D.M. Pests of Mango. Gaz. Agric. 1972, 24, 2–5. [Google Scholar]
- Bussmann, R.W.; Gilbreath, G.G.; Solio, J.; Lutura, M.; Lutuluo, R.; Kunguru, K.; Wood, N.; Mathenge, S.G. Plant use of the Maasai of Sekenani valley, Maasai Mara, Kenya. J. Ethnobiol. Ethnomedicine 2006, 2, 22. [Google Scholar] [CrossRef]
- Kipkore, W.; Wanjohi, B.; Rono, H.; Kigen, G. A study of the medicinal plants used by the Marakwet community in Kenya. J. Ethnobiol. Ethnomed. 2014, 10, 24. [Google Scholar] [CrossRef]
- Johns, T.; Kokwaro, J.O.; Kimanani, E.K. Herbal remedies of the Luo of Siaya district, Kenya: Establishing quantitative criteria for consensus. Econ. Bot. 1990, 44, 369–381. [Google Scholar] [CrossRef]
- Tian, X. Ethnobotanical knowledge acquisition during daily chores: The firewood collection of pastoral Maasai girls in southern Kenya. J. Ethnobiol. Ethnomed. 2017, 13, s13002–s13016. [Google Scholar] [CrossRef] [PubMed]
- Orwa, C.; Mutua, A.; Kindt, R.; Jamnadass, R.; Anthony, S. Agroforestry Database: A Tree Reference and Selection Guide Version 4.0; World Agroforestry Centre: Nairobi, Kenya, 2009. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Characteristics | |||
|---|---|---|---|---|
| Moth Color | Hindwing Eyespot Description (Innermost to Outermost Color) | Mature Larvae Color | 5th Instar Spine Color | |
| Gonimbrasia zambesina | Green | Greenish-yellow center, black, greenish-yellow, white rings | Black with grey and yellow speckles | Black or red |
| Gonimbrasia zambesina | Brown | Yellowish brown center, black, pink, white rings | Black with grey and yellow speckles | Red |
| Gonimbrasia cocaulti | Brown | White center, reddish, black and white rings | Black with whitish speckles | Yellow |
| Gonimbrasia krucki | Yellow | Yellow center, black, pink, red rings | Black with greenish-yellow speckles and orange spots on spiracles | Black |
| Cirina forda | Light Brown | Small with a black ring | Black with yellow bands | White |
| Gonimbrasia belina | Reddish-Brown | Brown center, black, white rings | Black with red, grey and green speckles | Black |
| Bunaea alcinoe | Dark brown | Orange center, black, white rings | Black with orange spots on spiracles | White/yellow |
| Bunaea alcinoe | - | - | Red | White |
| Gynanisa nigra | - | - | Green with white speckles | White |
| Gonimbrasia belina | - | - | Black with red, grey and greenish speckles | Black |
| Collection Site | Sample Code | Species | % Similarity to BOLD Sequences | Sequence ID of Similar Sequences-Top Published Hit with Default BOLD Query (Collection Site) |
|---|---|---|---|---|
| Mwingi, Kenya | S30 | Go. cocaulti | 98.88 | SPBIS152-09 (Kenya) |
| Taita, Kenya | S34 | Go. cocaulti | 98.88 | SPBIS152-09 (Kenya) |
| Matuu, Kenya | S22 | Go. cocaulti | 98.72 | SPBIS152-09 (Kenya) |
| Taita, Kenya | S32 | Go. cocaulti | 98.87 | SPBIS152-09 (Kenya) |
| Taita, Kenya | S33 | Go. cocaulti | 98.88 | SPBIS152-09 (Kenya) |
| Matuu, Kenya | S28 | Go. cocaulti | 98.72 | SPBIS152-09 (Kenya) |
| Muhaka, Kenya | S48 | Go. belina | 100 | GBMNC60703-20 (Kenya) |
| Ibadan, Nigeria | Nigeria-1 | B. alcinoe | 100 | SATWA891-07 (Burkina-Faso) |
| Matuu, Kenya | S84 | B. alcinoe | 99.24 | LSAFR2238-12 (South Africa) |
| Matuu, Kenya | S85 | B. alcinoe | 99.39 | LSAFR2238-12 (South Africa) |
| Nanyuki, Kenya | S87 | B. alcinoe | 99.24 | LSAFR2238-12 (South Africa) |
| Nanyuki, Kenya | S88 | B. alcinoe | 99.24 | LSAFR2238-12 (South Africa) |
| Mbita, Kenya | S90 | B. alcinoe | 99.39 | LSAFR2238-12 (South Africa) |
| Mbita, Kenya | S91 | B. alcinoe | 99.08 | LSAFR2238-12 (South Africa) |
| Embu, Kenya | S92 | B. alcinoe | 99.23 | LSAFR2238-12 (South Africa) |
| Embu, Kenya | S93 | B. alcinoe | 98.78 | LSAFR2238-12 (South Africa) |
| Nairobi, Kenya | S2 | Go. krucki | 100 | SAPBA635-07 (Kenya) |
| Nairobi, Kenya | S3 | Go. krucki | 100 | SAPBA635-07 (Kenya) |
| Mbita, Kenya | S6 | C. forda | 99.54 | SATWA281-07 (Cameroon) |
| Mbita, Kenya | S7 | C. forda | 99.54 | SATWA281-07 (Cameroon) |
| Kilifi, Kenya | S55 | C. forda | 99.41 | STBOB620-08 (Malawi) |
| Ngong, Kenya | 2CF | C. forda | 99.38 | STBOB620-08 (Malawi) |
| Ngong, Kenya | 5CF | C. forda | 99.69 | STBOB620-08 (Malawi) |
| Gilgil, Kenya | S54 | C. forda | 99.85 | STBOB620-08 (Malawi) |
| Kilifi, Kenya | S17 | Go. zambesina | 100 | SAPBA773-07 (Kenya) |
| Kilifi, Kenya | S18 | Go. zambesina | 100 | SAPBA773-07 (Kenya) |
| Kambiti, Kenya | S79 | Go. zambesina | 99.54 | SAPBA773-07 (Kenya) |
| Kambiti, Kenya | S94 | Go. zambesina | 99.39 | SAPBA773-07 (Kenya) |
| Embu, Kenya | S95 | Go. zambesina | 99.39 | SAPBA773-07 (Kenya) |
| Embu, Kenya | S96 | Go. zambesina | 99.58 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S97 Brown | Go. zambesina | 99.54 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S98 Brown | Go. zambesina | 99.54 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S99 Green | Go. zambesina | 99.54 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S100 Green | Go. zambesina | 99.09 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S101 Green | Go. zambesina | 99.39 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S102 Green | Go. zambesina | 99.39 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S104 Brown | Go. zambesina | 99.39 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S105 Brown | Go. zambesina | 99.54 | SAPBA773-07 (Kenya) |
| Makuyu, Kenya | S107 Brown | Go. zambesina | 99.24 | SAPBA773-07 (Kenya) |
| Botswana | IBB-1 | Go. belina | 99.84 | SATWA003-06 (Zambia) |
| Botswana | IBB-2 | Go. belina | 99.85 | SATWA003-06 (Zambia) |
| Kenya | GMB-2 | Gy. nigra | 100 | GBMNC60687-20 (Botswana) |
| Botswana | GM-1 | Gy. westwoodi | 99.69 | STBOA580-07 (Kenya) |
| Botswana | GM-2 | Gy. westwoodi | 99.69 | STBOA580-07 (Kenya) |
| Insect Species (Sample Size) | Genetic p-Distance Range within Sample Species | BOLD Sequence Used for Comparison (BOLD BIN Cluster Number) | Genetic p-Distance Range between Sample Species and BOLD Sequence |
|---|---|---|---|
| Gonimbrasia zambesina (15) | 0–0.61 | SAPBA773-07 (BOLD:AAD1339) | 0–0.91 |
| Gonimbrasia krucki (2) | 0.0 | SAPBA635-07 (BOLD:AAD8374) | 0.0 |
| Gonimbrasia belina-Kenya (1) | 0.0 | GBMNC60703-20 (BOLD:AAB6786) | 0.0 |
| Gonimbrasia cocaulti (7) | 0–1.52 | SPBIS152-09 (BOLD:AEH8028) | 1.42–2.88 |
| Gonimbrasia belina-Botswana (2) | 0.0 | SATWA003-06 (BOLD:AAB6786) | 0.15–0.16 |
| Cirina forda (4) | 0.3–2.13 | SATWA281-07 (BOLD:AAB6982) | 0.46–2.13 |
| Bunaea alcinoe-Kenya (8) | 0–1.52 | LSAFR2238-12 (BOLD:AAA6757) | 0.61–1.98 |
| Bunaea alcinoe-Nigeria (1) | 0.0 | SATWA891-07 (BOLD:AAA6756) | 0.0 |
| Gynanisa nigra (1) | 0.0 | STBOC836-08 (BOLD:AED6623) | 0.61 |
| Gynanisa westwoodi (2) | 0.0 | STBOA580-07 (BOLD:ABY4629) | 0.3 |
| Saturniid | Place Found | April–June | October–December |
|---|---|---|---|
| Gonimbrasia zambesina | Kilifi, Embu, Machakos, Kwale, Murang’a | Present | Present |
| Cirina forda | Kilifi, Nakuru, Embu, HomaBay, Kajiado | Present | Present |
| Gonimbrasia cocaulti | Taita, Makueni, Machakos, Kitui, Isiolo | Present | Absent |
| Bunaea alcinoe | Machakos, Makueni, Homabay, Meru, Kitui, Embu, Laikipia | Present | Present |
| Gonimbrasia krucki | Nairobi | Present | Present |
| Gonimbrasia belina | Kwale | Present | Present |
| Species | Current | RCP8.5 | ||
|---|---|---|---|---|
| Training | Test | Training | Test | |
| Bunaea alcinoe | 0.855 | 0.915 | 0.877 | 0.928 |
| Cirina forda | 0.850 | 0.867 | 0.876 | 0.860 |
| Saturniid | Host Plant |
|---|---|
| Gonimbrasia zambesina | Mangifera indica, Anacardium occidentale |
| Cirina forda | Euclea divinorum, Acacia mearnsii, Manilkara sulcata |
| Gonimbrasia cocaulti | Vachellia tortilis, Vachellia nilotica |
| Bunaea alcinoe | Balanites aegyptiaca, Balanites glabra |
| Gonimbrasia krucki | Schinus terebinthifolia, Schinus molle |
| Gonimbrasia belina | Anacardium occidentale |
| Collection Site | Sample Code | Species | % Similarity to GenBank Sequences | ID of Similar Sequences |
|---|---|---|---|---|
| Gilgil, Nakuru | HP7 | Euclea divinorum | 100 | DQ924074.1 |
| Mbita, Homabay | HP17 | Euclea divinorum | 97.06 | DQ924074.1 |
| Mbita, Homabay | HP18 | Euclea divinorum | 98.11 | DQ924074.1 |
| Gilgil, Nakuru | HP37 | Euclea divinorum | 99.64 | DQ924074.1 |
| Embu | HP38 | Euclea divinorum | 99.76 | DQ924074.1 |
| Gilgil, Nakuru | HP39 | Euclea divinorum | 100 | DQ924074.1 |
| Embu | HP40 | Euclea divinorum | 99.76 | DQ924074.1 |
| Matuu, Machakos | HP21 | Vachellia tortilis | 99.54 | AF274140.1 |
| Taita | HP22 | Vachellia tortilis | 99.77 | AF274140.1 |
| Makueni | HP23 | Vachellia tortilis | 99.77 | AF274140.1 |
| Mwingi | HP24 | Vachellia tortilis | 99.88 | AF274140.1 |
| Ngong, Kajiado | HP25 | Acacia mearnsii | 99.88 | HM020723.1 |
| Ngong, Kajiado | HP26 | Acacia mearnsii | 99.76 | HM020723.1 |
| Ngong, Kajiado | HP27 | Acacia mearnsii | 100 | HM020723.1 |
| Ngong, Kajiado | HP28 | Acacia mearnsii | 100 | HM020723.1 |
| Kilifi, Malindi | HP16 | Manilkara sp. | 99.40 | DQ924092.1 |
| Kilifi, Malindi | HP33 | Manilkara sp. | 99.40 | DQ924092.1 |
| Kilifi, Malindi | HP36 | Manilkara sp. | 99.40 | DQ924092.1 |
| Muhaka, Kwale | HP11 | Anacardium occidentale | 100 | KY635877.1 |
| Malindi, Kilifi | HP10 | Mangifera indica | 100 | KX871231.1 |
| Mbita, Homabay | HP3 | Balanites sp. | 99.35 | JX517722.1 |
| Mbita, Homabay | HP32 | Balanites sp. | 99.22 | JX517722.1 |
| Matuu, Machakos | HP45 | Balanites sp. | 99.48 | JX517722.1 |
| Matuu, Machakos | HP46 | Balanites sp. | 99.48 | JX517722.1 |
| Nanyuki | HP47 | Balanites sp. | 99.48 | JX517722.1 |
| Embu | HP48 | Balanites sp. | 99.48 | JX517722.1 |
| Embu | HP45 | Balanites sp. | 99.48 | JX517722.1 |
| Matuu, Machakos | HP2 | Vachelia nilotica | 99.30 | KY10024.1 |
| Buruburu, Nairobi | HP49 | Schinus terebinthifolia | 100 | KP149521.1 |
| Buruburu, Nairobi | HP51 | Schinus terebinthifolia | 100 | KP149521.1 |
| Kasarani, Nairobi | HP63 | Schinus terebinthifolia | 100 | KP149521.1 |
| Kasarani, Nairobi | HP62 | Schinus terebinthifolia | 100 | KP149521.1 |
| Kasarani, Nairobi | HP60 | Schinus terebinthifolia | 99.53 | KP149521.1 |
| Kasarani, Nairobi | HP61 | Schinus terebinthifolia | 99.53 | KP149521.1 |
| Kasarani, Nairobi | HP58 | Schinus terebinthifolia | 99.41 | KP149521.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kusia, E.S.; Borgemeister, C.; Khamis, F.M.; Copeland, R.S.; Tanga, C.M.; Ombura, F.L.; Subramanian, S. Diversity, Host Plants and Potential Distribution of Edible Saturniid Caterpillars in Kenya. Insects 2021, 12, 600. https://doi.org/10.3390/insects12070600
Kusia ES, Borgemeister C, Khamis FM, Copeland RS, Tanga CM, Ombura FL, Subramanian S. Diversity, Host Plants and Potential Distribution of Edible Saturniid Caterpillars in Kenya. Insects. 2021; 12(7):600. https://doi.org/10.3390/insects12070600
Chicago/Turabian StyleKusia, Elizabeth Siago, Christian Borgemeister, Fathiya M. Khamis, Robert S. Copeland, Chrysantus M. Tanga, Fidelis Levi Ombura, and Sevgan Subramanian. 2021. "Diversity, Host Plants and Potential Distribution of Edible Saturniid Caterpillars in Kenya" Insects 12, no. 7: 600. https://doi.org/10.3390/insects12070600
APA StyleKusia, E. S., Borgemeister, C., Khamis, F. M., Copeland, R. S., Tanga, C. M., Ombura, F. L., & Subramanian, S. (2021). Diversity, Host Plants and Potential Distribution of Edible Saturniid Caterpillars in Kenya. Insects, 12(7), 600. https://doi.org/10.3390/insects12070600