
Discrimination between Eggs from Stink Bugs Species in Europe Using MALDI-TOF MS



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panizzi, A.R. Stink Bugs (Hemiptera: Pentatomidae), Emphasizing Economic Importance. In Encyclopedia of Entomology, 1st ed.; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 2120–2122. [Google Scholar]
- Hulme, P.H. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- McPherson, J.E.; Bundy, C.S.; Wheeler, A.G., Jr. Overview of the superfamily Pentatomoidea. In Invasive Stink Bugs and Related Species (Pentatomoidea)-Biology, Higher Systematics, Semiochemistry, and Management, 1st ed.; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 9–11. [Google Scholar]
- Maistrello, L.; Vaccari, G.; Caruso, S.; Costi, E.; Bortolini, S.; Macavei, L.; Foca, G.; Ulrici, A.; Bortolotti, P.P.; Nannini, R.; et al. Monitoring of the invasive Halyomorpha halys, a new key pest of fruit orchards in northern Italy. J. Pest. Sci. 2017, 90, 1231–1244. [Google Scholar] [CrossRef]
- Bosco, L.; Moraglio, S.T.; Tavella, L. Halyomorpha halys, a serious threat for hazelnut in newly invaded areas. J. Pest. Sci. 2018, 91, 661–670. [Google Scholar] [CrossRef]
- Wyniger, D.; Kment, P. Key for the separation of Halyomorpha halys (Stål) from similar-appearing pentatomids (Insecta: Heteroptera: Pentatomidae) occurring in Central Europe, with new Swiss records. Mitt. Schweiz. Entomol. Ges. 2010, 83, 261–270. [Google Scholar] [CrossRef]
- Derjanschi, V.; Péricart, J. Hémiptères Pentatomoidea. Volume 1. Faune de France 90, 1st ed.; Fédération Française des Sociétés de Sciences Naturelles: Paris, France, 2005. [Google Scholar]
- Moulet, P. Hémiptères Coreoidea euro-méditerranéens, Faune de France 81, 1st ed.; Fédération Française des Sociétés de Sciences Naturelles: Paris, France, 1995. [Google Scholar]
- Karas, M.; Bachmann, D.; Hillenkamp, F. Influence of the wavelength in high-irradiance ultraviolet laser desorption mass spectrometry of organic molecules. Anal. Chem. 1985, 57, 2935–2939. [Google Scholar] [CrossRef]
- Knochenmuss, R. Ion formation mechanisms in UV-MALDI. Analyst 2006, 131, 966–986. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.E.; Kaleta, E.J.; Arora, A.; Wolk, D.M. Matrix-assisted laser desorption ionization–time of flight mass spectrometry: A fundamental shift in the routine practice of clinical microbiology. Clin. Microbiol. Rev. 2013, 26, 547–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, M.; Brown, Z.; Houldsworth, M.; Borman, A.M.; Johnson, E.M. Rapid identification of 6328 isolates of pathogenic yeasts using MALDI-ToF MS and a simplified rapid extraction procedure that is compatible with the Bruker Biotyper platform and database. Med. Mycol. 2016, 54, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bader, O. MALDI-TOF-MS-based species identification and typing approaches in medical mycology. Proteomics 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Cassagne, C.; Ranque, S.; Normand, A.C.; Fourquet, P.; Thiebault, S.; Planard, C.; Hendrickx, M.; Piarroux, R. Mould routine identification in the clinical laboratory by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. PLoS ONE 2011, 6, e28425. [Google Scholar] [CrossRef]
- Reeve, M.A.; Buddie, A.G.; Pollard, K.M.; Varia, S.; Seier, M.K.; Offord, L.C.; Cock, M.J.W. A highly-simplified and inexpensive MALDI-TOF mass spectrometry sample-preparation method with broad applicability to microorganisms, plants, and insects. J. Biol. Methods 2018, 5, e103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.; Silva, L.P. MALDI-TOF MS profiling approach: How much can we get from it? Front Plant Sci. 2015, 6, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, M.A.; Pollard, K.M.; Kurose, D. Differentiation between closely-related Impatiens spp. and regional biotypes of Impatiens glandulifera using a highly-simplified and inexpensive method for MALDI-TOF MS. Plant Methods 2018, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, M.A.; Buddie, A.G. A simple and inexpensive method for practical storage of field-sample proteins for subsequent MALDI-TOF MS analysis. Plant Methods 2018, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Reeve, M.A.; Pollard, K.M. Discrimination between regional biotypes of Impatiens glandulifera using a simple MALDI-TOF MS-based method for use with seeds. Plant Methods 2019, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Reeve, M.A.; Pollard, K.M. MALDI-TOF MS-based analysis of dried seed proteins immobilized on filter paper. Biol. Methods Protoc. 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Reeve, M.A.; Seehausen, M.L. Discrimination between Asian populations of the parasitoid wasp Ganaspis cf. brasiliensis using a simple MALDI-TOF MS-based method for use with insects. Biol. Methods Protoc. 2019, 4. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Giromini, D.; Pansa, M.G.; Visentin, S.; Tavella, L. Field collection of Egg Parasitoids of Pentatomidae and Scutelleridae in Northwest Italy and Their Efficacy in Parasitizing Halyomorpha Halys under Laboratory Conditions. Entomol. Exp. Appl. 2021, 169, 52–63. Available online: https://onlinelibrary.wiley.com/doi/10.1111/eea.12966 (accessed on 17 June 2021). [CrossRef]
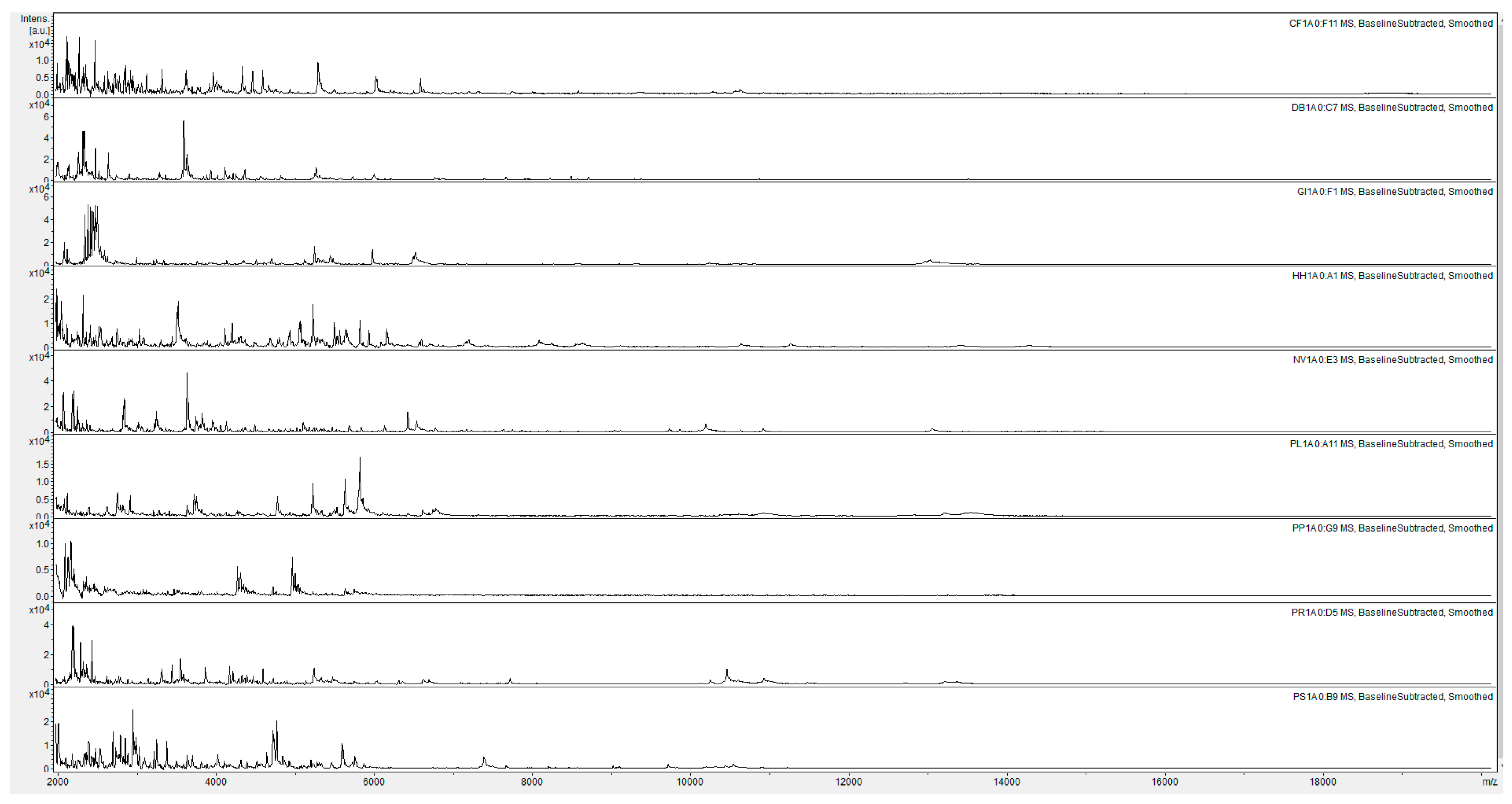


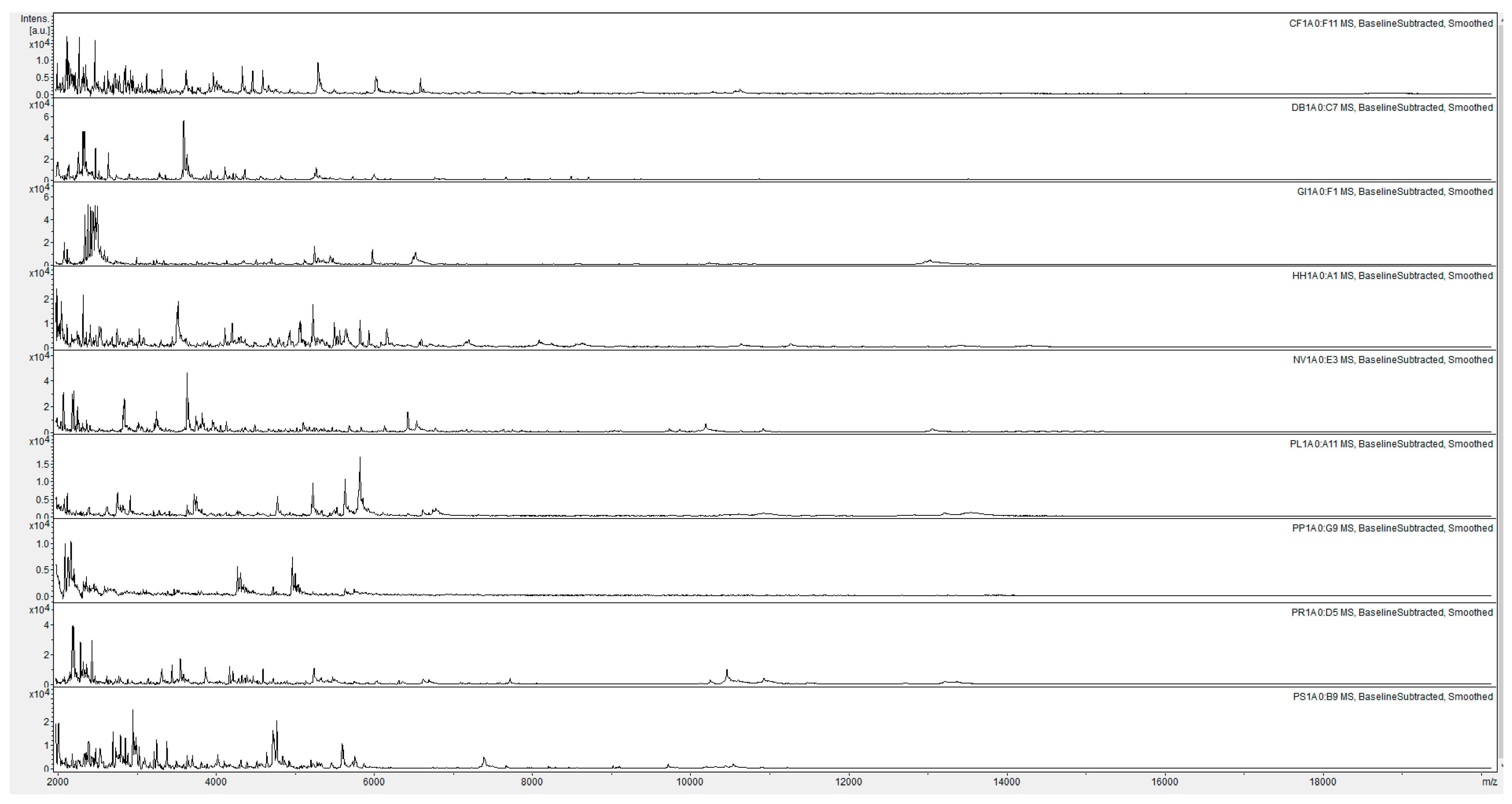{kind=link}
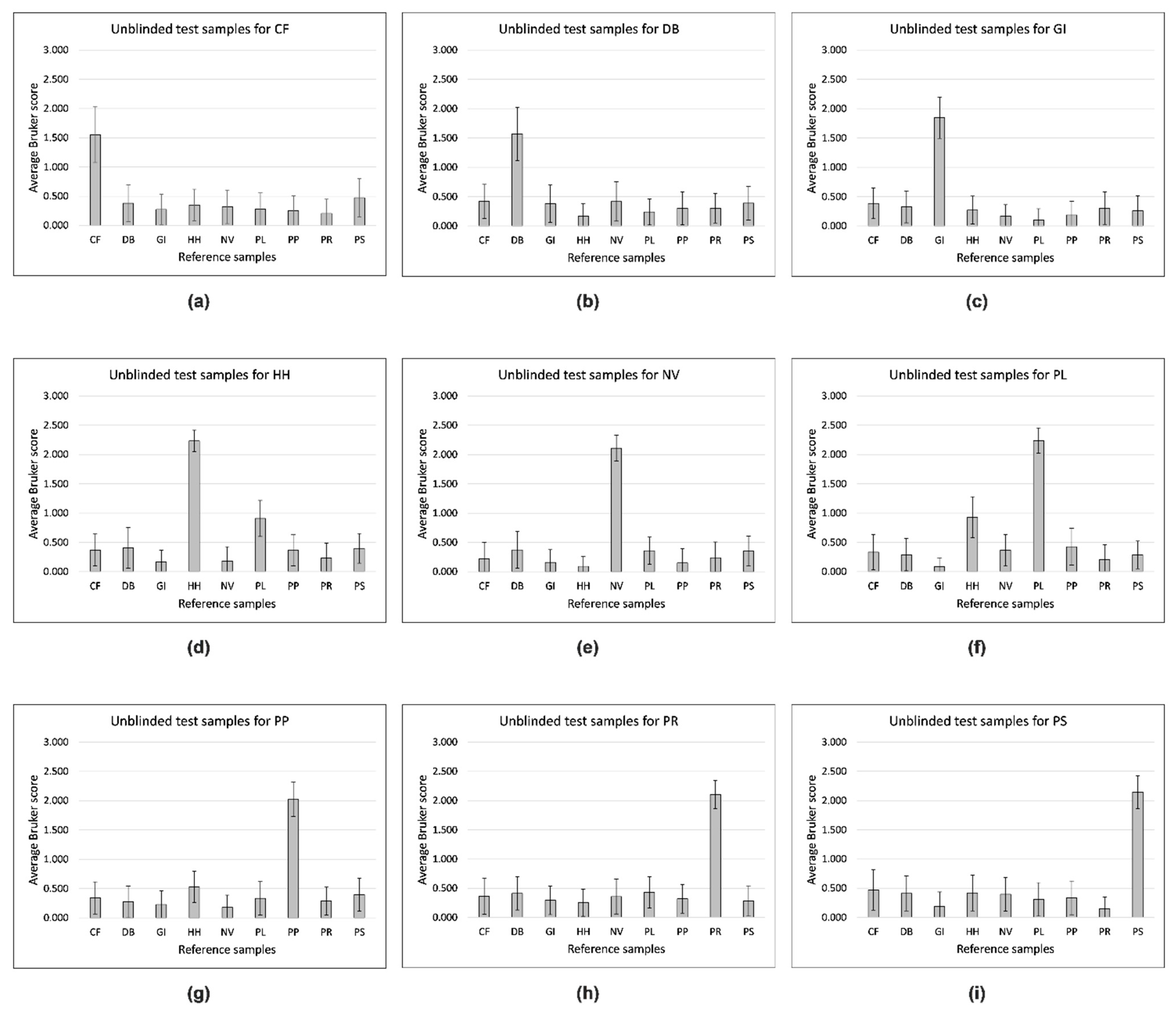
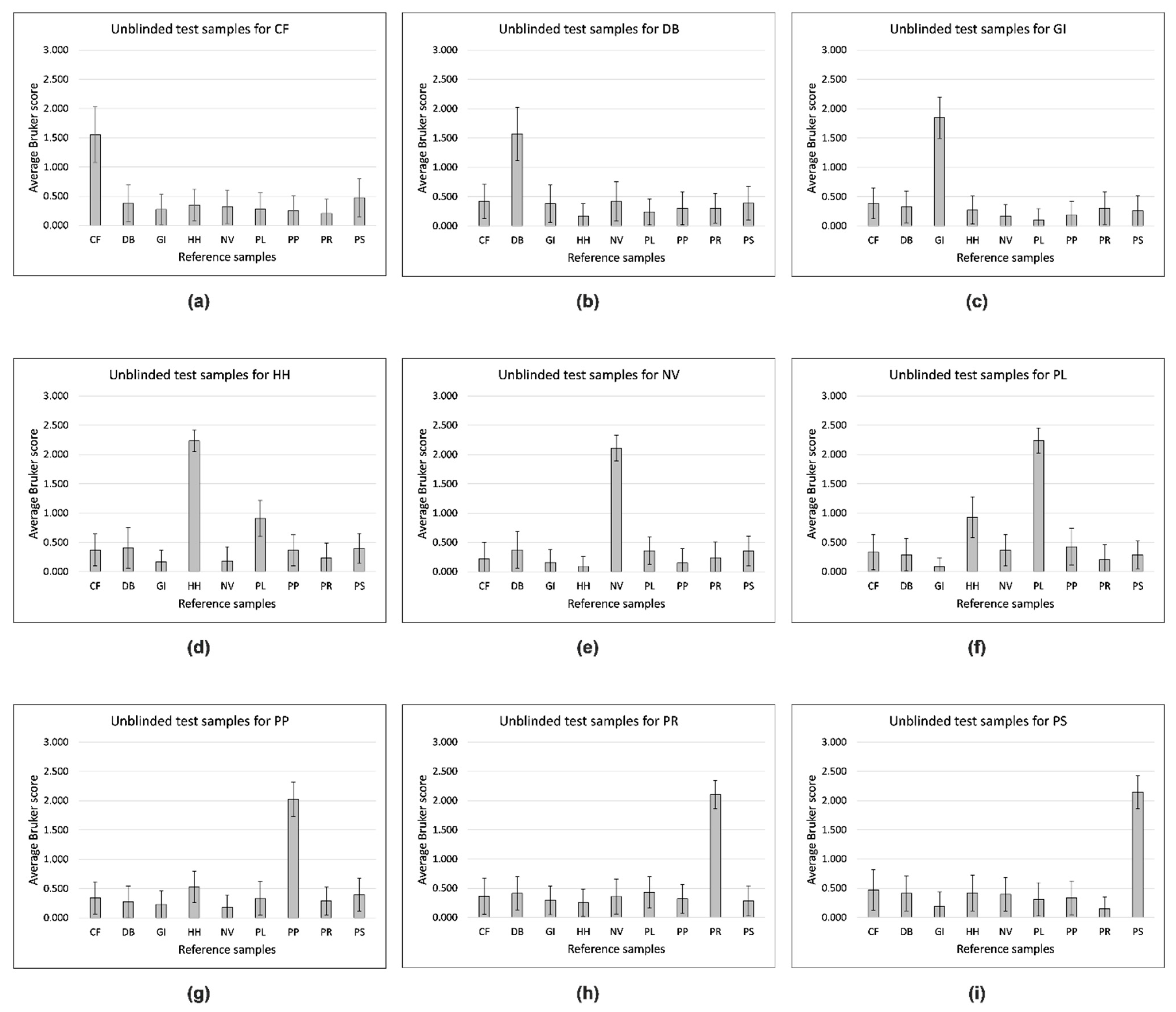{kind=link}
{kind=link}
| Species | Status | Number of Samples | Pest On |
|---|---|---|---|
| Carpocoris fuscispinus (Boheman) | native | 10 | - |
| Dolycoris baccarum (L.) | native | 10 | berry crops |
| Graphosoma lineatum (L.) | native | 10 | - |
| Halyomorpha halys (Stål) | exotic | 10 | fruit crops, vegetables, nuts |
| Nezara viridula (L.) | exotic | 10 | vegetables, fruit crops |
| Palomena prasina (L.) | native | 7 | nuts, fruit crops |
| Pentatoma rufipes (L.) | native | 10 | fruit crops |
| Peribalus strictus (F.) | native | 10 | - |
| Piezodorus lituratus (F.) | native | 10 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reeve, M.A.; Haye, T. Discrimination between Eggs from Stink Bugs Species in Europe Using MALDI-TOF MS. Insects 2021, 12, 587. https://doi.org/10.3390/insects12070587
Reeve MA, Haye T. Discrimination between Eggs from Stink Bugs Species in Europe Using MALDI-TOF MS. Insects. 2021; 12(7):587. https://doi.org/10.3390/insects12070587
Chicago/Turabian StyleReeve, Michael A., and Tim Haye. 2021. "Discrimination between Eggs from Stink Bugs Species in Europe Using MALDI-TOF MS" Insects 12, no. 7: 587. https://doi.org/10.3390/insects12070587
APA StyleReeve, M. A., & Haye, T. (2021). Discrimination between Eggs from Stink Bugs Species in Europe Using MALDI-TOF MS. Insects, 12(7), 587. https://doi.org/10.3390/insects12070587