
Identification of 35 C-Type Lectins in the Oriental Armyworm, Mythimna separata (Walker)


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Microorganisms
2.2. RNA Sample Preparation, Library Construction, and Sequencing
2.3. De Novo Transcriptome Analysis Flow
2.4. Identification of CTLs from the Transcriptome and Bioinformatic Analyses
2.5. Analyses of the Expression Profiles by Real-Time qPCR (RT-qPCR)
3. Results
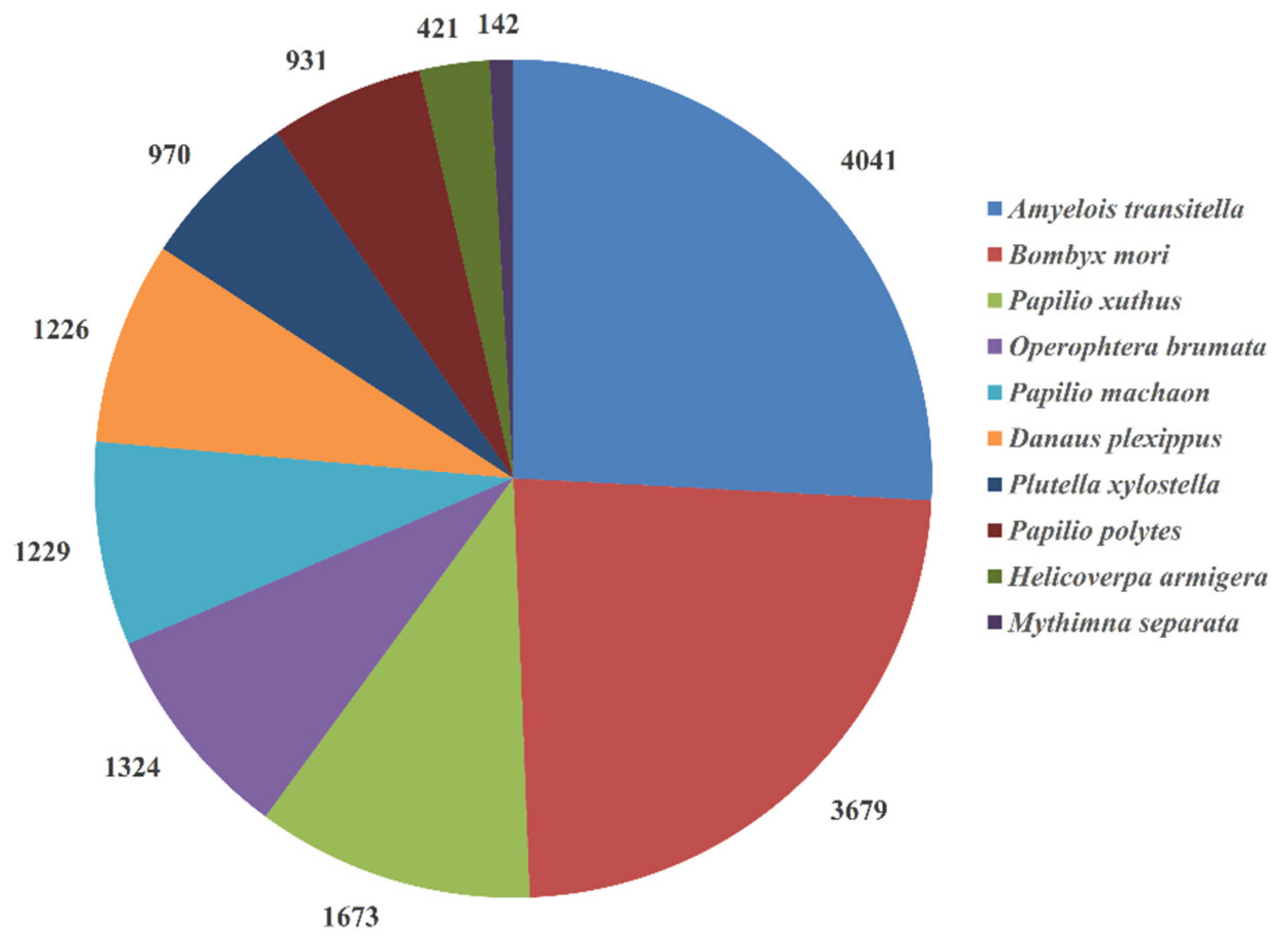
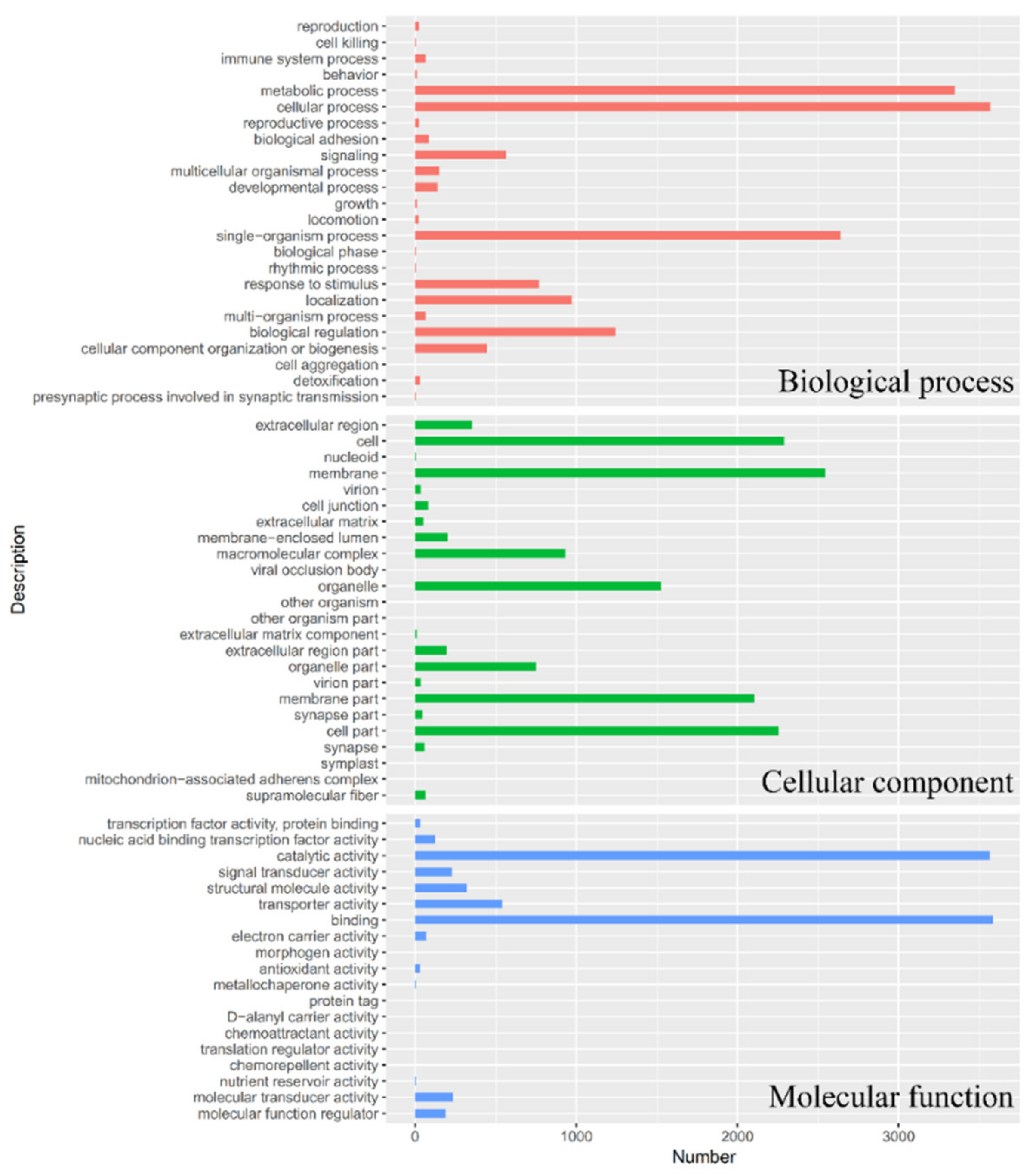
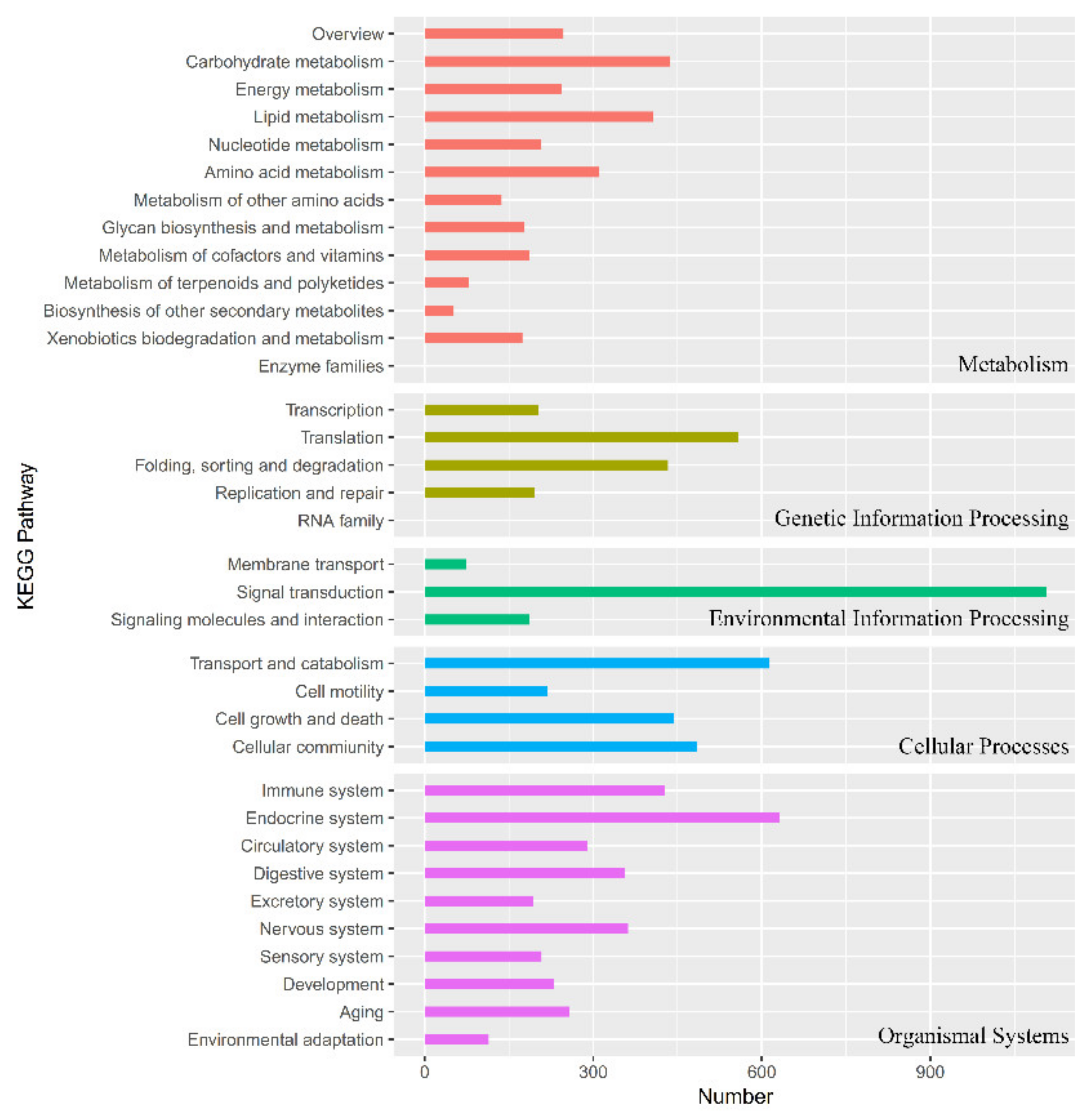
3.1. Transcriptome Sequencing, Unigene Assembly, and Functional Annotation
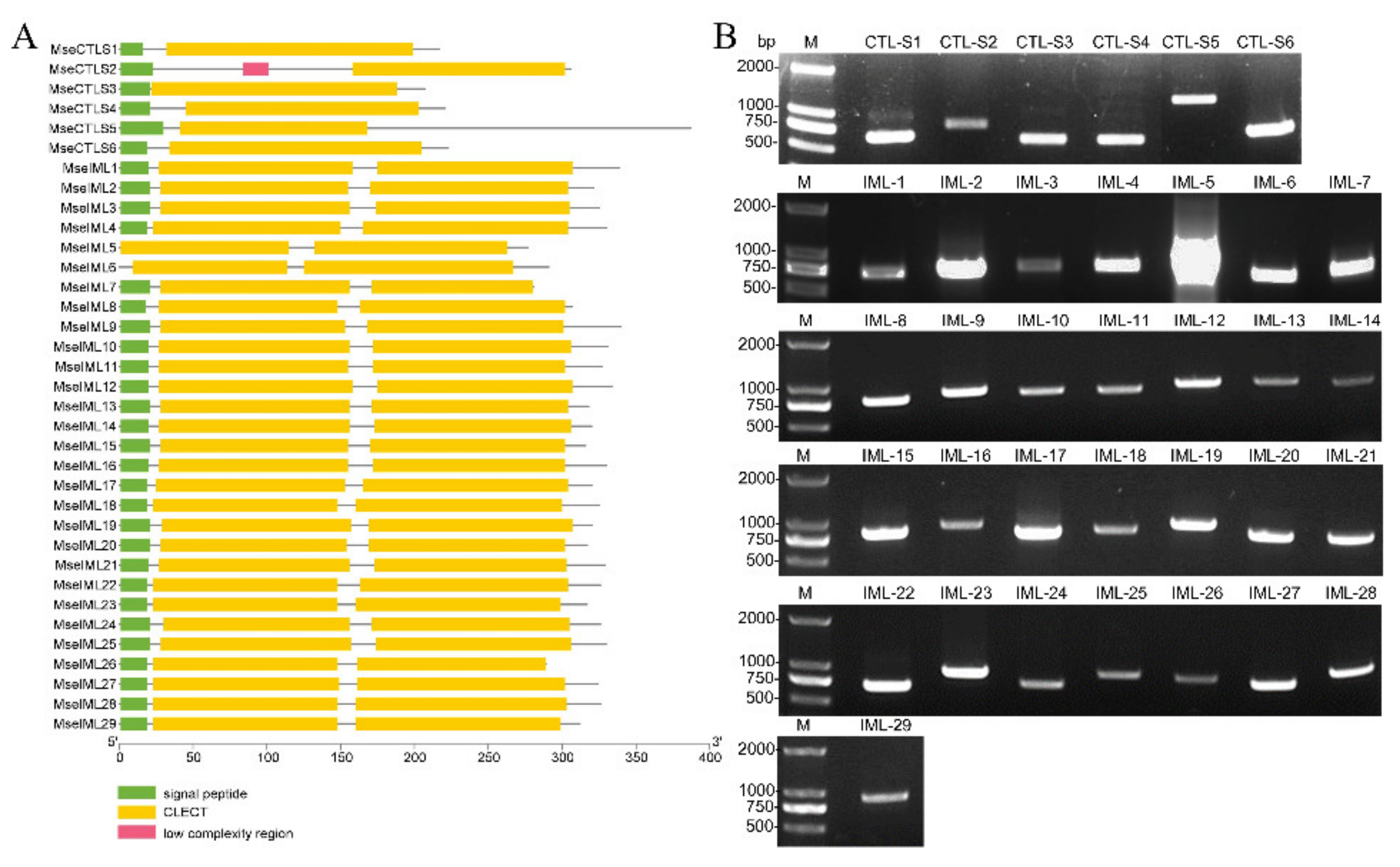
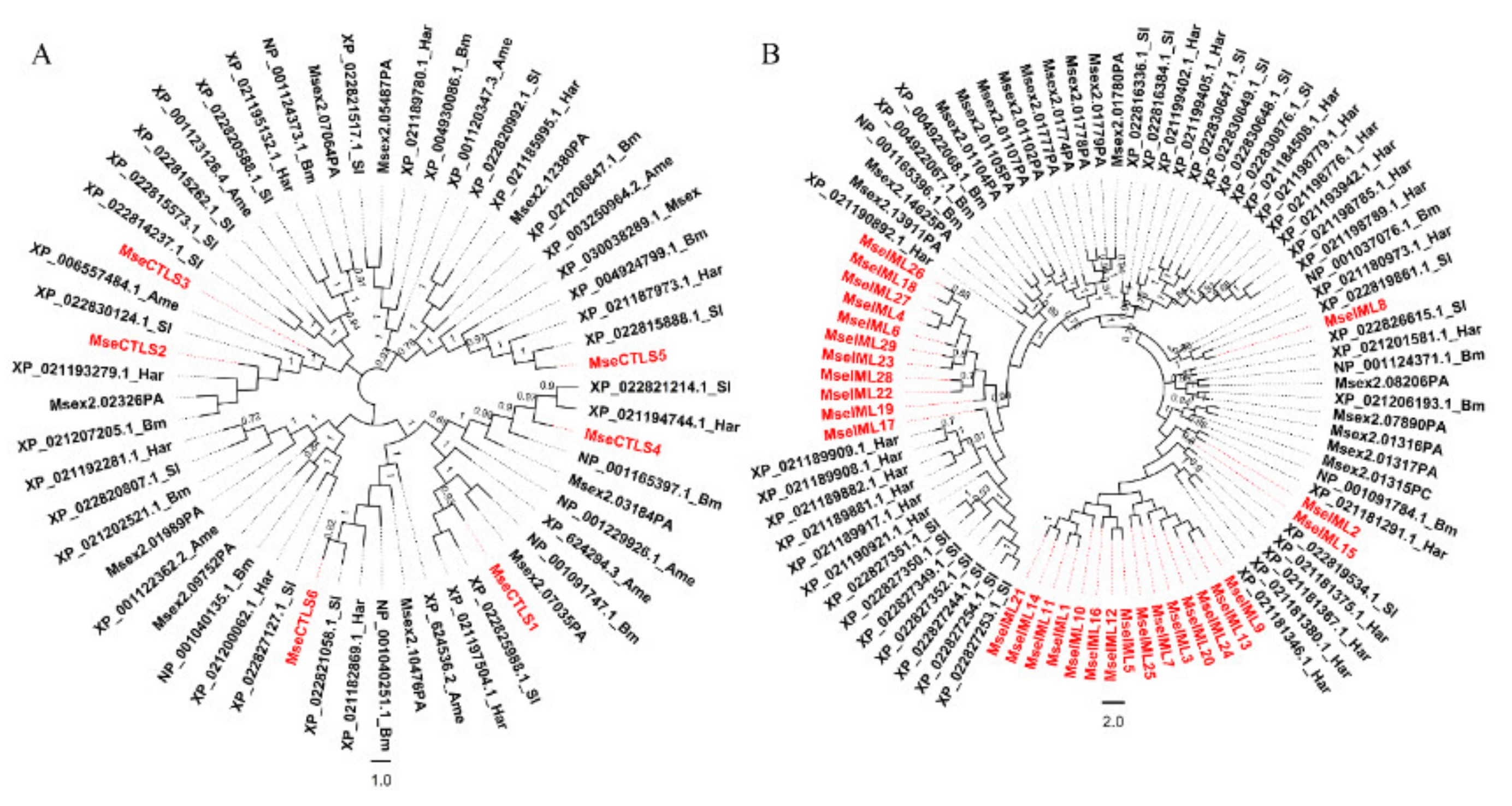
3.2. Identification of CTLs
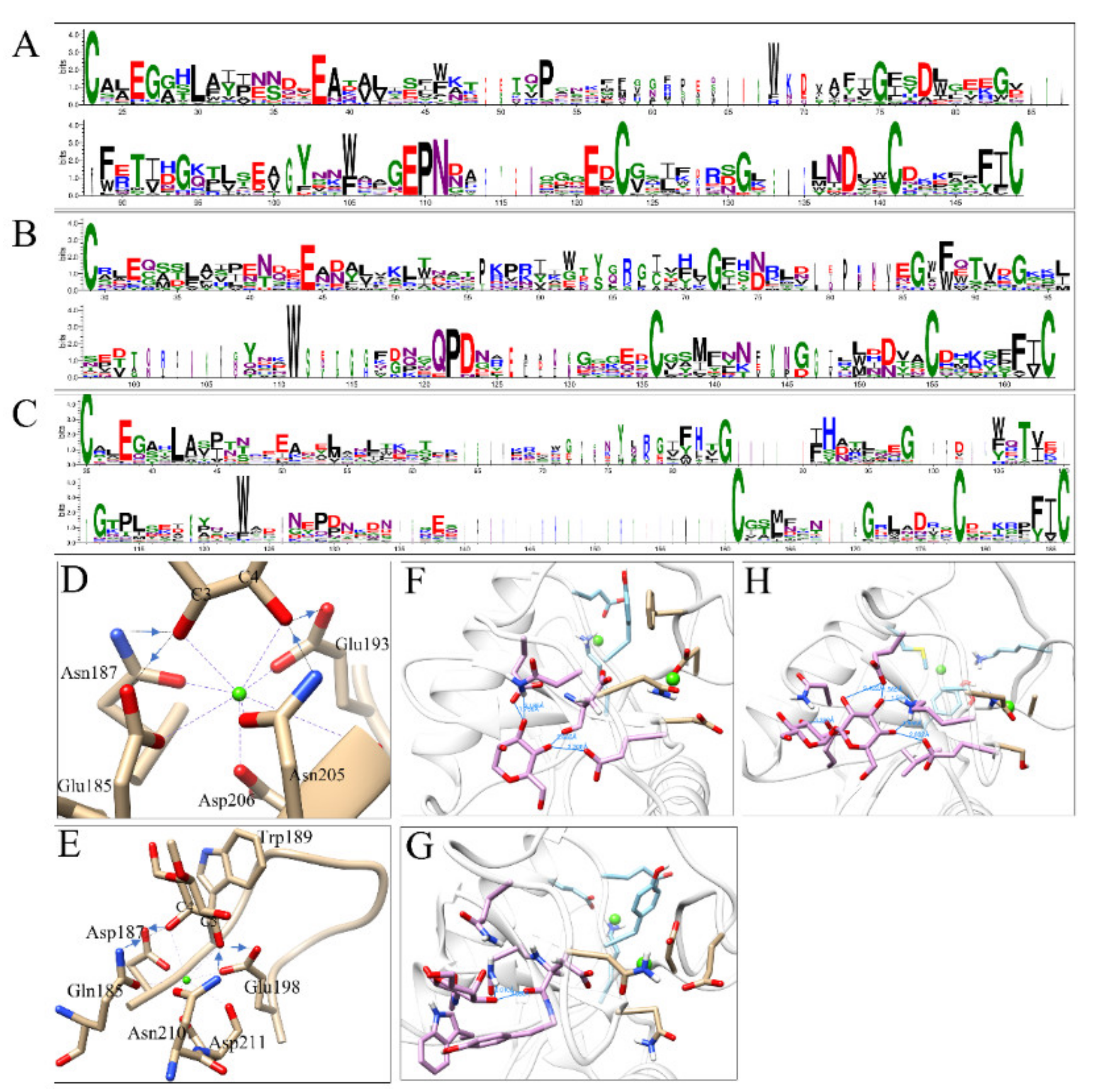
3.3. Sequence and Structure Analyses of CRDs
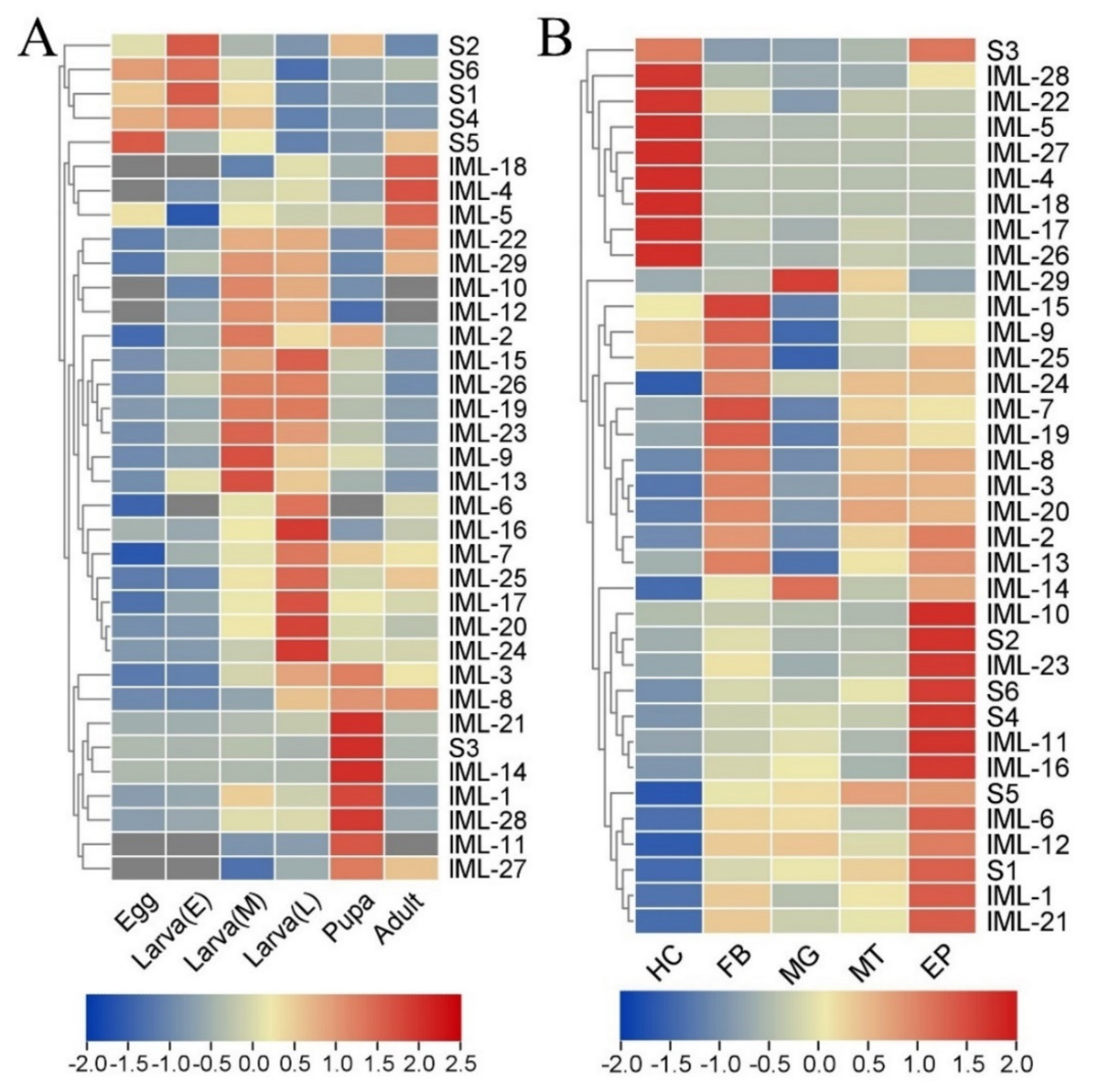
3.4. Spatial and Temporal Expression Profiles
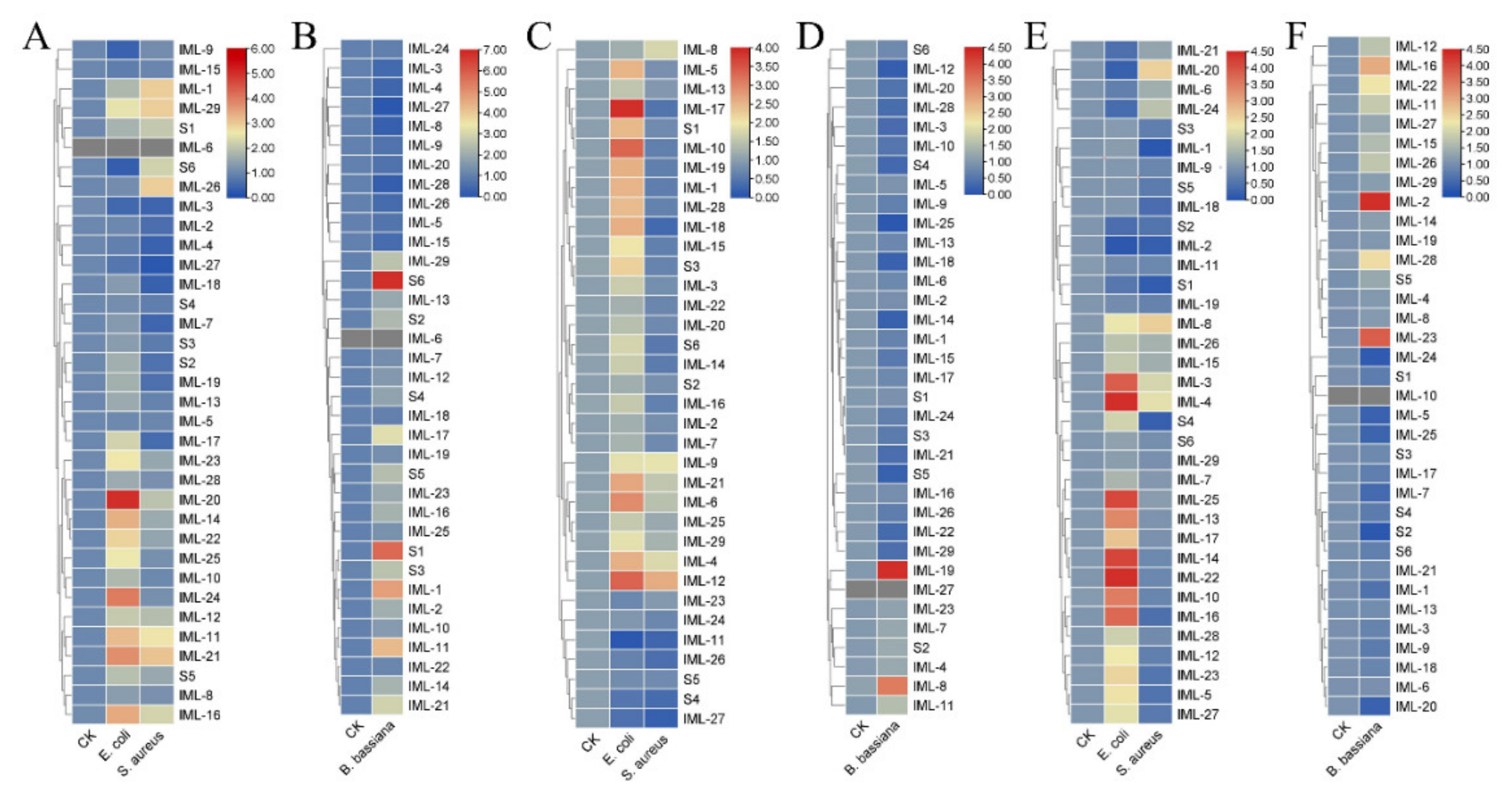
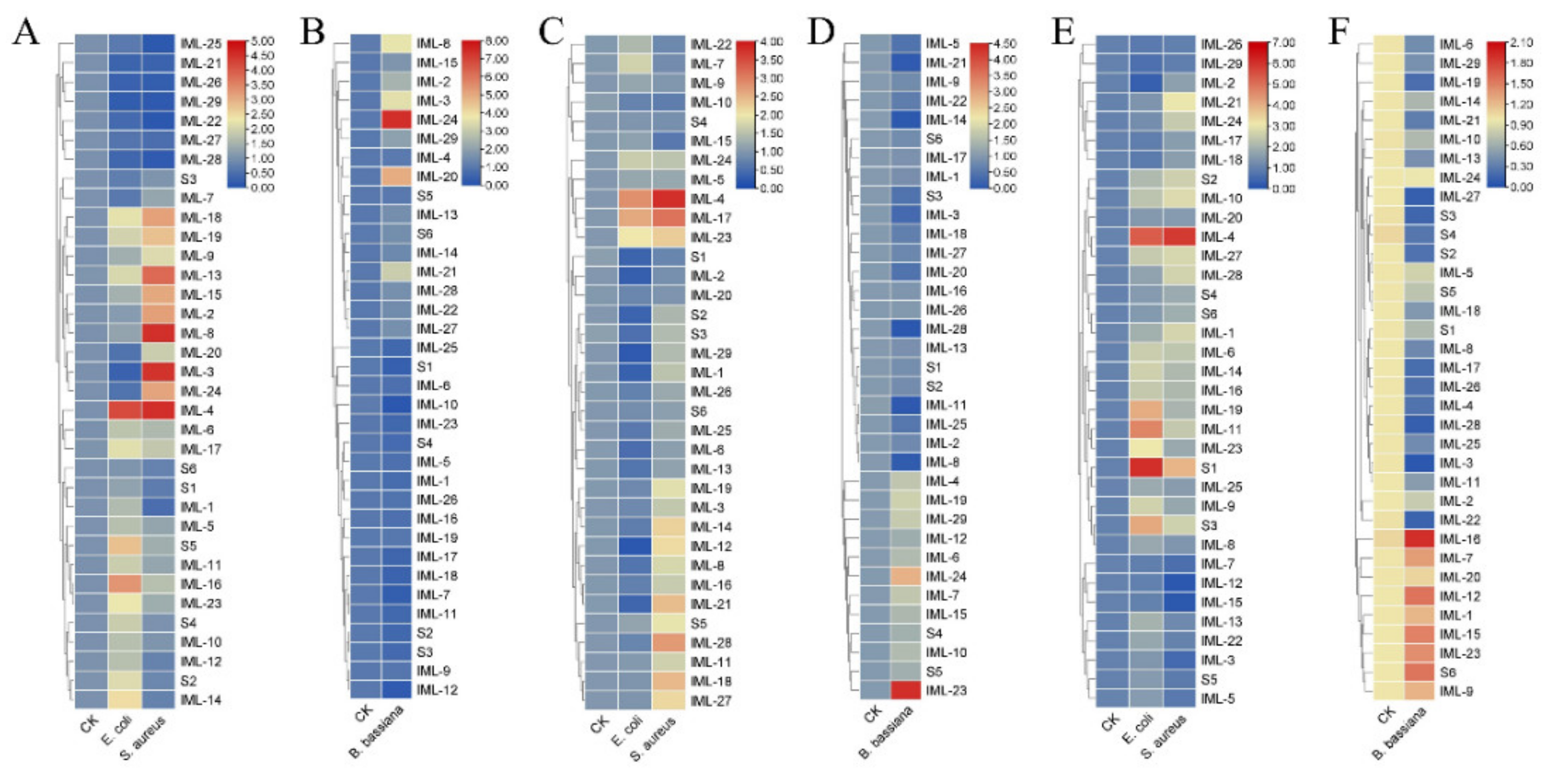
3.5. Expression of CTLs Responding to Bacterial Cells and Fungal Spores
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, J.L.; Cheng, Y.; Wang, J.X.; Zou, Z. Pattern recognition receptors from lepidopteran insects and their biological functions. Dev. Comp. Immunol. 2020, 108, 103688. [Google Scholar] [CrossRef] [PubMed]
- Imler, J.L. Overview of Drosophila immunity: A historical perspective. Dev. Comp. Immunol. 2014, 42, 3–15. [Google Scholar] [CrossRef]
- Rao, X.J.; Zhan, M.Y.; Pan, Y.M.; Liu, S.; Yang, P.J.; Yang, L.L.; Yu, X.Q. Immune functions of insect βGRPs and their potential application. Dev. Comp. Immunol. 2018, 83, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; You, M.; Rao, X.J.; Yu, X.Q. Insect C-type lectins in innate immunity. Dev. Comp. Immunol. 2018, 83, 70–79. [Google Scholar] [CrossRef]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef]
- Rao, X.J.; Shahzad, T.; Liu, S.; Wu, P.; He, Y.T.; Sun, W.J.; Fan, X.Y.; Yang, Y.F.; Shi, Q.; Yu, X.Q. Identification of C-type lectin-domain proteins (CTLDPs) in silkworm Bombyx mori. Dev. Comp. Immunol. 2015, 53, 328–338. [Google Scholar] [CrossRef]
- Rao, X.J.; Cao, X.; He, Y.; Hu, Y.; Zhang, X.; Chen, Y.R.; Blissard, G.; Kanost, M.R.; Yu, X.Q.; Jiang, H. Structural features, evolutionary relationships, and transcriptional regulation of C-type lectin-domain proteins in Manduca sexta. Insect Biochem. Mol. Biol. 2015, 62, 75–85. [Google Scholar] [CrossRef]
- Nagae, M.; Yamaguchi, Y. Structural Aspects of Carbohydrate Recognition Mechanisms of C-Type Lectins. In C-Type Lectins in Immune Homeostasis, 2019/11/30 ed.; Yamasaki, S., Ed.; Springer: Cham, Switzerland, 2019; Volume 429, pp. 147–176. [Google Scholar]
- Weis, W.I.; Drickamer, K.; Hendrickson, W.A. Structure of a C-type mannose-binding protein complexed with an oligosaccharide. Nature 1992, 360, 127–134. [Google Scholar] [CrossRef]
- Yu, X.Q.; Gan, H.; Kanost, M.R. Immulectin, an inducible C-type lectin from an insect, Manduca sexta, stimulates activation of plasma prophenol oxidase. Insect Biochem. Mol. Biol. 1999, 29, 585–597. [Google Scholar] [CrossRef]
- Yu, X.Q.; Kanost, M.R. Immulectin-2, a lipopolysaccharide-specific lectin from an insect, Manduca sexta, is induced in response to gram-negative bacteria. J. Biol. Chem. 2000, 275, 37373–37381. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Q.; Ling, E.; Tracy, M.E.; Zhu, Y. Immulectin-4 from the tobacco hornworm Manduca sexta binds to lipopolysaccharide and lipoteichoic acid. Insect Mol. Biol. 2006, 15, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Ling, E.; Yu, X.Q. Drosophila C-type lectins enhance cellular encapsulation. Mol. Immunol. 2007, 44, 2541–2548. [Google Scholar] [CrossRef]
- Bi, J.; Ning, M.; Li, J.; Zhang, P.; Wang, L.; Xu, S.; Zhong, Y.; Wang, Z.; Song, Q.; Li, B. A C-type lectin with dual-CRD from Tribolium castaneum is induced in response to bacterial challenge. Pest Manage. Sci. 2020, 76, 3965–3974. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhang, Q.; Tang, L.; Chen, L.; Liu, X.S.; Wang, Y.F. Involvement of a pattern recognition receptor C-type lectin 7 in enhancing cellular encapsulation and melanization due to its carboxyl-terminal CRD domain in the cotton bollworm, Helicoverpa armigera. Dev. Comp. Immunol. 2014, 44, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Lin, Z.; Wang, J.M.; Xing, L.S.; Xiong, G.H.; Zou, Z. CTL14, a recognition receptor induced in late stage larvae, modulates anti-fungal immunity in cotton bollworm Helicoverpa armigera. Dev. Comp. Immunol. 2018, 84, 142–152. [Google Scholar] [CrossRef]
- Koizumi, N.; Imamura, M.; Kadotani, T.; Yaoi, K.; Iwahana, H.; Sato, R. The lipopolysaccharide-binding protein participating in hemocyte nodule formation in the silkworm Bombyx mori is a novel member of the C-type lectin superfamily with two different tandem carbohydrate-recognition domains. FEBS Lett. 1999, 443, 139–143. [Google Scholar] [CrossRef]
- Watanabe, A.; Miyazawa, S.; Kitami, M.; Tabunoki, H.; Ueda, K.; Sato, R. Characterization of a novel C-type lectin, Bombyx mori multibinding protein, from the B. mori hemolymph: Mechanism of wide-range microorganism recognition and role in immunity. J. Immunol. 2006, 177, 4594–4604. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Wang, F.; Gatehouse, J.A.; Gatehouse, A.M.; Chen, X.X.; Hu, C.; Ye, G.Y. Venom of parasitoid, Pteromalus puparum, suppresses host, Pieris rapae, immune promotion by decreasing host C-type lectin gene expression. PLoS ONE 2011, 6, e26888. [Google Scholar] [CrossRef]
- Cheng, G.; Cox, J.; Wang, P.; Krishnan, M.N.; Dai, J.; Qian, F.; Anderson, J.F.; Fikrig, E. A C-type lectin collaborates with a CD45 phosphatase homolog to facilitate West Nile virus infection of mosquitoes. Cell 2010, 142, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Luo, L.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of migration in Mythimna separata (Walker) in China: A review integrating environmental, physiological, hormonal, genetic, and molecular factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef]
- Wang, C.; Wang, S. Insect Pathogenic Fungi: Genomics, Molecular Interactions, and Genetic Improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Wang, S.B. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.F.; Li, H.; Yang, P.J.; Rao, X.J. Structure-function analysis of PGRP-S1 from the oriental armyworm, Mythimna separata. Arch. Insect Biochem. Physiol. 2021, 106, e21763. [Google Scholar] [CrossRef]
- Liu, F.F.; Ding, C.; Yang, L.L.; Li, H.; Rao, X.J. Identification and analysis of two lebocins in the oriental armyworm Mythimna separata. Dev. Comp. Immunol. 2021, 116, 103962. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.-J.; Zhan, M.-Y.; Yang, L.-L.; Liu, Q.-Q.; Xu, Y.; Pan, Y.-M.; Rao, X.-J. Characterization of PGRP-S1 from the oriental armyworm, Mythimna separata. Dev. Comp. Immunol. 2019, 90, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 3. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Su, F.; Zhu, K.; Zhu, M.; Li, Q.; Hu, Q.; Zhang, J.; Zhang, R.; Yu, X.-Q. Comparative genomic analysis of C-type lectin-domain genes in seven holometabolous insect species. Insect Biochem. Mol. Biol. 2020, 126, 103451. [Google Scholar] [CrossRef]
- Koizumi, N.; Imai, Y.; Morozumi, A.; Imamura, M.; Kadotani, T.; Yaoi, K.; Iwahana, H.; Sato, R. Lipopolysaccharide-binding protein of Bombyx mori participates in a hemocyte-mediated defense reaction against gram-negative bacteria. J. Insect Physiol. 1999, 45, 853–859. [Google Scholar] [CrossRef]
- Stark, R.; Grzelak, M.; Hadfield, J. RNA sequencing: The teenage years. Nat. Rev. Genet. 2019, 20, 631–656. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qi, M.; Chi, Y.; Wuriyanghan, H. De Novo Assembly of the Transcriptome for Oriental Armyworm Mythimna separata (Lepidoptera: Noctuidae) and Analysis on Insecticide Resistance-Related Genes. J. Insect Sci. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.D.; Wang, W.Z.; Wang, Y.R.; Gao, S.J.; Elzaki, M.; Wang, R.; Wang, R.; Wu, M. Response of detoxification and immune genes and of transcriptome expression in Mythimna separata following chlorantraniliprole exposure. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 28, 90–98. [Google Scholar] [CrossRef]
- Duan, Y.; Gong, Z.; Wu, R.; Miao, J.; Jiang, Y.; Li, T.; Wu, X.; Wu, Y. Transcriptome analysis of molecular mechanisms responsible for light-stress response in Mythimna separata (Walker). Sci. Rep. 2017, 7, 45188. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, X.; Lei, C.; Zhu, F. Sensory genes identification with head transcriptome of the migratory armyworm, Mythimna separata. Sci. Rep. 2017, 7, 46033. [Google Scholar] [CrossRef]
- Chang, X.Q.; Nie, X.P.; Zhang, Z.; Zeng, F.F.; Lv, L.; Zhang, S.; Wang, M.Q. De novo analysis of the oriental armyworm Mythimna separata antennal transcriptome and expression patterns of odorant-binding proteins. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 22, 120–130. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Q.; Feng, B.; Guo, Q.S.; Du, Y. Age influences the olfactory profiles of the migratory oriental armyworm mythimna separate at the molecular level. BMC Genom. 2017, 18, 32. [Google Scholar] [CrossRef]
- Du, L.; Zhao, X.; Liang, X.; Gao, X.; Liu, Y.; Wang, G. Identification of candidate chemosensory genes in Mythimna separata by transcriptomic analysis. BMC Genom. 2018, 19, 518. [Google Scholar] [CrossRef]
- Huang, Y.; Yin, H.; Zhu, Z.; Jiang, X.; Li, X.; Dong, Y.; Sheng, C.; Liao, M.; Cao, H. Expression and functional analysis of cytochrome P450 genes in the integument of the oriental armyworm, Mythimna separata (Walker). Pest Manage. Sci. 2021, 77, 577–587. [Google Scholar] [CrossRef]
- Cummings, R.; McEver, R. C-Type Lectins. In Essentials of Glycobiology [Internet], 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017. [Google Scholar]
- Kubo, T.; Kawasaki, K.; Natori, S. Transient appearance and localization of a 26-kDa lectin, a novel member of the Periplaneta lectin family, in regenerating cockroach leg. Dev. Biol. 1993, 156, 381–390. [Google Scholar] [CrossRef]
- Wang, W.; Wang, G.; Zhuo, X.; Liu, Y.; Tang, L.; Liu, X.; Wang, J. C-type lectin-mediated microbial homeostasis is critical for Helicoverpa armigera larval growth and development. PLoS Pathog. 2020, 16, e1008901. [Google Scholar] [CrossRef]
- Kim, J.C.; Lee, M.R.; Kim, S.; Park, S.E.; Lee, S.J.; Shin, T.Y.; Kim, W.J.; Kim, J. Transcriptome Analysis of the Japanese Pine Sawyer Beetle, Monochamus alternatus, Infected with the Entomopathogenic Fungus Metarhizium anisopliae JEF-197. J. Fungi 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Guo, L.; Tu, C.; Wang, A.; Xu, L.; Luo, J. Comparative Analysis of Adelphocoris suturalis Jakovlev (Hemiptera: Miridae) Immune Responses to Fungal and Bacterial Pathogens. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Shen, D.X.; Liu, Y.; Zhou, F.; Wang, G.R.; An, C.J. Identification of Immunity-Related Genes in Ostrinia furnacalis against Entomopathogenic Fungi by RNA-Seq Analysis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Drickamer, K. Engineering galactose-binding activity into a C-type mannose-binding protein. Nature 1992, 360, 183–186. [Google Scholar] [CrossRef]
- Moriuchi, H.; Unno, H.; Goda, S.; Tateno, H.; Hirabayashi, J.; Hatakeyama, T. Mannose-recognition mutant of the galactose/N-acetylgalactosamine-specific C-type lectin CEL-I engineered by site-directed mutagenesis. Biochim. Biophys. Acta 2015, 1850, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Iobst, S.T.; Drickamer, K. Binding of sugar ligands to Ca(2+)-dependent animal lectins. II. Generation of high-affinity galactose binding by site-directed mutagenesis. J. Biol. Chem. 1994, 269, 15512–15519. [Google Scholar] [CrossRef]
- Kolatkar, A.R.; Weis, W.I. Structural basis of galactose recognition by C-type animal lectins. J. Biol. Chem. 1996, 271, 6679–6685. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, T.; Kamiya, T.; Kusunoki, M.; Nakamura-Tsuruta, S.; Hirabayashi, J.; Goda, S.; Unno, H. Galactose recognition by a tetrameric C-type lectin, CEL-IV, containing the EPN carbohydrate recognition motif. J. Biol. Chem. 2011, 286, 10305–10315. [Google Scholar] [CrossRef]
- Poget, S.F.; Legge, G.B.; Proctor, M.R.; Butler, P.J.; Bycroft, M.; Williams, R.L. The structure of a tunicate C-type lectin from Polyandrocarpa misakiensis complexed with D -galactose. J. Mol. Biol. 1999, 290, 867–879. [Google Scholar] [CrossRef]
- Swaminathan, G.J.; Myszka, D.G.; Katsamba, P.S.; Ohnuki, L.E.; Gleich, G.J.; Acharya, K.R. Eosinophil-granule major basic protein, a C-type lectin, binds heparin. Biochemistry 2005, 44, 14152–14158. [Google Scholar] [CrossRef] [PubMed]
- Unno, H.; Itakura, S.; Higuchi, S.; Goda, S.; Yamaguchi, K.; Hatakeyama, T. Novel Ca2+-independent carbohydrate recognition of the C-type lectins, SPL-1 and SPL-2, from the bivalve Saxidomus purpuratus. Protein Sci. 2019, 28, 766–778. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database | Number | Percentage |
|---|---|---|
| NR | 18,157 | 39.57 |
| GO | 8035 | 17.51 |
| KEGG | 8836 | 19.26 |
| Pfam | 9527 | 20.76 |
| eggNOG | 17,125 | 37.32 |
| Swiss-Prot | 11,142 | 24.28 |
| In all database | 3564 | 7.77 |
| Name | Accession Number | ORF (aa) | BLASTX Best Hit | |||
|---|---|---|---|---|---|---|
| Reference Organism | Accession Number | E Value | Identity (%) | |||
| CTL-S1 | MW658722 | 217 | Manduca sexta | XP_030027697.2 | 4 × 10−147 | 99 |
| CTL-S2 | MW658723 | 306 | Spodoptera litura | XP_022830124.1 | 2 × 10−179 | 86 |
| CTL-S3 | MW658724 | 207 | Helicoverpa armigera | XP_021189783.1 | 5 × 10−143 | 100 |
| CTL-S4 | MW658725 | 221 | Manduca sexta | XP_030021432.1 | 1 × 10−149 | 95 |
| CTL-S5 | MW658726 | 387 | Spodoptera litura | XP_022815888.1 | 0 | 89 |
| CTL-S6 | MW658727 | 223 | Spodoptera litura | XP_022821058.1 | 3 × 10−152 | 99 |
| IML-1 | MW658728 | 339 | Spodoptera frugiperda | XP_035436562.1 | 6 × 10−131 | 58 |
| IML-2 | MW658729 | 321 | Spodoptera frugiperda | XP_035436562.1 | 0 | 77 |
| IML-3 | MW658730 | 325 | Spodoptera exigua | KAF9421847.1 | 6 × 10−122 | 54 |
| IML-4 | MW658731 | 330 | Mythimna separata | BBC20960.1 | 2 × 10−165 | 71 |
| IML-5 | MW658732 | 277 | Spodoptera exigua | AQX37248.1 | 6 × 10−106 | 54 |
| IML-6 | MW658733 | 291 | Mythimna separata | BBC20960.1 | 3 × 10−118 | 62 |
| IML-7 | MW658734 | 281 | Spodoptera frugiperda | XP_035436562.1 | 2 × 10−104 | 53 |
| IML-8 | MW658735 | 307 | Trichoplusia ni | XP_026724737.1 | 9 × 10−150 | 65 |
| IML-9 | MW658736 | 340 | Spodoptera frugiperda | XP_035436562.1 | 3 × 10−110 | 51 |
| IML-10 | MW658737 | 331 | Spodoptera exigua | AQX37248.1 | 2 × 10−110 | 51 |
| IML-11 | MW658738 | 327 | Spodoptera frugiperda | XP_035436562.1 | 1 × 10−112 | 50 |
| IML-12 | MW658739 | 334 | Spodoptera frugiperda | XP_035436562.1 | 8 × 10−111 | 54 |
| IML-13 | MW658740 | 318 | Spodoptera frugiperda | XP_035436562.1 | 4 × 10−114 | 54 |
| IML-14 | MW658741 | 320 | Spodoptera frugiperda | XP_035436562.1 | 2 × 10−121 | 57 |
| IML-15 | MW658742 | 316 | Spodoptera frugiperda | XP_035436562.1 | 3 × 10−119 | 56 |
| IML-16 | MW658743 | 330 | Spodoptera frugiperda | XP_035436562.1 | 3 × 10−116 | 52 |
| IML-17 | MW658744 | 320 | Mythimna separata | BAV01248.1 | 0 | 98 |
| IML-18 | MW658745 | 325 | Mythimna separata | BBC20960.1 | 1 × 10−146 | 63 |
| IML-19 | MW658746 | 320 | Mythimna separata | BAV01248.1 | 9 × 10−127 | 59 |
| IML-20 | MW658747 | 317 | Spodoptera frugiperda | XP_035436562.1 | 4 × 10−121 | 56 |
| IML-21 | MW658748 | 329 | Spodoptera frugiperda | XP_035436562.1 | 1 × 10−120 | 54 |
| IML-22 | MW658749 | 326 | Trichoplusia ni | XP_026732761.1 | 1 × 10−112 | 53 |
| IML-23 | MW658750 | 317 | Mythimna separata | BAV01248.1 | 2 × 10−120 | 57 |
| IML-24 | MW658751 | 326 | Spodoptera frugiperda | XP_035436562.1 | 8 × 10−126 | 55 |
| IML-25 | MW658752 | 330 | Spodoptera frugiperda | XP_035436562.1 | 1 × 10−119 | 59 |
| IML-26 | MW658753 | 289 | Mythimna separata | BBC20960.1 | 3 × 10−12 | 62 |
| IML-27 | MW658754 | 324 | Mythimna separata | BBC20960.1 | 0 | 99 |
| IML-28 | MW658755 | 326 | Mythimna separata | BBC20960.1 | 2 × 10−135 | 59 |
| IML-29 | MW658756 | 312 | Trichoplusia ni | XP_026732761.1 | 1 × 10−142 | 64 |
| CRD | Motif | PDB Hit | Ca2+ Coordinators 2 | Ligand 1 | Ligand Binding Residues 2 | Disulfide Bridges |
|---|---|---|---|---|---|---|
| CTL-S1 | QPD | 1dv8A | 38,40,44,158 [0.14] | GQ2 | 59,115,117,118,132,145,146,147,153 [0.48] | 70–198; 171–190 |
| CTL-S2 | EPN | 4kzvA | 62,66,90,99,100 [0.39] | MAN | 87,89,99,115,116,118 [0.87] | 197–301; 275–293 |
| CTL-S3 | VPQ | 1htnA | 36,38,42,160 [0.13] | 1SL5A | 104,105,106,107,108,109,110,111,116,117,119,120,121,122,123,124,125,126,148,153 [0.24] | 55–187; 164–179 |
| CTL-S4 | QPD | 4kzvA | 35,37,41,154 [0.13] | 4RS | 56,112,114,124,130,141,142,147 [0.51] | 75–202; 175–194 |
| CTL-S5 | - | 1wmyA | 31,33,37,121 [0.12] | GQ2 | 53,85,87,88,98,108,109,110,116 [0.35] | 69–167; 143–159 |
| CTL-S6 | QPD | 4kzvA | 38,40,44,163 [0.10] | BM3 | 120,122,124,133,150,151,152 [0.50] | 71–204; 177–196 |
| IML-1-CRD1 | QPD | 4kzvA | 56,60,85,90,91 [0.44] | NGA | 49,82,84,86,90,94,96,105,106 [0.57] | 57–157; 132–149 |
| IML-1-CRD2 | QPD | 4yliA | 31,33,37,126 [0.22] | 291 | 93,95,101,107,113,114,115 [0.32] | 203–306; 284–298 |
| IML-2-CRD1 | EPN | 4yliA | 55,59,62,84,89,90 [0.27] | MAN | 81,83,89,101,102,104 [0.73] | 58–154; 132–146 |
| IML-2-CRD2 | QPD | 1wmyA | 31,33,37,126 [0.13] | 4RS | 61,94,96,101,107,113,114,119 [0.66] | 200–303; 281–295 |
| IML-3-CRD1 | QPD | 4yliA | 56,60,85,90,91 [0.38] | GQ2 | 49,82,84,86,94,102,103,104,110 [0.58] | 58–155; 133–147 |
| IML-3-CRD2 | QPE | 4yliA | 31,33,37,125 [0.16] | 4RS | 61,93,95,100,106,112,113,118 [0.51] | 202–304; 282–296 |
| IML-4-CRD1 | EPD | 4yliA | 24,26,30,114 [0.03] | MAN | 81,83,89,101,102,104 [0.81] | 51–149; 127–141 |
| IML-4-CRD2 | EPN | 4yliA | 31,33,37,131 [0.11] | 4RS | 64,98,100,106,112,118,119,124 [0.64] | 195–303; 281–295 |
| IML-5-CRD1 | QPD | 4yliA | 56,60,63,85,90,91 [0.10] | TRE | 49,82,84,86,90,96,102,103,108 [0.67] | 17–114; 92–106 |
| IML-5-CRD2 | KPA | 4yliA | 29,31,35,123 [0.23] | BM3 | 92,94,96,98,110,111,112 [0.21] | 160–262; 240–254 |
| IML-6-CRD1 | EPD | 4yliA | 37,41,44,66,71,72 [0.15] | NAG | 63,65,71,83,84,89 [0.79] | 19–114; 91–105 |
| IML-6-CRD2 | EPN | 4yliA | 31,33,37,131 [0.11] | MAN | 61,98,100,106,118,119,121 [0.67] | 158–266; 244–258 |
| IML-7-CRD1 | QPS | 4yliA | 61,65,90,95,96 [0.38] | MAN | 87,89,95,107,108,110 [0.65] | 58–155; 133–147 |
| IML-7-CRD2 | LPQ | 4yliA | 66,70,72,95,98,99 [0.15] | MAN | 92,94,96,98 [0.39] | 201-?; 281-? |
| IML-8-CRD1 | NTD | 1htnA | 40,42,46,122 [0.16] | NGA | 63,89,90,91,92,109,110,111 [0.23] | 57–147; 125–139 |
| IML-8-CRD2 | EPN | 1jznA | 27,29,33,127 [0.10] | MAN | 54,93,95,101,114,115,117 [0.72] | 193–301; 278–293 |
| IML-9-CRD1 | HPD | 4yliA | 31,33,37,117 [0.16] | 4RS | 54,87,89,92,98,104,105,110 [0.45] | 58–152; 130–144 |
| IML-9-CRD2 | QAD | 4yliA | 31,33,37,125 [0.22] | GQ2 | 61,92,94,95,104,112,113,114,120 [0.34] | 198–300; 278–292 |
| IML-10-CRD1 | EPN | 4yliA | 56,60,85,90,91 [0.22] | MAN | 82,84,90,103,104,106 [0.71] | 57–155; 132–147 |
| IML-10-CRD2 | QPD | 4yliA | 29,31,35,125 [0.20] | NGA | 59,92,94,96,100,104,106,112,113 [0.20] | 201–305; 283–297 |
| IML-11-CRD1 | TPD | 4yliA | 56,60,85,90,91 [0.24] | MAN | 82,84,90,102,103,105 [0.62] | 57–154; 132–146 |
| IML-11-CRD2 | KPD | 4yliA | 31,33,37,124 [0.14] | GAL | 94,96,99,111,112 [0.21] | 200–301; 279–293 |
| IML-12-CRD1 | TPD | 4kzvA | 56,60,85,90,91 [0.27] | NGA | 49,82,84,86,90,94,96,105,106 [0.55] | 57–157; 132–149 |
| IML-12-CRD2 | EPD | 4yliA | 31,33,37,126 [0.19] | GQ2 | 61,93,95,96,105,113,114,115,121 [0.52] | 203–306; 284–298 |
| IML-13-CRD1 | EPN | 4yliA | 57,61,86,90,91 [0.23] | MAN | 83,85,90,102,103,105 [0.69] | 59–155; 133–147 |
| IML-13-CRD2 | QPD | 4yliA | 31,33,37,125 [0.21] | 4RS | 61,93,95,100,106,112,113,118 [0.47] | 201–303; 281–295 |
| IML-14-CRD1 | QPD | 4yliA | 56,60,63,86,91,92 [0.35] | MAN | 83,85,91,103,104,106 [0.67] | 57–155; 133–147 |
| IML-14-CRD2 | QPD | 4yliA | 31,33,37,127 [0.32] | GQ2 | 61,94,96,97,106,114,115,116,122 [0.09] | 201–305; 283–297 |
| IML-15-CRD1 | EPN | 4yliA | 56,60,63,85,89,90 [0.29] | MAN | 82,84,89,101,102,104 [0.70] | 58–154; 132–146 |
| IML-15-CRD2 | SRL | 4yliA | 31,33,37,124 [0.24] | GQ2 | 61,91,93,94,103,111,112,113,119 [0.30] | 200–301; 279–293 |
| IML-16-CRD1 | EPN | 4yliA | 56,60,85,90,91 [0.23] | MAN | 82,84,90,102,103,105 [0.70] | 57–154; 132–146 |
| IML-16-CRD2 | QPV | 4yliA | 31,33,37,124 [0.13] | MMA | 94,96,98,99,111,112,113 [0.18] | 200–301; 279–293 |
| IML-17-CRD1 | EPD | 4yliA | 18,20,24,109 [0.05] | MAN | 76,78,84,96,97,99 [0.79] | 53–152; 130–144 |
| IML-17-CRD2 | EPN | 4yliA | 31,33,37,128 [0.11] | TRE | 61,95,97,99,103,109,115,116,121 [0.63] | 198–303; 281–295 |
| IML-18-CRD1 | EPD | 4yliA | 18,20,24,106 [0.04] | MAN | 73,75,81,93,94,96 [0.78] | 51–147; 125–139 |
| IML-18-CRD2 | EPT | 4yliA | - | 4RS | 63,97,99,104,110,116,117,122 [0.68] | 193–299, 277–291 |
| IML-19-CRD1 | EPD | 4yliA | 26,28,32,117 [0.04] | TRE | 51,84,86,88,92,98,104,105,110 [0.79] | 57–156; 134–148 |
| IML-19-CRD2 | NPD | 1b08A | 31,33,37,127 [0.18] | GQ2 | 61,95,97,98,106,114,115,116,122 [0.56] | 202–306; 284–298 |
| IML-20-CRD1 | QPN | 4yliA | 26,28,32,113 [0.07] | MAN | 82,84,89,100,101,103 [0.66] | 58–153; 132–145 |
| IML-20-CRD2 | QPI | 4yliA | 31,33,37,126 [0.15] | NGA | 94,96,98,101,113,114,115 [0.41] | 198–301; 279–293 |
| IML-21-CRD1 | IPD | 4yliA | 56,60,86,91,92 [0.23] | TRE | 49,83,85,87,91,97,103,104,109 [0.62] | 57–155; 133–147 |
| IML-21-CRD2 | QPV | 1wmyA | 31,33,37,124 [0.21] | NAG | 91,93,100,111,112,113 [0.35] | 201–302; 280–294 |
| IML-22-CRD1 | EPD | 4yliA | 26,28,32,114 [0.04] | TRE | 48,81,83,85,89,95,101,102,107 [0.81] | 51–147; 125–139 |
| IML-22-CRD2 | KPT | 4yliA | 31,33,37,133 [0.17] | GQ2 | 67,101,103,104,112,120,121,122,128 [0.54] | 193–303; 281–295 |
| IML-23-CRD1 | EPD | 4yliA | 26,28,32,114 [0.04] | MAN | 81,83,89,101,102,104 [0.81] | 51–147; 125–139 |
| IML-23-CRD2 | DPN | 4yliA | 31,33,37,129 [0.10] | 4RS | 61,95,97,104,110,116,117,122 [0.64] | 192–298; 276–290 |
| IML-24-CRD1 | EPN | 4yliA | 56,60,85,90,91 [0.36] | MAN | 82,84,90,102,103,105 [0.68] | 58–155; 133–147 |
| IML-24-CRD2 | VPT | 4yliA | 31,33,37,126 [0.13] | GQ2 | 61,94,96,97,105,113,114,115,121 [0.27] | 201–304; 282–296 |
| IML-25-CRD1 | EPN | 4yliA | 57,61,64,86,91,92 [0.16] | MAN | 83,85,91,103,104,106 [0.69] | 58–156; 134–148 |
| IML-25-CRD2 | QPG | 4yliA | 29,31,35,124 [0.19] | GQ2 | 59,92,94,95,103,111,112,113,119 [0.31] | 202–305; 283–297 |
| IML-26-CRD1 | EPD | 4kzvA | 34,36,40,122 [0.04] | NGA | 56,89,91,93,97,101,103,109,110 [0.78] | 51–147; 125–139 |
| IML-26-CRD2 | EPT | 3zhgA | - | MMA | 97,99,101,104 [0.23] | 194-?; 278-? |
| IML-27-CRD1 | EPD | 4yliA | 24,26,30,113 [0.04] | MAN | 80,82,88,100,101,103 [0.80] | 51–148; 126–140 |
| IML-27-CRD2 | EPN | 4yliA | 31,33,37,130 [0.14] | FUC | 98,100,102,105,117,118,119 [0.32] | 194–301; 279–293 |
| IML-28-CRD1 | EPD | 4yliA | 24,26,30,112 [0.04] | MAN | 79,81,87,99,100,102 [0.81] | 51–147; 125–139 |
| IML-28-CRD2 | EPN | 4yliA | 31,33,37,133 [0.14] | 4RS | 67,101,103,108,114,120,121,126 [0.59] | 192–302; 280–294 |
| IML-29-CRD1 | EPD | 4yliA | 26,28,32,114 [0.04] | NGA | 48,81,83,85,89,93,95,101,102 [0.80] | 51–147; 125–139 |
| IML-29-CRD2 | EPN | 4yliA | 68,72,75,98,103,104 [0.03] | TRE | 61,95,97,99,103,109,115,116,121 [0.65] | 193–298; 276–290 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Liu, F.-F.; Fu, L.-Q.; Liu, Z.; Zhang, W.-T.; Wang, Q.; Rao, X.-J. Identification of 35 C-Type Lectins in the Oriental Armyworm, Mythimna separata (Walker). Insects 2021, 12, 559. https://doi.org/10.3390/insects12060559
Li H, Liu F-F, Fu L-Q, Liu Z, Zhang W-T, Wang Q, Rao X-J. Identification of 35 C-Type Lectins in the Oriental Armyworm, Mythimna separata (Walker). Insects. 2021; 12(6):559. https://doi.org/10.3390/insects12060559
Chicago/Turabian StyleLi, Hao, Fang-Fang Liu, Li-Qing Fu, Ze Liu, Wen-Ting Zhang, Qian Wang, and Xiang-Jun Rao. 2021. "Identification of 35 C-Type Lectins in the Oriental Armyworm, Mythimna separata (Walker)" Insects 12, no. 6: 559. https://doi.org/10.3390/insects12060559
APA StyleLi, H., Liu, F.-F., Fu, L.-Q., Liu, Z., Zhang, W.-T., Wang, Q., & Rao, X.-J. (2021). Identification of 35 C-Type Lectins in the Oriental Armyworm, Mythimna separata (Walker). Insects, 12(6), 559. https://doi.org/10.3390/insects12060559