
Climate Mismatch between Introduced Biological Control Agents and Their Invasive Host Plants: Improving Biological Control of Tropical Weeds in Temperate Regions

 ,
,  , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Why Does Climate Mismatch Occur?
2.1. Inadequate Exploration
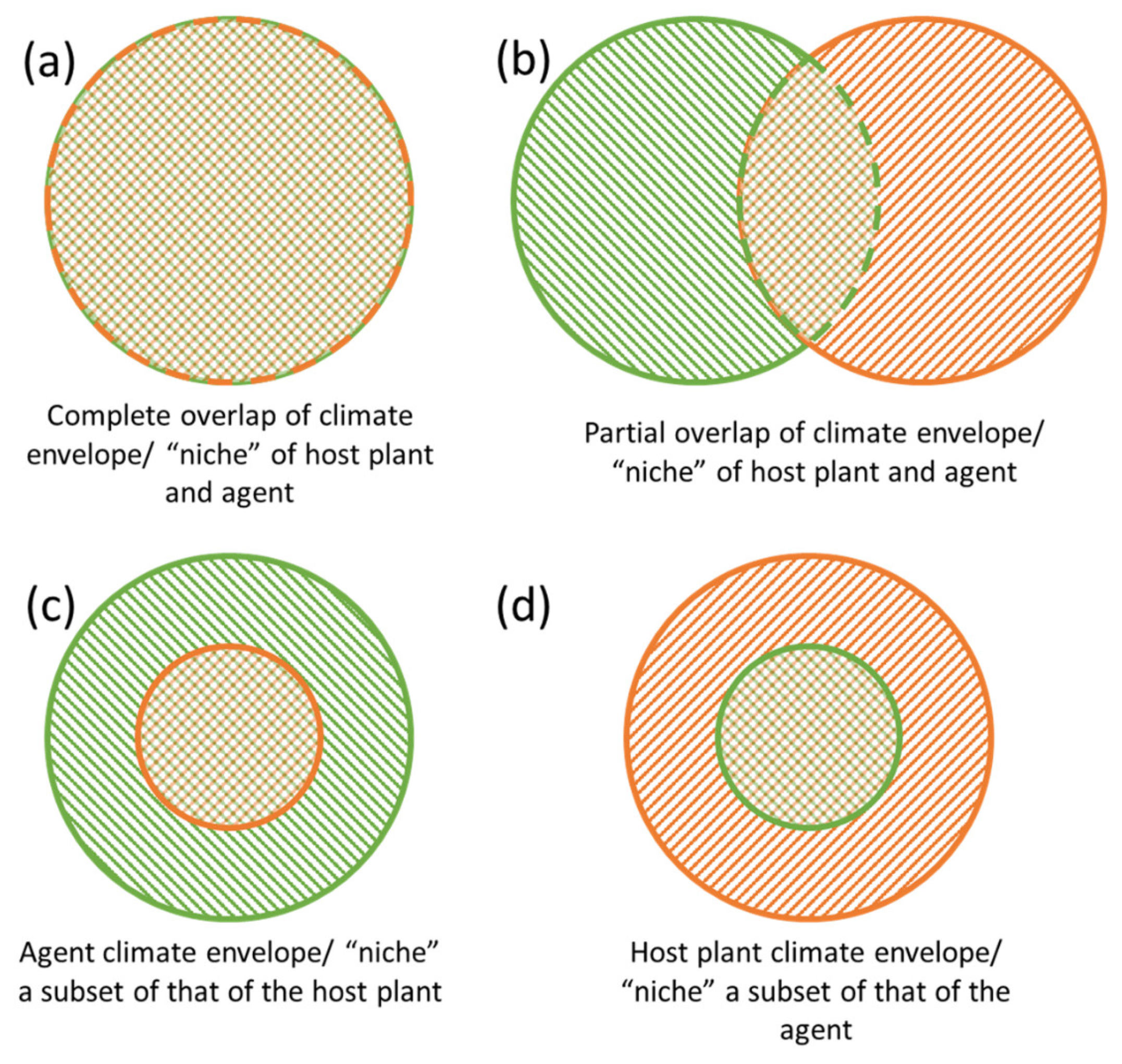
2.2. Climate Niche Shifts and Expansions
2.3. Genetic Bottlenecks, Post-Introduction Evolution, and Hybrids
3. Addressing Climatic Mismatches
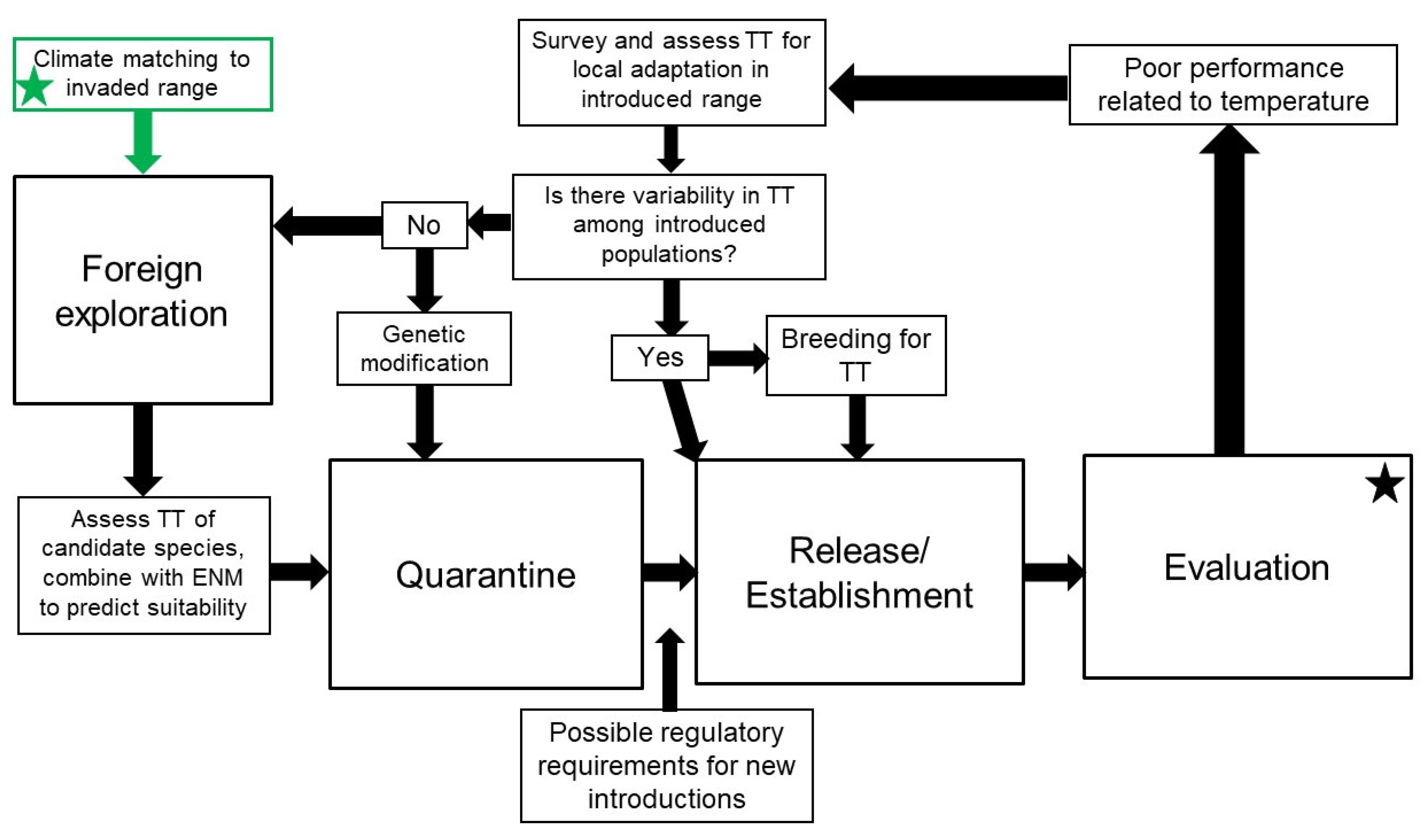
3.1. Traditional Approaches
3.2. Non-Traditional Approaches
4. Cold Tolerance Metrics and Their Application
4.1. Measures of Failure
4.2. Measures of Performance
5. Predicting Distributions of Agents and Hosts
5.1. Environmental Niche Modelling (ENM)
5.2. Mechanistic Modelling
5.3. Fitted Process-Based Models
6. Non-Classical Biological Control: Molecular Approaches to Addressing Climate Mismatch
6.1. Molecular Mechanisms Underlying Cold Tolerance
6.2. Enhancing Cold Hardiness
7. Regulatory Considerations
8. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wainger, L.A.; Harms, N.E.; Magen, C.; Liang, D.; Nesslage, G.M.; McMurray, A.M.; Cofrancesco, A.F. Evidence-based economic analysis demonstrates that ecosystem service benefits of water hyacinth management greatly exceed research and control costs. PeerJ 2018, 6, e4824. [Google Scholar] [CrossRef]
- Harris, P. Cost of biological control of weeds by insects in Canada. Weed Sci. 1979, 27, 242–250. [Google Scholar] [CrossRef]
- Culliney, T.W. Benefits of classical biological control for managing invasive plants. Crit. Rev. Plant Sci. 2005, 24, 131–150. [Google Scholar] [CrossRef]
- Page, A.; Lacey, K. Economic Impact Assessment of Australian Weed Biological Control; CRC for Australian Weed Management: Adelaide, Australia, 2006. [Google Scholar]
- Hinz, H.L.; Winston, R.L.; Schwarzländer, M. How safe is weed biological control? A global review of direct nontarget attack. Q. Rev. Biol. 2019, 94, 1–27. [Google Scholar] [CrossRef]
- Schwarzländer, M.; Hinz, H.L.; Winston, R.L.; Day, M.D. Biological control of weeds: An analysis of introductions, rates of establishment and estimates of success, worldwide. BioControl 2018, 63, 319–331. [Google Scholar] [CrossRef]
- Winston, R.; Schwarzlander, M.; Hinz, H.; Day, M.; Cock, M.J.; Julien, M. Biological Control of Weeds: A World Catalogue of Agents and Their Target Weeds; Forest Health Technology Enterprise Team, USDA Forest Service: Morgantown, WV, USA, 2014.
- Harms, N.E.; Cronin, J.T.; Diaz, R.; Winston, R.L. A review of the causes and consequences of geographical variability in weed biological control successes. Biol. Control 2020, 151, 104398. [Google Scholar] [CrossRef]
- Mukherjee, A.; Knutson, A.; Hahn, D.A.; Heinz, K.M. Biological control of giant salvinia (Salvinia molesta) in a temperate region: Cold tolerance and low temperature oviposition of Cyrtobagous salviniae. BioControl 2014, 59, 781–790. [Google Scholar] [CrossRef]
- Reddy, A.M.; Pratt, P.D.; Hopper, J.V.; Cibils-Stewart, X.; Walsh, G.C.; Mc Kay, F. Variation in cool temperature performance between populations of Neochetina eichhorniae (Coleoptera: Curculionidae) and implications for the biological control of water hyacinth, Eichhornia crassipes, in a temperate climate. Biol. Control 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Julien, M.H.; Skarratt, B.; Maywald, G. Potential geographical distribution of alligator weed and its biological control by Agasicles hygrophila. J. Aquat. Plant Manag. 1995, 33, 55–60. [Google Scholar]
- Howard, G.W.; Harley, K.L.S. How do floating aquatic weeds affect wetland conservation and development? How can these effects be minimised? Wetl. Ecol. Manag. 1997, 5, 215–225. [Google Scholar] [CrossRef]
- Andrewartha, H.G.; Birch, L.C. The Distribution and Abundance of Animals, 1st ed.; University of Chicago Press: Chicago, IL, USA, 1954. [Google Scholar]
- Sexton, J.P.; McIntyre, P.J.; Angert, A.L.; Rice, K.J. Evolution and ecology of species range limits. Annu. Rev. Ecol. Evol. S 2009, 40, 415–436. [Google Scholar] [CrossRef]
- Buckingham, G.R.; Boucias, D. Release of Potentially Cold Tolerant Alligatorweed Flea Beetles (Agasicles hygrophila Selman and Vogt) into the United States from Argentina; Environmental Laboratory, Waterways Experiment Station, Corps of Engineers: Vicksburg, MS, USA, 1982. [Google Scholar]
- Trethowan, P.D.; Robertson, M.P.; McConnachie, A.J. Ecological niche modelling of an invasive alien plant and its potential biological control agents. S. Afr. J. Bot. 2011, 77, 137–146. [Google Scholar] [CrossRef]
- Hopper, K.; Roush, R.T.; Powell, W. Management of genetics of biological-control introductions. Annu. Rev. Entomol. 1993, 38, 27–51. [Google Scholar] [CrossRef]
- Zwölfer, H.; Ghani, M.; Rao, V. Foreign exploration and importation of natural enemies. In Theory and Practice of Biological Control; Academic Press: New York, NY, USA, 1976; pp. 189–207. [Google Scholar]
- DeBach, P.; Rosen, D. Biological Control by Natural Enemies, 2nd ed.; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Maddox, D.; Andres, L.; Hennessey, R.; Blackburn, R.; Spencer, N. Insects to control alligatorweed: An invader of aquatic ecosystems in the United States. Bioscience 1971, 21, 985–991. [Google Scholar] [CrossRef]
- Buckingham, G.R. Biological control of alligatorweed, Alternanthera philoxeroides, the world’s first aquatic weed success story. Castanea 1996, 61, 232–243. [Google Scholar]
- Vogt, G.B.; Quimby, P.C.; Kay, S. Effects of Weather on the Biological Control of Alligatorweed in the Lower Mississippi Valley Region, 1973–1983; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1992; p. 143.
- Zhao, L.L.; Jia, D.; Yuan, X.S.; Guo, Y.Q.; Zhou, W.W.; Ma, R.Y. Cold hardiness of the biological control agent, Agasicles hygrophila, and implications for its potential distribution. Biol. Control 2015, 87, 1–5. [Google Scholar] [CrossRef]
- Harms, N.E.; Cronin, J.T. Biological control agent attack timing and population variability, but not density, best explain target weed density across an environmental gradient. Sci. Rep. 2020, 10, 11062. [Google Scholar] [CrossRef]
- Hough-Goldstein, J.; Lake, E.C.; Shropshire, K.J.; Moore, R.A.; D’Amico, V. Laboratory and field-based temperature-dependent development of a monophagous weevil: Implications for integrated weed management. Biol. Control 2016, 92, 120–127. [Google Scholar] [CrossRef]
- Cowie, B.W.; Venturi, G.; Witkowski, E.T.F.; Byrne, M.J. Does climate constrain the spread of Anthonomus santacruzi, a biological control agent of Solanum mauritianum, in South Africa? Biol. Control 2016, 101, 1–7. [Google Scholar] [CrossRef]
- Mukherjee, A.; Christman, M.C.; Overholt, W.A.; Cuda, J.P. Prioritizing areas in the native range of hygrophila for surveys to collect biological control agents. Biol. Control 2011, 56, 254–262. [Google Scholar] [CrossRef]
- Mc Kay, F.; Logarzo, G.; Natale, E.; Sosa, A.; Walsh, G.C.; Pratt, P.D.; Sodergren, C. Feasibility assessment for the classical biological control of Tamarix in Argentina. BioControl 2018, 63, 169–184. [Google Scholar] [CrossRef]
- Kriticos, D.; Crossman, N.D.; Ota, N.; Scott, J.K. Climate Change and Invasive Plants in South Australia; CSIRO Climate Adaptation Flagship: Canberra, Australia, 2010.
- Scott, J.; Murphy, H.; Kriticos, D.; Webber, B.; Ota, N.; Loechel, B. Weeds and Climate Change: Supporting Weed Management Adaptation; CSIRO: Canberra, Australia, 2014.
- Mukherjee, A.; Diaz, R.; Thom, M.; Overholt, W.; Cuda, J. Niche-based prediction of establishment of biocontrol agents: An example with Gratiana boliviana and tropical soda apple. Biocontrol Sci. Technol. 2012, 22, 447–461. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Pearman, P.B.; Guisan, A.; Broennimann, O.; Randin, C.F. Niche dynamics in space and time. Trends Ecol. Evol. 2008, 23, 149–158. [Google Scholar] [CrossRef]
- Sax, D.F.; Stachowicz, J.J.; Brown, J.H.; Bruno, J.F.; Dawson, M.N.; Gaines, S.D.; Grosberg, R.K.; Hastings, A.; Holt, R.D.; Mayfield, M.M.; et al. Ecological and evolutionary insights from species invasions. Trends Ecol. Evol. 2007, 22, 465–471. [Google Scholar] [CrossRef]
- Wiens, J.J. The niche, biogeography and species interactions. Philos. T. Roy. Soc. B 2011, 366, 2336–2350. [Google Scholar] [CrossRef]
- Tingley, R.; Vallinoto, M.; Sequeira, F.; Kearney, M.R. Realized niche shift during a global biological invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 10233–10238. [Google Scholar] [CrossRef]
- Warren, D.L.; Cardillo, M.; Rosauer, D.F.; Bolnick, D.I. Mistaking geography for biology: Inferring processes from species distributions. Trends Ecol. Evol. 2014, 29, 572–580. [Google Scholar] [CrossRef]
- Holt, R.D.; Barfield, M. Trophic interactions and range limits: The diverse roles of predation. Proc. Biol. Sci. 2009, 276, 1435–1442. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Brunel, S. Assessing and managing the current and future pest risk from water hyacinth, (Eichhornia crassipes), an invasive aquatic plant threatening the environment and water security. PLoS ONE 2016, 11, e0120054. [Google Scholar] [CrossRef]
- Messenger, P.; Wilson, F.; Whitten, M. Variation, fitness, and adaptability of natural enemies. In Theory and Practice of Biological Control; Huffaker, C.B., Messenger, P.S., Eds.; Academic Press: New York, NY, USA, 1976; pp. 209–231. [Google Scholar]
- Ward, S.M.; Gaskin, J.F.; Wilson, L.M. Ecological genetics of plant invasion: What do we know? Invasive Plant Sci. Manag. 2008, 1, 98–109. [Google Scholar] [CrossRef]
- Smith, A.L.; Hodkinson, T.R.; Villellas, J.; Catford, J.A.; Csergő, A.M.; Blomberg, S.P.; Crone, E.E.; Ehrlén, J.; Garcia, M.B.; Laine, A.-L.; et al. Global gene flow releases invasive plants from environmental constraints on genetic diversity. Proc. Natl. Acad. Sci. USA 2020, 117, 4218–4227. [Google Scholar] [CrossRef]
- Prentis, P.J.; Wilson, J.R.U.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. Adaptive evolution in invasive species. Trends Plant Sci. 2008, 13, 288–294. [Google Scholar] [CrossRef]
- Taylor, S.J.; Downie, D.A.; Paterson, I.D. Genetic diversity of introduced populations of the water hyacinth biological control agent Eccritotarsus catarinensis (Hemiptera: Miridae). Biol. Control 2011, 58, 330–336. [Google Scholar] [CrossRef]
- McKibben, G.H.; Grodowitz, M.J.; Villavaso, E.J. Comparison of flight ability of native and two laboratory-reared strains of boll weevils (Coleoptera: Curculionidae) on a flight mill. Environ. Entomol. 1988, 17, 852–854. [Google Scholar] [CrossRef]
- Gloria-Soria, A.; Soghigian, J.; Kellner, D.; Powell, J.R. Genetic diversity of laboratory strains and implications for research: The case of Aedes aegypti. PLOS Negl. Trop. Dis. 2019, 13, e0007930. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.V.; Peterson, P.; Barrett, D.P.; Forgie, S.; Gleeson, D.M.; Harman, H.; Houliston, G.J.; Smith, L. Investigating the poor performance of heather beetle, Lochmaea suturalis (Thompson) (Coleoptera: Chrysomelidae), as a weed biocontrol agent in New Zealand: Has genetic bottlenecking resulted in small body size and poor winter survival? Biol. Control 2015, 87, 32–38. [Google Scholar] [CrossRef]
- Hopper, J.V.; McCue, K.F.; Pratt, P.D.; Duchesne, P.; Grosholz, E.D.; Hufbauer, R.A. Into the weeds: Matching importation history to genetic consequences and pathways in two widely used biological control agents. Evol. Appl. 2019, 12, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.; Johnson, S.; Cibils, X.; McKay, F.; Moshman, L.; Madeira, P.; Blair, Z.; Diaz, R. Surveys in Argentina and Uruguay reveal Cyrtobagous salviniae (Coleoptera: Curculionidae) populations adapted to survive temperate climates in southeastern USA. Biol. Control 2017, 107, 41–49. [Google Scholar] [CrossRef]
- Coulson, J.R. Biological Control of Alligatorweed, 1959–1972: A Review and Evaluation; 0082-9811; U.S. Department of Agriculture, Agricultural Research Service: Hyattsville, MD, USA, 1977; p. 98.
- Pratt, P.D.; Moran, P.J.; Pitcairn, M.; Reddy, A.M.; O’Brien, J. Biological control of invasive plants in California’s Delta: Past, present, and future. J. Aquat. Plant Manag. 2021, in press. [Google Scholar]
- Harris, P. The selection of effective agents for the biological control of weeds. Can. Entomol. 1973, 105, 1495–1503. [Google Scholar] [CrossRef]
- Manrique, V.; Lake, E.C.; Smith, M.C.; Diaz, R.; Franko, C.; Pratt, P.D.; Rayamajhi, M.B.; Overholt, W.A. Comparative evaluation of development and reproductive capacity of two biotypes of Lilioceris cheni (Coleoptera: Chrysomelidae), biological control agents of air potato (Dioscorea bulbifera) in Florida. Ann. Entomol. Soc. Am. 2017, 110, 310–316. [Google Scholar] [CrossRef]
- Smith, M.C.; Overholt, W.H.; Lake, E.C.; Diaz, R.; Manrique, V.; Hight, S.; Rohrig, E.; Minteer, C.R.; Wheeler, G.; Rayamajhi, M.; et al. Changes in latitude: Overwintering survival of two Lilioceris cheni (Coleoptera: Chrysomelidae) biotypes in Florida. Biocontrol Sci. Technol. 2018, 28, 293–306. [Google Scholar] [CrossRef]
- Foley, J.R.; Minteer, C.; Tipping, P.W. Differences in seasonal variation between two biotypes of Megamelus scutellaris (Hemiptera: Delphacidae), a biological control agent for Eichhornia crassipes (Pontederiaceae) in Florida. Fla. Entomol. 2016, 99, 569–571. [Google Scholar] [CrossRef]
- Tipping, P.W.; Center, T.D.; Sosa, A.J.; Dray, F.A. Host specificity assessment and potential impact of Megamelus scutellaris (Hemiptera: Delphacidae) on waterhyacinth Eichhornia crassipes (Pontederiales: Pontederiaceae). Biocontrol Sci. Technol. 2011, 21, 75–87. [Google Scholar] [CrossRef]
- Hosokawa, T.; Kikuchi, Y.; Shimada, M.; Fukatsu, T. Obligate symbiont involved in pest status of host insect. Proc. R. Soc. Lond. B 2007, 274, 1979–1984. [Google Scholar] [CrossRef] [PubMed]
- Lommen, S.T.E.; Koops, K.G.; Cornelder, B.A.; de Jong, P.W.; Brakefield, P.M. Genetics and selective breeding of variation in wing truncation in a flightless aphid control agent. Entomol. Exp. Appl. 2019, 167, 636–645. [Google Scholar] [CrossRef]
- Spivak, M.; Reuter, G.S. Resistance to American foulbrood disease by honey bee colonies Apis mellifera bred for hygienic behavior. Apidologie 2001, 32, 555–565. [Google Scholar] [CrossRef]
- Kruitwagen, A.; Beukeboom, L.W.; Wertheim, B. Optimization of native biocontrol agents, with parasitoids of the invasive pest Drosophila suzukii as an example. Evol. Appl. 2018, 11, 1473–1497. [Google Scholar] [CrossRef] [PubMed]
- Lommen, S.T.E.; de Jong, P.W.; Pannebakker, B.A. It is time to bridge the gap between exploring and exploiting: Prospects for utilizing intraspecific genetic variation to optimize arthropods for augmentative pest control—A review. Entomol. Exp. Appl. 2017, 162, 108–123. [Google Scholar] [CrossRef]
- Lirakis, M.; Magalhães, S. Does experimental evolution produce better biological control agents? A critical review of the evidence. Entomol. Exp. Appl. 2019, 167, 584–597. [Google Scholar] [CrossRef]
- Dierks, A.; Baumann, B.; Fischer, K. Response to selection on cold tolerance is constrained by inbreeding. Evolution 2012, 66, 2384–2398. [Google Scholar] [CrossRef]
- Wright, M.G.; Bennett, G.M. Evolution of biological control agents following introduction to new environments. BioControl 2018, 63, 105–116. [Google Scholar] [CrossRef]
- Hufbauer, R.A.; Roderick, G.K. Microevolution in biological control: Mechanisms, patterns, and processes. Biol. Control 2005, 35, 227–239. [Google Scholar] [CrossRef]
- Lapwong, Y.; Dejtaradol, A.; Webb, J.K. Shifts in thermal tolerance of the invasive Asian house gecko (Hemidactylus frenatus) across native and introduced ranges. Biol. Invasions 2021. [Google Scholar] [CrossRef]
- Geerts, A.N.; Vanoverbeke, J.; Vanschoenwinkel, B.; Van Doorslaer, W.; Feuchtmayr, H.; Atkinson, D.; Moss, B.; Davidson, T.A.; Sayer, C.D.; De Meester, L. Rapid evolution of thermal tolerance in the water flea Daphnia. Nat. Clim. Chang. 2015, 5, 665–668. [Google Scholar] [CrossRef]
- Coetzee, J.A.; Byrne, M.J.; Hill, M.P. Predicting the distribution of Eccritotarsus catarinensis, a natural enemy released on water hyacinth in South Africa. Entomol. Exp. Appl. 2007, 125, 237–247. [Google Scholar] [CrossRef]
- Griffith, T.C.; Paterson, I.D.; Owen, C.A.; Coetzee, J.A. Thermal plasticity and microevolution enhance establishment success and persistence of a water hyacinth biological control agent. Entomol. Exp. Appl. 2019, 167, 616–625. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sgrò, C.M. Climate change and evolutionary adaptation. Nature 2011, 470, 479–485. [Google Scholar] [CrossRef] [PubMed]
- McFadyen, R.E.C. Biological control of weeds. Annu. Rev. Entomol. 1998, 43, 369–393. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Alvarado, L.E.C.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef]
- Lee, R., Jr. A primer on insect cold-tolerance In Low Temperature Biology of Insects; Denlinger, D., Lee, R., Jr., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 3–34. [Google Scholar]
- Bale, J.S. Insect cold hardiness: A matter of life and death. Eur. J. Entomol. 1996, 93, 369–382. [Google Scholar]
- Storey, K.B.; Storey, J.M. Molecular biology of freezing tolerance. Compr. Physiol. 2013, 3, 1283–1308. [Google Scholar] [CrossRef]
- Duman, J.G. Antifreeze and ice nucleator proteins in terrestrial arthropods. Annu. Rev. Physiol. 2001, 63, 327–357. [Google Scholar] [CrossRef] [PubMed]
- Renault, D.; Salin, C.; Vannier, G.; Vernon, P. Survival at low temperatures in insects: What is the ecological significance of the supercooling point? CryoLetters 2002, 23, 217–228. [Google Scholar] [PubMed]
- Nedvěd, O.; Lavy, D.; Verhoef, H. Modelling the time–temperature relationship in cold injury and effect of high-temperature interruptions on survival in a chill-sensitive collembolan. Funct. Ecol. 1998, 12, 816–824. [Google Scholar] [CrossRef]
- Byrne, M.; Coetzee, J.; McConnachie, A.; Parasram, W.; Hill, M. Predicting climate compatibility of biological control agents in their region of introduction. In Proceedings of the 11th International Symposium on Biological Control of Weeds, Canberra, Australia, 27 April–2 May 2003; p. 28. [Google Scholar]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess Drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. R. Soc. Lond. B 2000, 267, 739–745. [Google Scholar] [CrossRef]
- Ditrich, T.; Janda, V.; Vaněčková, H.; Doležel, D. Climatic variation of supercooling point in the Linden bug Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Insects 2018, 9, 144. [Google Scholar] [CrossRef]
- Verdú, J.R. Chill tolerance variability within and among populations in the dung beetle Canthon humectus hidalgoensis along an altitudinal gradient in the mexican semiarid high plateau. J. Arid Environ. 2011, 75, 119–124. [Google Scholar] [CrossRef]
- Rogers, D.J.; Terblanche, J.S.; Owen, C.A. Low-temperature physiology of climatically distinct South African populations of the biological control agent Neochetina eichhorniae. Ecol. Entomol. 2020. [Google Scholar] [CrossRef]
- Ungerer, M.J.; Ayres, M.P.; Lombardero, M.J. Climate and the northern distribution limits of Dendroctonus frontalis Zimmermann (Coleoptera: Scolytidae). J. Biogeogr. 1999, 26, 1133–1145. [Google Scholar] [CrossRef]
- Grant, J.I.; Lamp, W.O. Cold tolerance of Megacopta cribraria (Hemiptera: Plataspidae): An invasive pest of soybeans. Environ. Entomol. 2017, 46, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Hanc, Z.; Nedvěd, O. Chill injury at alternating temperatures in Orchesella cincta (Collembola: Entomobryidae) and Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). Eur. J. Entomol. 1999, 96, 165–168. [Google Scholar]
- Zachariassen, K.E. The role of polyols and nucleating agents in cold-hardy beetles. J. Comp. Physiol. 1980, 140, 227–234. [Google Scholar] [CrossRef]
- Andersen, M.K.; Jensen, N.J.S.; Robertson, R.M.; Overgaard, J. Central nervous system shutdown underlies acute cold tolerance in tropical and temperate Drosophila species. J. Exp. Biol. 2018, 221, jeb179598. [Google Scholar] [CrossRef] [PubMed]
- Gibert, P.; Moreteau, B.; Petavy, G.; Karan, D.; David, J.R. Chill-coma tolerance, a major climatic adaptation among Drosophila species. Evolution 2001, 55, 1063–1068. [Google Scholar] [CrossRef]
- Augustinus, B.; Sun, Y.; Beuchat, C.; Schaffner, U.; Müller-Schärer, H. Predicting impact of a biocontrol agent: Integrating distribution modeling with climate-dependent vital rates. Ecol. Appl. 2020, 30, e02003. [Google Scholar] [CrossRef]
- Coulin, C.; de la Vega, G.J.; Chifflet, L.; Calcaterra, L.A.; Schilman, P.E. Linking thermo-tolerances of the highly invasive ant, Wasmannia auropunctata, to its current and potential distribution. Biol. Invasions 2019, 21, 3491–3504. [Google Scholar] [CrossRef]
- Gibert, P.; Huey, R.B. Chill-coma temperature in Drosophila: Effects of developmental temperature, latitude, and phylogeny. Physiol. Biochem. Zool. 2001, 74, 429–434. [Google Scholar] [CrossRef]
- Hazell, S.P.; Bale, J.S. Low temperature thresholds: Are chill coma and CTmin synonymous? J. Insect Physiol. 2011, 57, 1085–1089. [Google Scholar] [CrossRef]
- Terblanche, J.S.; Jaco Klok, C.; Krafsur, E.S.; Chown, S.L. Phenotypic plasticity and geographic varation in thermal tolerance and water loss of the Tsetse Glossina pallidipes (Diptera: Glossinidae): Implications for distribution modelling. Am. J. Trop. Med. Hyg. 2006, 74, 786–794. [Google Scholar] [CrossRef][Green Version]
- Ransberry, V.E.; MacMillan, H.A.; Sinclair, B.J. The relationship between chill-coma onset and recovery at the extremes of the thermal window of Drosophila melanogaster. Physiol. Biochem. Zool. 2011, 84, 553–559. [Google Scholar] [CrossRef]
- Anderson, A.R.; Hoffmann, A.A.; McKechnie, S.W. Response to selection for rapid chill-coma recovery in Drosophila melanogaster: Physiology and life-history traits. Genet. Res. 2005, 85, 15–22. [Google Scholar] [CrossRef]
- Lü, Z.-C.; Wang, Y.-M.; Zhu, S.-G.; Yu, H.; Guo, J.-Y.; Wan, F.-H. Trade-offs between survival, longevity, and reproduction, and variation of survival tolerance in Mediterranean Bemisia tabaci after temperature stress. J. Insect Sci. 2014, 14. [Google Scholar] [CrossRef][Green Version]
- Nyamukondiwa, C.; Terblanche, J.S.; Marshall, K.E.; Sinclair, B.J. Basal cold but not heat tolerance constrains plasticity among Drosophila species (Diptera: Drosophilidae). J. Evol. Biol. 2011, 24, 1927–1938. [Google Scholar] [CrossRef]
- MacMillan, H.A.; Walsh, J.P.; Sinclair, B.J. The effects of selection for cold tolerance on cross-tolerance to other environmental stressors in Drosophila melanogaster. Insect Sci. 2009, 16, 263–276. [Google Scholar] [CrossRef]
- Marshall, K.E.; Sinclair, B.J. Repeated stress exposure results in a survival-reproduction trade-off in Drosophila melanogaster. Proc. R. Soc. Lond. B 2010, 277, 963–969. [Google Scholar] [CrossRef]
- Shiota, H.; Kimura, M.T. Evolutionary trade-offs between thermal tolerance and locomotor and developmental performance in drosophilid flies. Biol. J. Linn. Soc. 2007, 90, 375–380. [Google Scholar] [CrossRef][Green Version]
- Julien, M.; White, G. Biological control of weeds: Theory and practical application. ACIAR Monogr. Ser. 1997, 49, 1–190. [Google Scholar]
- Müller-Schärer, H.; Schaffner, U. Classical biological control: Exploiting enemy escape to manage plant invasions. Biol. Invasions 2008, 10, 859–874. [Google Scholar] [CrossRef]
- Raghu, S.; Walton, C. Understanding the ghost of Cactoblastis past: Historical clarifications on a poster child of classical biological control. BioScience 2007, 57, 699–705. [Google Scholar] [CrossRef]
- Lantschner, M.V.; de la Vega, G.; Corley, J.C. Predicting the distribution of harmful species and their natural enemies in agricultural, livestock and forestry systems: An overview. Int. J. Pest Manag. 2019, 65, 190–206. [Google Scholar] [CrossRef]
- Mukherjee, A.; Banerjee, A.K.; Raghu, S. Biological control of Parkinsonia aculeata: Using species distribution models to refine agent surveys and releases. Biol. Control 2021, 159, 104630. [Google Scholar] [CrossRef]
- Peterson, A.T.; Soberon, J.; Pearson, R.G.; Anderson, R.P.; Martinez-meyer, E.; Nakamura, M.; Araujo, M. Ecological Niches and Geographic Distributions; Princeton Univeristy Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Oliver, T.H.; Gillings, S.; Girardello, M.; Rapacciuolo, G.; Brereton, T.M.; Siriwardena, G.M.; Roy, D.B.; Pywell, R.; Fuller, R.J. Population density but not stability can be predicted from species distribution models. J. Appl. Ecol. 2012, 49, 581–590. [Google Scholar] [CrossRef]
- Thuiller, W.; Münkemüller, T.; Schiffers, K.H.; Georges, D.; Dullinger, S.; Eckhart, V.M.; Edwards Jr, T.C.; Gravel, D.; Kunstler, G.; Merow, C. Does probability of occurrence relate to population dynamics? Ecography 2014, 37, 1155–1166. [Google Scholar] [CrossRef]
- Sun, Y.; Brönnimann, O.; Roderick, G.K.; Poltavsky, A.; Lommen, S.T.; Müller-Schärer, H. Climatic suitability ranking of biological control candidates: A biogeographic approach for ragweed management in Europe. Ecosphere 2017, 8, e01731. [Google Scholar] [CrossRef]
- Sun, Y.; Zhou, Z.; Wang, R.; Müller-Schärer, H. Biological control opportunities of ragweed are predicted to decrease with climate change in East Asia. Biodivers. Sci. 2017, 25, 1285–1294. [Google Scholar]
- Minghetti, E.; Olivera, L.; Montemayor, S.I. Ecological niche modelling of Gargaphia decoris (Heteroptera), a biological control agent of the invasive tree Solanum mauritianum (Solanales: Solanaceae). Pest Manag. Sci. 2020, 76, 1273–1281. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Manrique, V.; Cuda, J.P.; Overholt, W.A.; Diaz, R. Temperature-dependent development and potential distribution of Episimus utilis (Lepidoptera: Tortricidae), a candidate biological control agent of Brazilian peppertree (Sapindales: Anacardiaceae) in Florida. Environ. Entomol. 2008, 37, 862–870. [Google Scholar] [CrossRef][Green Version]
- Korzukhin, M.; Porter, S.; Thompson, L.; Wiley, S. Modeling temperature-dependent range limits for the red imported fire ant (Hymenoptera: Formicidae: Solenopsis invicta) in the United States. Environ. Entomol. 2001, 30, 645–655. [Google Scholar] [CrossRef]
- Damos, P.; Savopoulou-Soultani, M. Temperature-driven models for insect development and vital thermal requirements. Psyche 2012, 2012, 123405. [Google Scholar] [CrossRef]
- McClay, A.S.; Hughes, R.B. Effects of temperature on developmental rate, distribution, and establishment of Calophasia lunula (Lepidoptera, Noctuidae), a biocontrol agent for Toadflax (Linaria spp). Biol. Control 1995, 5, 368–377. [Google Scholar] [CrossRef]
- Dhileepan, K.; Taylor, D.B.; McCarthy, J.; King, A.; Shabbir, A. Development of cat’s claw creeper leaf-tying moth Hypocosmia pyrochroma (Lepidoptera: Pyralidae) at different temperatures: Implications for establishment as a biological control agent in Australia and South Africa. Biol. Control 2013, 67, 194–202. [Google Scholar] [CrossRef]
- Uyi, O.O.; Zachariades, C.; Hill, M.P.; McConnachie, A.J. Temperature-dependent performance and potential distribution of Pareuchaetes insulata, a biological control agent of Chromolaena odorata in South Africa. BioControl 2016, 61, 815–825. [Google Scholar] [CrossRef]
- May, B.; Coetzee, J. Comparisons of the thermal physiology of water hyacinth biological control agents: Predicting establishment and distribution pre-and post-release. Entomol. Exp. Appl. 2013, 147, 241–250. [Google Scholar] [CrossRef]
- Byrne, M.J.; Currin, S.; Hill, M.P. The influence of climate on the establishment and success of the biocontrol agent Gratiana spadicea, released on Solanum sisymbriifolium in South Africa. Biol. Control 2002, 24, 128–134. [Google Scholar] [CrossRef]
- Chidawanyika, F.; Chikowore, G.; Mutamiswa, R. Thermal tolerance of the biological control agent Neolema abbreviata and its potential geographic distribution together with its host Tradescantia fluminensis in South Africa. Biol. Control 2020, 104315. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Dormann, C.F.; Schymanski, S.J.; Cabral, J.; Chuine, I.; Graham, C.; Hartig, F.; Kearney, M.; Morin, X.; Römermann, C.; Schröder, B. Correlation and process in species distribution models: Bridging a dichotomy. J. Biogeogr. 2012, 39, 2119–2131. [Google Scholar] [CrossRef]
- Kumar, S.; Neven, L.G.; Yee, W.L. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef]
- Maywald, G.F.; Sutherst, R. User’s Guide to CLIMEX. A Computer Program for Comparing Climates in Ecology; CSIRO Division of Entomology: Canberra, Australia, 1985.
- Kriticos, D.; Yonow, T.; McFadyen, R. The potential distribution of Chromolaena odorata (Siam weed) in relation to climate. Weed Res. 2005, 45, 246–254. [Google Scholar] [CrossRef]
- Dhileepan, K.; Senaratne, K.W. How widespread is Parthenium hysterophorus and its biological control agent Zygogramma bicolorata in South Asia? Weed Res. 2009, 49, 557–562. [Google Scholar] [CrossRef]
- Simelane, D.O.; Mawela, K.V. Establishment and increased fitness of the seed-feeding weevil Cissoanthonomus tuberculipennis, a biological control agent for balloon vine Cardiospermum grandiflorum in South Africa. Biol. Control 2019, 135, 83–88. [Google Scholar] [CrossRef]
- Dhileepan, K.; Shi, B.; Callander, J.; Teshome, M.; Neser, S.; Senaratne, K. Gall thrips Acaciothrips ebneri (Thysanoptera: Phlaeothripidae) from Ethiopia, a promising biological control agent for prickly acacia in Australia. Afr. Entomol. 2018, 26, 237–241. [Google Scholar] [CrossRef]
- McCarren, K.L.; Scott, J.K. Host range and potential distribution of the rust fungus, Miyagia pseudosphaeria, a biological control agent for Sonchus species. Australas. Plant Pathol. 2017, 46, 473–482. [Google Scholar] [CrossRef]
- Kumaran, N.; Choudhary, A.; Legros, M.; Sheppard, A.W.; Barrett, L.G.; Gardiner, D.M.; Raghu, S. Gene technologies in weed management: A technical feasibility analysis. Curr. Opin. Insect Sci. 2020, 38, 6–14. [Google Scholar] [CrossRef]
- Webber, B.L.; Raghu, S.; Edwards, O.R. Opinion: Is CRISPR-based gene drive a biocontrol silver bullet or global conservation threat? Proc. Natl. Acad. Sci. USA 2015, 112, 10565–10567. [Google Scholar] [CrossRef]
- Gong, P. Invasive Species Management on Military Lands: Clustered Regularly Interspaced Short Palindromic Repeat/CRISPR Associated Protein 9 (CRISPR/Cas9) Based Gene Drives; US Army Engineer Research and Development Center, Environmental Laboratory: Vicksburg, MS, USA, 2017. [Google Scholar]
- ISAC. Advanced Biotechnology Tools for Invasive Species Management; National Invasive Species Council Secretariat, U.S. Department of the Interior: Washington, DC, USA, 2017.
- Huang, J.; Hirji, R.; Adam, L.; Rozwadowski, K.L.; Hammerlindl, J.K.; Keller, W.A.; Selvaraj, G. Genetic engineering of glycinebetaine production toward enhancing stress tolerance in plants: Metabolic limitations. Plant Physiol. 2000, 122, 747–756. [Google Scholar] [CrossRef]
- Yu, C.; Qiao, G.; Qiu, W.; Yu, D.; Zhou, S.; Shen, Y.; Yu, G.; Jiang, J.; Han, X.; Liu, M.; et al. Molecular breeding of water lily: Engineering cold stress tolerance into tropical water lily. Hortic. Res. 2018, 5, 73. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, R.-R.; Gao, G.-Q.; Ma, M.-Y.; Chen, H. Cold tolerance and silencing of three cold-tolerance genes of overwintering Chinese white pine larvae. Sci. Rep. 2016, 6, 34698. [Google Scholar] [CrossRef]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef]
- Teets, N.M.; Denlinger, D.L. Physiological mechanisms of seasonal and rapid cold-hardening in insects. Physiol. Entomol. 2013, 38, 105–116. [Google Scholar] [CrossRef]
- Teets, N.M.; Gantz, J.D.; Kawarasaki, Y. Rapid cold hardening: Ecological relevance, physiological mechanisms and new perspectives. J. Exp. Biol. 2020, 223, jeb203448. [Google Scholar] [CrossRef]
- Des Marteaux, L.E.; McKinnon, A.H.; Udaka, H.; Toxopeus, J.; Sinclair, B.J. Effects of cold-acclimation on gene expression in Fall field cricket (Gryllus pennsylvanicus) ionoregulatory tissues. BMC Genom. 2017, 18, 357. [Google Scholar] [CrossRef]
- Yamaguchi, A.; Tanaka, S.; Yamaguchi, S.; Kuwahara, H.; Takamura, C.; Imajoh-Ohmi, S.; Horikawa, D.D.; Toyoda, A.; Katayama, T.; Arakawa, K.; et al. Two novel heat-soluble protein families abundantly expressed in an anhydrobiotic tardigrade. PLoS ONE 2012, 7, e44209. [Google Scholar] [CrossRef] [PubMed]
- Grohme, M.A.; Mali, B.; Wełnicz, W.; Michel, S.; Schill, R.O.; Frohme, M. The aquaporin channel repertoire of the tardigrade Milnesium tardigradum. Bioinform. Biol. Insights 2013, 7, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Horikawa, D.D.; Saito, Y.; Kuwahara, H.; Kozuka-Hata, H.; Shin-I, T.; Minakuchi, Y.; Ohishi, K.; Motoyama, A.; Aizu, T.; et al. Extremotolerant tardigrade genome and improved radiotolerance of human cultured cells by tardigrade-unique protein. Nat. Commun. 2016, 7, 12808. [Google Scholar] [CrossRef]
- Boothby, T.C.; Tapia, H.; Brozena, A.H.; Piszkiewicz, S.; Smith, A.E.; Giovannini, I.; Rebecchi, L.; Pielak, G.J.; Koshland, D.; Goldstein, B. Tardigrades use intrinsically disordered proteins to survive desiccation. Mol. Cell 2017, 65, 975–984.e975. [Google Scholar] [CrossRef]
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi efficiency, systemic properties, and novel delivery methods for pest insect control: What we know so far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed]
- Tellam, R.; Vuocolo, T.; Johnson, S.; Jarmey, J.; Pearson, R. Insect chitin synthase. FEBS J. 2000, 269, 6025–6043. [Google Scholar] [CrossRef]
- Tenlen, J.R. Microinjection of dsRNA in Tardigrades. Cold Spring Harb. Protoc. 2018, 2018, prot102368. [Google Scholar] [CrossRef]
- Tenlen, J.R.; McCaskill, S.; Goldstein, B. RNA interference can be used to disrupt gene function in tardigrades. Dev. Genes Evol. 2013, 223, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genom. 2011, 12, 30–43. [Google Scholar] [CrossRef]
- Wang, H.; Russa, M.L.; Qi, L.S. CRISPR/Cas9 in Genome Editing and Beyond. Annu. Rev. Biochem. 2016, 85, 227–264. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.; Zhu, H. CRISPR technology is revolutionizing the improvement of tomato and other fruit crops. Hortic. Res. 2019, 6, 13. [Google Scholar] [CrossRef]
- Mumford, J.D. Science, regulation, and precedent for genetically modified insects. PLoS Negl. Trop. Dis. 2012, 6, e1504. [Google Scholar] [CrossRef][Green Version]
- Ernst, K.C.; Haenchen, S.; Dickinson, K.; Doyle, M.S.; Walker, K.; Monaghan, A.J.; Hayden, M.H. Awareness and support of release of genetically modified “sterile” mosquitoes, Key West, Florida, USA. Emerg. Infect. Dis. 2015, 21, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Amin, L.; Hashim, H. Factors influencing stakeholders attitudes toward genetically modified Aedes mosquito. Sci. Eng. Ethics 2015, 21, 655–681. [Google Scholar] [CrossRef] [PubMed]
- Kola, V.S.R.; Renuka, P.; Madhav, M.S.; Mangrauthia, S.K. Key enzymes and proteins of crop insects as candidate for RNAi based gene silencing. Front. Physiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, A.; Hill, R.S.; Declerck-Floate, R.; McClay, A.; Olckers, T.; Quimby, P.C.; Zimmermann, H. A global review of risk-benefit-cost analysis for the introduction of classical biological control agents against weeds: A crisis in the making? Biocontrol News Inform. 2003, 24, 91–108. [Google Scholar]
- Balciunas, J.; Coombs, E.M. International code of best practices for classical biological control of weed. In Biological Control of Invasive Plants in the US; Coombs, E.M., Clark, J.K., Piper, G.L., Cofrancesco, A.F., Eds.; Oregon State University Press: Corvallis, OR, USA, 2004; pp. 130–136. [Google Scholar]
- Paterson, I.D.; Coetzee, J.A.; Weyl, P.; Griffith, T.C.; Voogt, N.; Hill, M.P. Cryptic species of a water hyacinth biological control agent revealed in South Africa: Host specificity, impact, and thermal tolerance. Entomol. Exp. Appl. 2019, 167, 682–691. [Google Scholar] [CrossRef]
- United States 7 Code of Federal Regulations Part 330. Available online: https://beta.regulations.gov/document/APHIS-2008-0076-0093 (accessed on 7 January 2021).
- Western Australia Biosecurity and Agriculture Management Act 2007. Available online: https://www.legislation.wa.gov.au/legislation/statutes.nsf/law_a146629.html (accessed on 7 January 2021).
- National Environmental Management: Biodiversity Act 2014. Available online: https://www.environment.gov.za/sites/default/files/legislations/nemba10of2004_alienandinvasive_speciesregulations.pdf (accessed on 12 January 2021).



{kind=link}
{kind=link}
| Cold Tolerance Metric | What It Measures | Effort Required | Ecological Relevance | Example Reference |
|---|---|---|---|---|
| Super Cooling Point (SCP) | Failure; the temperature at which ice crystals form in insect tissues. | Low | Delineates lower bound for freeze-intolerant taxa. Not useful for chill-susceptible taxa, as most will have died before supercooling. Not useful for freeze-tolerant taxa, as mortality will depend on temperatures and exposure times below the SCP. May be correlated with other cold tolerance metrics. Useful for distinguishing chill-susceptible and freeze-avoidant taxa, and quick comparison among populations. | Zachariassen [88] |
| Lower Lethal Temperature (LLT) | Failure; the highest (cold) temperature which results in a prescribed rate of mortality (e.g., LT50, LT100). | Low | Delineates lower bound for all cold tolerance categories, but is confounded by exposure time. Will be equal to the SCP for freeze-avoidant taxa. | Sinclair et al. [72] |
| Lethal Time (LTi) | Failure; the length of time at a given temperature which results in a prescribed rate of mortality. | Low | Confounded with LLT; useful for comparing relative cold tolerance among populations. | Andersen et al. [80] |
| Sum of Injurious Temperatures (SIT) | Failure; the degree-day relationship between exposure temperature and time which results in 50% mortality. | High | SIT and ULCIZ are determined together, and represent a comprehensive measure of chill induced mortality. ULCIZ delineates the isotherms between which there should be no expected cold-related mortality. Most relevant for delineating bounds of chill-susceptible taxa. | Nedvěd et al. [78] |
| Upper Limit of Chill Injury Zone (ULCIZ) | Failure; the lowest temperature which results in no chill injury, regardless of exposure duration. | High | Zhao et al. [23] | |
| Critical Thermal Minimum (CTmin)/Chill Coma Induction Temperature (CCIT) | Failure; the temperature, in a decreasing ramp, preceding loss of coordination (i.e., chill coma). | Medium | CCIT and CCRT are best used as relative measures of cold tolerance among populations. Due to their relative ease to measure, these may be useful for screening for cold tolerance traits; however, they are of limited use for species distribution models (SDM). CCRT may be preferable due to its relative ease to conduct. | Andersen et al. [89] |
| Chill Coma Recovery Time (CCRT) | Failure; the time until spontaneous movement or coordination is regained upon warming, after induction of chill coma. | Low | Gibert et al. [90] | |
| Temperature-Dependent Development (TDD) | Performance; rate of development along a temperature gradient. | High | Development time determines how quickly agents can build to effective population sizes, both within a single generation and over multiple generations. Can be used to model performance within a SDM. | Augustinus et al. [91] |
| Temperature-Dependent Performance (fecundity, etc.) | Performance; feeding, fecundity along a temperature gradient. | High | Similar to TDD; can predict reproductive success as well as potential impact. Has also been used in SDMs. | Reddy et al. [10] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harms, N.E.; Knight, I.A.; Pratt, P.D.; Reddy, A.M.; Mukherjee, A.; Gong, P.; Coetzee, J.; Raghu, S.; Diaz, R. Climate Mismatch between Introduced Biological Control Agents and Their Invasive Host Plants: Improving Biological Control of Tropical Weeds in Temperate Regions. Insects 2021, 12, 549. https://doi.org/10.3390/insects12060549
Harms NE, Knight IA, Pratt PD, Reddy AM, Mukherjee A, Gong P, Coetzee J, Raghu S, Diaz R. Climate Mismatch between Introduced Biological Control Agents and Their Invasive Host Plants: Improving Biological Control of Tropical Weeds in Temperate Regions. Insects. 2021; 12(6):549. https://doi.org/10.3390/insects12060549
Chicago/Turabian StyleHarms, Nathan E., Ian A. Knight, Paul D. Pratt, Angelica M. Reddy, Abhishek Mukherjee, Ping Gong, Julie Coetzee, S. Raghu, and Rodrigo Diaz. 2021. "Climate Mismatch between Introduced Biological Control Agents and Their Invasive Host Plants: Improving Biological Control of Tropical Weeds in Temperate Regions" Insects 12, no. 6: 549. https://doi.org/10.3390/insects12060549
APA StyleHarms, N. E., Knight, I. A., Pratt, P. D., Reddy, A. M., Mukherjee, A., Gong, P., Coetzee, J., Raghu, S., & Diaz, R. (2021). Climate Mismatch between Introduced Biological Control Agents and Their Invasive Host Plants: Improving Biological Control of Tropical Weeds in Temperate Regions. Insects, 12(6), 549. https://doi.org/10.3390/insects12060549