
Warm Temperatures Reduce Flower Attractiveness and Bumblebee Foraging


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Insects
2.3. Experimental Plant Growth Conditions
2.4. Visual Floral Trait and Flower Reward Measurements
2.4.1. Visual Signals
2.4.2. Floral Resources
2.5. Experimental Design of Plant–Insect Interactions
2.6. Training Phase and Visiting Observations
2.7. Statistical Analyses
3. Results
3.1. Effects of Higher Temperature on Plant Attractiveness
3.1.1. Floral Traits
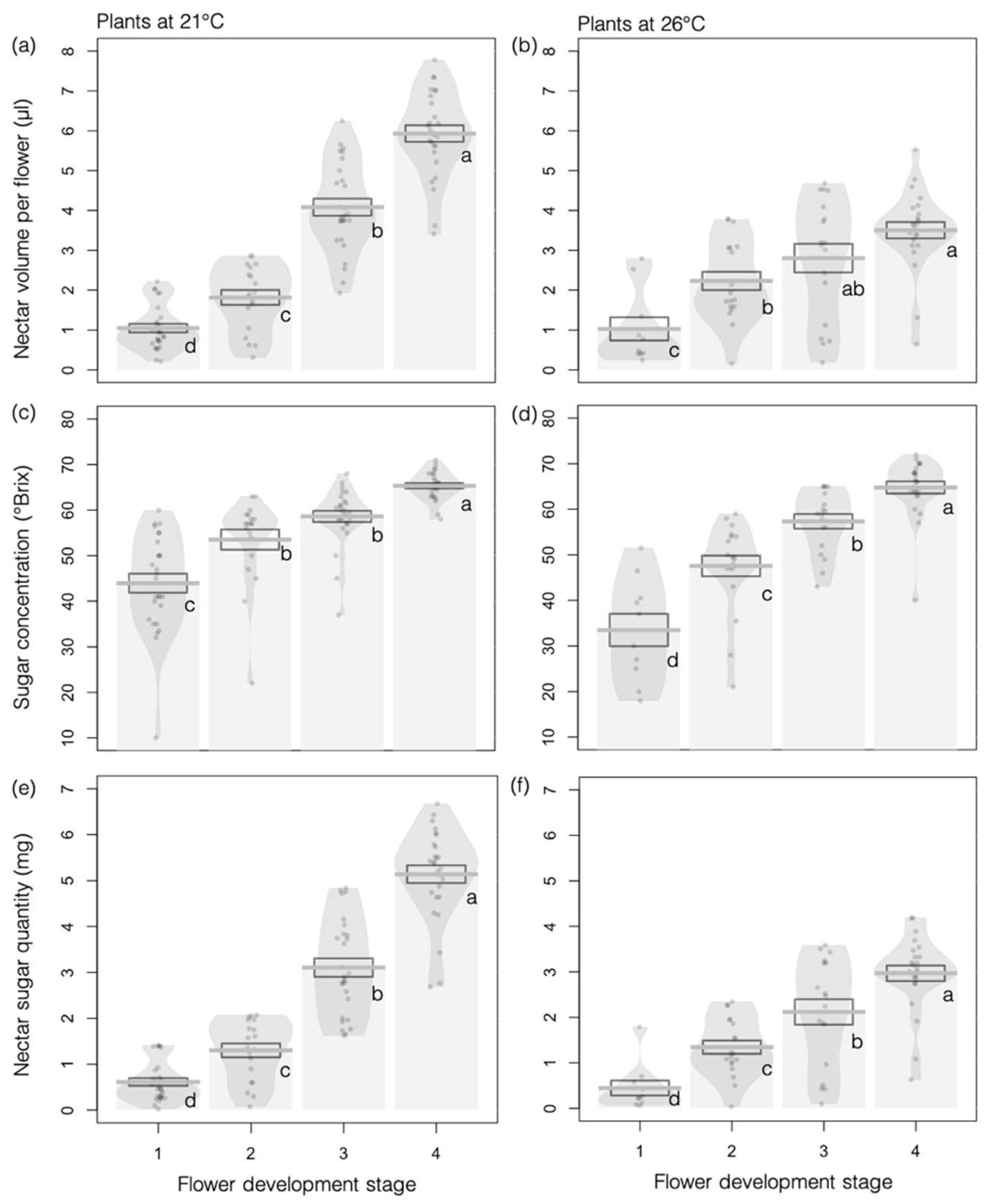
3.1.2. Floral Rewards
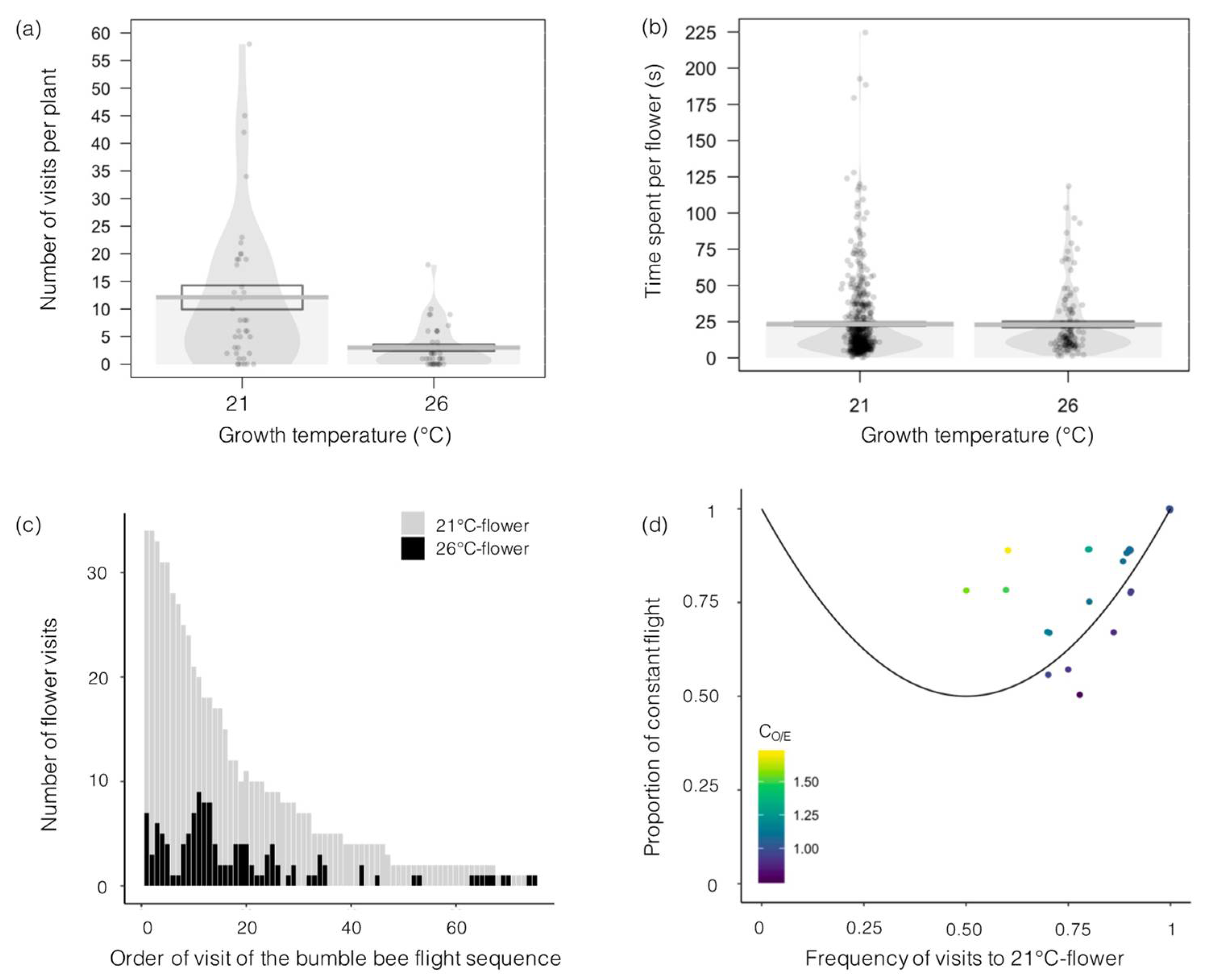
3.2. Effects of Higher Temperature on Plant–Insect Interactions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPBES. Thematic Assessment of Pollinators, Pollination and Food Production. Available online: http://www.ipbes.net/publication/thematic-assessment-pollinators-pollination-and-food-production (accessed on 29 November 2016).
- Soroye, P.; Newbold, T.; Kerr, J. Climate Change Contributes to Widespread Declines among Bumble Bees across Continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.-M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. Royal Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Aizen, M.A.; Aguiar, S.; Biesmeijer, J.C.; Garibaldi, L.A.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Morales, C.L.; Ngo, H.; et al. Global Agricultural Productivity Is Threatened by Increasing Pollinator Dependence without a Parallel Increase in Crop Diversification. Glob. Chang. Biol. 2019, 25, 3516–3527. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, J.E.; Griffin, S.R.; Gezon, Z.J.; Inouye, B.D.; Underwood, N.; Inouye, D.W.; Irwin, R.E. Interannual Bumble Bee Abundance Is Driven by Indirect Climate Effects on Floral Resource Phenology. Ecol. Lett. 2017, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Gérard, M.; Vanderplanck, M.; Wood, T.; Michez, D. Global Warming and Plant–Pollinator Mismatches. Emerg. Top Life Sci. 2020. [Google Scholar] [CrossRef]
- Urban, M.C. Accelerating Extinction Risk from Climate Change. Science 2015, 348, 571–573. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature Extremes: Effect on Plant Growth and Development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Korres, N.E.; Norsworthy, J.K.; Tehranchian, P.; Gitsopoulos, T.K.; Loka, D.A.; Oosterhuis, D.M.; Gealy, D.R.; Moss, S.R.; Burgos, N.R.; Miller, M.R.; et al. Cultivars to Face Climate Change Effects on Crops and Weeds: A Review. Agron. Sustain. Dev. 2016, 36, 12. [Google Scholar] [CrossRef]
- Hedhly, A. Sensitivity of Flowering Plant Gametophytes to Temperature Fluctuations. Environ. Exp. Bot. 2011, 74, 9–16. [Google Scholar] [CrossRef]
- Aubert, L.; Konrádová, D.; Kebbas, S.; Barris, S.; Quinet, M. Comparison of High Temperature Resistance in Two Buckwheat Species Fagopyrum Esculentum and Fagopyrum Tataricum. J. Plant Physiol. 2020, 251, 153222. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The Effect of Drought and Heat Stress on Reproductive Processes in Cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-H.; Offler, C.E.; Ruan, Y.-L. Regulation of Fruit and Seed Response to Heat and Drought by Sugars as Nutrients and Signals. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Lemoine, R.; Camera, S.L.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.-L.; Laloi, M.; Coutos-Thévenot, P.; Maurousset, L.; et al. Source-to-Sink Transport of Sugar and Regulation by Environmental Factors. Front Plant Sci. 2013, 4. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Staggenborg, S.A.; Ristic, Z. Impacts of Drought and/or Heat Stress on Physiological, Developmental, Growth, and Yield Processes of Crop Plants. In Response Crops Ltd. Water Underst. Modeling Water Stress Effects Plant Growth Process; American Society of Agronomy, Inc., Crop Science Society of America, Inc., Soil Science Society of America, Inc.: Madison, WI, USA, 2008; pp. 301–355. [Google Scholar] [CrossRef]
- Scaven, V.L.; Rafferty, N.E. Physiological Effects of Climate Warming on Flowering Plants and Insect Pollinators and Potential Consequences for Their Interactions. Curr. Zool. 2013, 59, 418–426. [Google Scholar] [CrossRef]
- Chittka, L.; Raine, N.E. Recognition of Flowers by Pollinators. Curr. Opin. Plant Biol. 2006, 9, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.A.; Schiestl, F.P. The Evolution of Floral Scent: The Influence of Olfactory Learning by Insect Pollinators on the Honest Signalling of Floral Rewards. Funct. Ecol. 2009, 23, 841–851. [Google Scholar] [CrossRef]
- Leonard, A.S.; Dornhaus, A.; Papaj, D.R. Forget-Me-Not: Complex Floral Displays, Inter-Signal Interactions, and Pollinator Cognition. Curr. Zool. 2011, 57, 215–224. [Google Scholar] [CrossRef]
- Knauer, A.C.; Schiestl, F.P. Bees Use Honest Floral Signals as Indicators of Reward When Visiting Flowers. Ecol. Lett. 2015, 18, 135–143. [Google Scholar] [CrossRef]
- Ruxton, G.D.; Schaefer, H.M. Floral Colour Change as a Potential Signal to Pollinators. Curr. Opin. Plant Biol. 2016, 32, 96–100. [Google Scholar] [CrossRef]
- Sprayberry, J.D.H. The Prevalence of Olfactory- versus Visual-Signal Encounter by Searching Bumblebees. Sci. Rep. 2018, 8, 14590. [Google Scholar] [CrossRef] [PubMed]
- Fowler, R.E.; Rotheray, E.L.; Goulson, D. Floral Abundance and Resource Quality Influence Pollinator Choice. Insect Conserv. Divers. 2016, 9, 481–494. [Google Scholar] [CrossRef]
- Junker, R.R.; Parachnowitsch, A.L. Working towards a Holistic View on Flower Traits-How Floral Scents Mediate Plant-Animal Interactions in Concert with Other Floral Characters. J. Indian Inst. Sci. 2015, 95, 43–67. [Google Scholar]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Nicolson, S.W.; Thornburg, R.W. Nectar chemistry. In Nectaries and Nectar; Springer: Berlin/Heidelberg, Germany, 2007; pp. 215–264. [Google Scholar]
- Vaudo, A.D.; Tooker, J.F.; Grozinger, C.M.; Patch, H.M. Bee Nutrition and Floral Resource Restoration. Curr. Opin. Insect Sci. 2015, 10, 133–141. [Google Scholar] [CrossRef]
- Cnaani, J.; Thomson, J.D.; Papaj, D.R. Flower Choice and Learning in Foraging Bumblebees: Effects of Variation in Nectar Volume and Concentration. Ethology 2006, 112, 278–285. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Pianka, E.R. On Optimal Use of a Patchy Environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef]
- Jones, P.L.; Ryan, M.J.; Chittka, L. The Influence of Past Experience with Flower Reward Quality on Social Learning in Bumblebees. Anim. Behav. 2015, 101, 11–18. [Google Scholar] [CrossRef]
- Zhao, Z.; Lu, N.; Conner, J.K. Adaptive Pattern of Nectar Volume within Inflorescences: Bumblebee Foraging Behavior and Pollinator-Mediated Natural Selection. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Gumbert, A. Color Choices by Bumble Bees (Bombus Terrestris): Innate Preferences and Generalization after Learning. Behav. Ecol. Sociobiol. 2000, 48, 36–43. [Google Scholar] [CrossRef]
- Cartar, R.V. Resource Tracking by Bumble Bees: Responses to Plant-Level Differences in Quality. Ecology 2004, 85, 2764–2771. [Google Scholar] [CrossRef]
- Kudo, G.; Harder, L.D. Floral and Inflorescence Effects on Variation in Pollen Removal and Seed Production among Six Legume Species. Funct. Ecol. 2005, 19, 245–254. [Google Scholar] [CrossRef]
- Larsson, M.; Franzén, M. Critical Resource Levels of Pollen for the Declining Bee Andrena Hattorfiana (Hymenoptera, Andrenidae). Biol. Conserv. 2007, 134, 405–414. [Google Scholar] [CrossRef]
- Wallisdevries, M.F.; Van Swaay, C.A.M.; Plate, C.L. Changes in Nectar Supply: A Possible Cause of Widespread Butterfly Decline. Curr. Zool. 2012, 58, 384–391. [Google Scholar] [CrossRef]
- Somme, L.; Vanderplanck, M.; Michez, D.; Lombaerde, I.; Moerman, R.; Wathelet, B.; Wattiez, R.; Lognay, G.; Jacquemart, A.-L. Pollen and Nectar Quality Drive the Major and Minor Floral Choices of Bumble Bees. Apidologie 2014, 46, 92–106. [Google Scholar] [CrossRef]
- Bailes, E.J.; Ollerton, J.; Pattrick, J.G.; Glover, B.J. How Can an Understanding of Plant–Pollinator Interactions Contribute to Global Food Security? Curr. Opin. Plant Biol. 2015, 26, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Höfer, R.J.; Ayasse, M.; Kuppler, J. Bumblebee Behavior on Flowers, but Not Initial Attraction, Is Altered by Short-Term Drought Stress. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Kuppler, J.; Wieland, J.; Junker, R.R.; Ayasse, M. Drought-Induced Reduction in Flower Size and Abundance Correlates with Reduced Flower Visits by Bumble Bees. AoB Plants 2021. [Google Scholar] [CrossRef]
- Descamps, C.; Quinet, M.; Jacquemart, A.-L. The Effects of Drought on Plant–Pollinator Interactions: What to Expect? Environ. Exp. Bot. 2021, 182, 104297. [Google Scholar] [CrossRef]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature Stress and Plant Sexual Reproduction: Uncovering the Weakest Links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef]
- Mu, J.; Peng, Y.; Xi, X.; Wu, X.; Li, G.; Niklas, K.J.; Sun, S. Artificial Asymmetric Warming Reduces Nectar Yield in a Tibetan Alpine Species of Asteraceae. Ann. Bot. 2015, 116, 899–906. [Google Scholar] [CrossRef]
- Petanidou, T.; Smets, E. Does Temperature Stress Induce Nectar Secretion in Mediterranean Plants? New Phytol. 1996, 133, 513–518. [Google Scholar] [CrossRef]
- Pacini, E.; Nepi, M.; Vesprini, J.L. Nectar Biodiversity: A Short Review. Plant Syst. Evol. 2003, 238, 7–21. [Google Scholar] [CrossRef]
- Takkis, K.; Tscheulin, T.; Tsalkatis, P.; Petanidou, T. Climate Change Reduces Nectar Secretion in Two Common Mediterranean Plants. AoB Plants 2015, 7, plv111. [Google Scholar] [CrossRef] [PubMed]
- Mesihovic, A.; Iannacone, R.; Firon, N.; Fragkostefanakis, S. Heat Stress Regimes for the Investigation of Pollen Thermotolerance in Crop Plants. Plant Reprod 2016, 29, 93–105. [Google Scholar] [CrossRef]
- Descamps, C.; Quinet, M.; Baijot, A.; Jacquemart, A.-L. Temperature and Water Stress Affect Plant–Pollinator Interactions in Borago Officinalis (Boraginaceae). Ecol. Evol. 2018, 8, 3443–3456. [Google Scholar] [CrossRef]
- Thom, M.D.; Eberle, C.A.; Forcella, F.; Gesch, R.; Weyers, S.; Lundgren, J.G. Nectar Production in Oilseeds: Food for Pollinators in an Agricultural Landscape. Crop Sci. 2016, 56, 727–739. [Google Scholar] [CrossRef]
- Baijot, A. Etude Des Impacts Du Stress Hydrique et de Hausses de Temp ́eratures Sur Les Ressources Florales de La Bourrache (Borago Officinalis) et Des r ́epercussions Possibles Sur Les Relations Plantes-Pollinisateurs; Faculté des Bioingénieurs: Louvain-la-Neuve, Belgique, 2016. [Google Scholar]
- Abramoff, M.D.; Magalhães, P.J.; Ram, S.J. Image Processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Prys-Jones, O.E.; Corbet, S.A. Naturalists’ Handbooks 6: Bumblebees; Richmond Publishing Compagny: Slough, UK, 1991. [Google Scholar]
- Ishii, H.S. Analysis of Bumblebee Visitation Sequences within Single Bouts: Implication of the Overstrike Effect on Short-Term Memory. Behav. Ecol. Sociobiol. 2005, 57, 599–610. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Brunet, J.; Van Etten, M.L. The Response of Floral Traits Associated with Pollinator Attraction to Environmental Changes Expected under Anthropogenic Climate Change in High-Altitude Habitats. Int. J. Plant Sci. 2019, 180, 954–964. [Google Scholar] [CrossRef]
- Liu, Y.; Mu, J.; Niklas, K.J.; Li, G.; Sun, S. Global Warming Reduces Plant Reproductive Output for Temperate Multi-Inflorescence Species on the Tibetan Plateau. New Phytol. 2012, 195, 427–436. [Google Scholar] [CrossRef]
- Takkis, K.; Tscheulin, T.; Petanidou, T. Differential Effects of Climate Warming on the Nectar Secretion of Early- and Late-Flowering Mediterranean Plants. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Descamps, C.; Marée, S.; Hugon, S.; Quinet, M.; Jacquemart, A.-L. Species-Specific Responses to Combined Water Stress and Increasing Temperatures in Two Bee-Pollinated Congeners (Echium, Boraginaceae). Ecol. Evol. 2020, 10, 6549–6561. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Kurepin, L.V.; Reid, D.M. Effects of Temperature and Watering Regime on Growth, Gas Exchange and Abscisic Acid Content of Canola (Brassica Napus) Seedlings. Environ. Exp. Bot. 2012, 75, 107–113. [Google Scholar] [CrossRef]
- Porch, T.G.; Jahn, M. Effects of High-Temperature Stress on Microsporogenesis in Heat-Sensitive and Heat-Tolerant Genotypes of Phaseolus Vulgaris. Plant Cell Environ. 2001, 24, 723–731. [Google Scholar] [CrossRef]
- Paupière, M.J.; Müller, F.; Li, H.; Rieu, I.; Tikunov, Y.M.; Visser, R.G.F.; Bovy, A.G. Untargeted Metabolomic Analysis of Tomato Pollen Development and Heat Stress Response. Plant Reprod 2017, 30, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; Willmer, P. Linking Bees and Flowers: How Do Floral Communities Structure Pollinator Communities? Ecology 2003, 84, 2628–2642. [Google Scholar] [CrossRef]
- Rowe, L.; Gibson, D.; Bahlai, C.A.; Gibbs, J.; Landis, D.A.; Isaacs, R. Flower Traits Associated with the Visitation Patterns of Bees. Oecologia 2020, 193, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Ouvrard, P.; Transon, J.; Jacquemart, A.-L. Flower-Strip Agri-Environment Schemes Provide Diverse and Valuable Summer Flower Resources for Pollinating Insects. Biodivers Conserv. 2018, 27, 2193–2216. [Google Scholar] [CrossRef]
- Cakmak, I.; Sanderson, C.; Blocker, T.D.; Lisa Pham, L.; Checotah, S.; Norman, A.A.; Harader-Pate, B.K.; Tyler Reidenbaugh, R.; Nenchev, P.; Barthell, J.F.; et al. Different Solutions by Bees to a Foraging Problem. Anim. Behav. 2009, 77, 1273–1280. [Google Scholar] [CrossRef]
- Latty, T.; Trueblood, J.S. How Do Insects Choose Flowers? A Review of Multi-Attribute Flower Choice and Decoy Effects in Flower-Visiting Insects. J. Anim. Ecol. 2020, 89, 2750–2762. [Google Scholar] [CrossRef]
- Carvell, C.; Bourke, A.F.G.; Dreier, S.; Freeman, S.N.; Hulmes, S.; Jordan, W.C.; Redhead, J.W.; Sumner, S.; Wang, J.; Heard, M.S. Bumblebee Family Lineage Survival Is Enhanced in High-Quality Landscapes. Nature 2017, 543, 547–549. [Google Scholar] [CrossRef]
- Raguso, R.A. Why Do Flowers Smell? The Chemical Ecology of Fragrance-Driven Pollination. Adv. Insect Chem. Ecol. 2004, 151–178. [Google Scholar]
- Leonard, A.S.; Masek, P. Multisensory Integration of Colors and Scents: Insights from Bees and Flowers. J. Comp. Physiol. A 2014, 200, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Leadbeater, E.; Chittka, L. Social Learning in Insects—From Miniature Brains to Consensus Building. Curr. Biol. 2007, 17, R703–R713. [Google Scholar] [CrossRef] [PubMed]
- Austin, M.W.; Horack, P.; Dunlap, A.S. Choice in a Floral Marketplace: The Role of Complexity in Bumble Bee Decision-Making. Behav. Ecol. 2019, 30, 500–508. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Descamps, C.; Jambrek, A.; Quinet, M.; Jacquemart, A.-L. Warm Temperatures Reduce Flower Attractiveness and Bumblebee Foraging. Insects 2021, 12, 493. https://doi.org/10.3390/insects12060493
Descamps C, Jambrek A, Quinet M, Jacquemart A-L. Warm Temperatures Reduce Flower Attractiveness and Bumblebee Foraging. Insects. 2021; 12(6):493. https://doi.org/10.3390/insects12060493
Chicago/Turabian StyleDescamps, Charlotte, Anne Jambrek, Muriel Quinet, and Anne-Laure Jacquemart. 2021. "Warm Temperatures Reduce Flower Attractiveness and Bumblebee Foraging" Insects 12, no. 6: 493. https://doi.org/10.3390/insects12060493
APA StyleDescamps, C., Jambrek, A., Quinet, M., & Jacquemart, A.-L. (2021). Warm Temperatures Reduce Flower Attractiveness and Bumblebee Foraging. Insects, 12(6), 493. https://doi.org/10.3390/insects12060493