
Costs and Benefits of Wax Production in the Larvae of the Ladybeetle Scymnus nubilus



{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Insect Rearing
2.2. Development Time, Adult Body Weight and Ovarioles Number
2.3. Behavioral Compensation: Feeding Parameters
2.4. Intraguild Predation
2.5. Statistical Analysis
3. Results
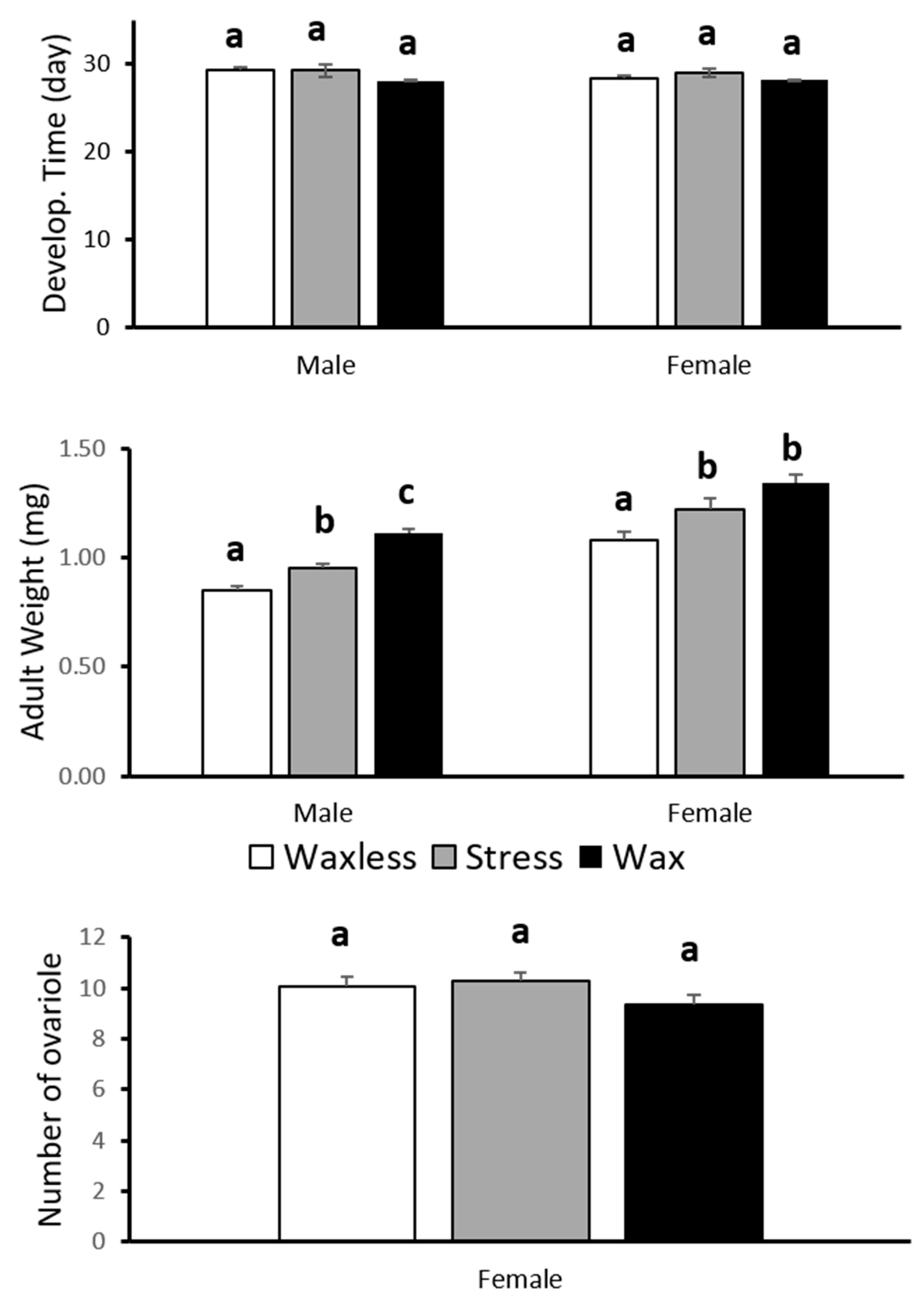
3.1. Development Time, Adult Body Weight and Ovarioles Number
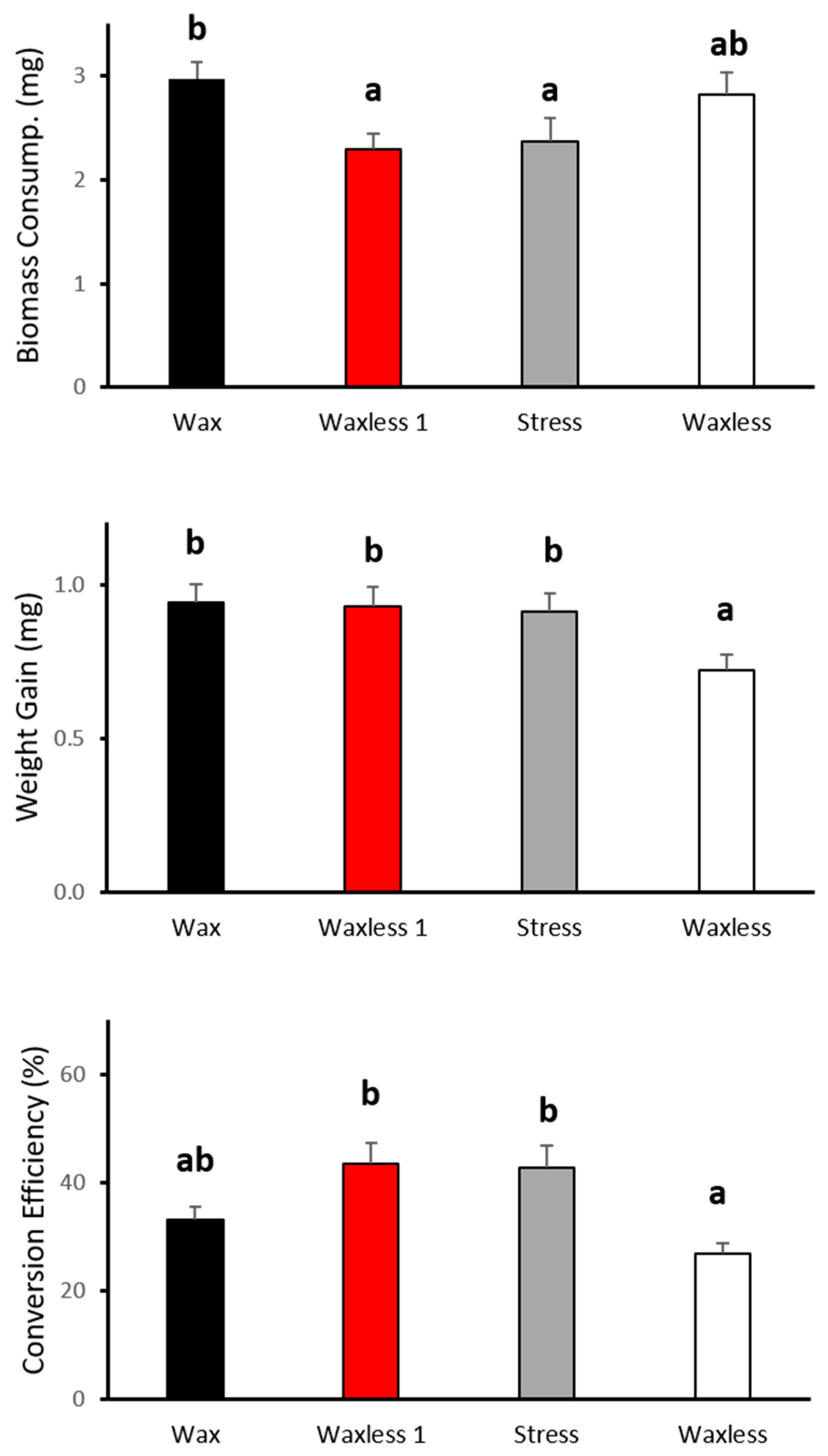
3.2. Behavioral Compensation: Feeding Parameters
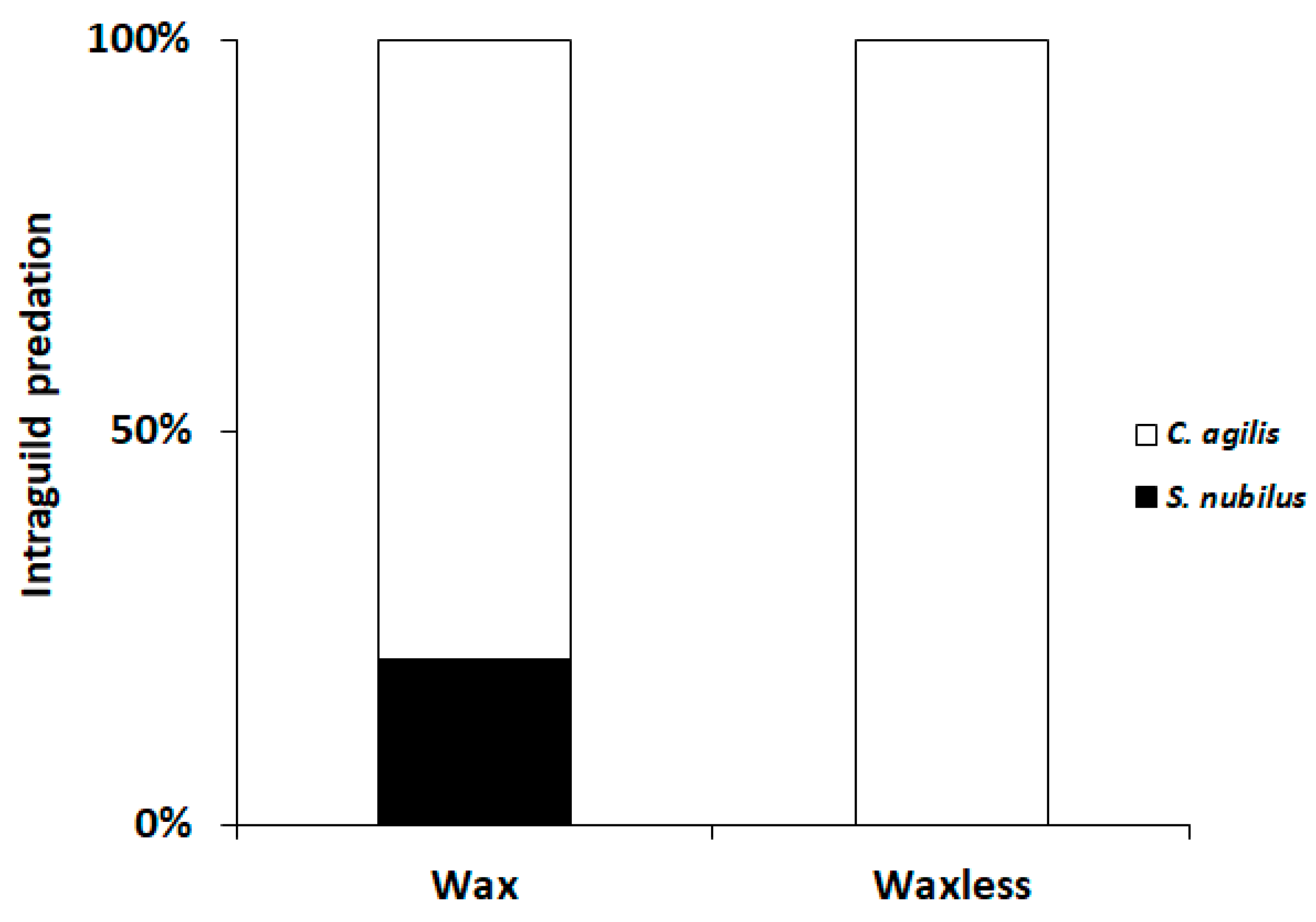
3.3. Defensive Value against Intraguild Predation
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gorb, E.V.; Gorb, S.N. Anti-adhesive effects of plant wax coverage on insect attachment. J. Expl. Bot. 2017, 68, 5323–5337. [Google Scholar] [CrossRef]
- Schoeller, E.N.; Yassin, M.; Redak, R.A. Host-produced wax affects the searching behavior and efficacy of parasitoids of the giant whitefly Aleurodicus dugesii (Hemiptera: Aleyrodidae). Biol. Control 2018, 121, 74–79. [Google Scholar] [CrossRef]
- Yamazaki, K. White plant shoots, wax-producing insects and other white structures made by arthropods: A mimicry complex? Eur. J. Entomol. 2017, 114, 343–349. [Google Scholar] [CrossRef]
- Pope, R.D. Wax production by coccinellid larvae (Coleoptera). Syst. Entomol. 1979, 4, 171–196. [Google Scholar] [CrossRef]
- Agarwala, B.K.; Yasuda, H. Larval interactions in aphidophagous predators: Effectiveness of wax cover as defence shield of Scymnus larvae against predation from syrphids. Entomol. Exp. Appl. 2001, 100, 101–107. [Google Scholar] [CrossRef]
- Wheeler, A.G., Jr.; Stocks, I.C. Ceraeochrysa lineaticornis (Fitch) (Neuroptera: Chrysopidae): Larval plant associations and construction of debris packets from wax of planthoppers (Hemiptera: Flatidae). Proc. Entomol. Soc. Wash. 2019, 121, 299–310. [Google Scholar] [CrossRef]
- Borges, I.; Nóia, M.; Camarinho, R.; Rodrigues, A.S.; Soares, A. Characterization of the alimentary canal of the aphidophagous ladybird, Adalia bipunctata (Coleoptera: Coccinellidae): Anatomical and histological approach. Entomol. Sci. 2015, 18, 66–73. [Google Scholar] [CrossRef]
- Honěk, A.; Dixon, A.F.G.; Soares, A.O.; Skuhrovec, J.; Martinkova, Z. Spatial and temporal changes in the abundance of species and species composition of communities of ladybirds (Coleoptera: Coccinellidae). Curr. Opin. Insect. Sci. 2017, 20, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wu, H.; Pang, H.; De Clercq, P. An artificial diet containing plant pollen for the mealybug predator Cryptolaemus montrouzieri. Pest Manag. Sci. 2017, 73, 541–545. [Google Scholar] [CrossRef]
- Sloggett, J.J.; Magro, A.; Verheggen, F.J.; Hemptinne, J.L.; Hutchison, W.D.; Riddick, E.W. The chemical ecology of Harmonia axyridis. BioControl 2011, 56, 643–661. [Google Scholar] [CrossRef]
- Lucas, E. Intraguild interactions. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., Honěk, A., van Emden, H.F., Eds.; Wiley-Blackwell Publishing: London, UK, 2012; pp. 343–374. [Google Scholar]
- Hemptinne, J.L.; Dixon, A.F.G. Defense, oviposition and sex: Semiochemical parsimony in two species of ladybird beetles (Coleoptera, Coccinellidae)? A short review. Eur. J. Entomol. 2000, 97, 443–447. [Google Scholar] [CrossRef]
- Lucas, E. Intraguild predation among aphidophagous predators. Eur. J. Entomol. 2005, 102, 351–364. [Google Scholar] [CrossRef]
- Hayashi, M.; Nomura, N. Larvae of the green lacewing Mallada desjardinsi (Neuroptera: Chrysopidae) protect themselves against aphid-tending ants by carrying dead aphids on their backs. Appl. Entomol. Zool. 2011, 46, 407–413. [Google Scholar] [CrossRef]
- Völkl, W.; Vohland, K. Wax covers in larvae of two Scymnus species: Do they enhance coccinellid larval survival? Oecologia 1996, 107, 498–503. [Google Scholar] [CrossRef]
- Merlin, J.; Lemaitre, O.; Gregoire, J.C. Oviposition in Cryptolaemus montrouzieri stimulated by wax filaments of its prey. Entomol. Exp. Appl. 1996, 79, 141–146. [Google Scholar] [CrossRef]
- Merlin, J.; Lemaitre, O.; Gregoire, J.C. Chemical cues produced by conspecific larvae deter oviposition by the coccidophagous ladybird beetle, Cryptolaemus montrouzieri. Entomol. Exp. Appl. 1996, 79, 147–151. [Google Scholar] [CrossRef]
- Soares, A.O.; Honěk, A.; Martinkova, Z.; Skuhrovec, J.; Cardoso, P.; Borges, I. Harmonia axyridis failed to establish in the Azores: The role of specie richness, intraguild interactions and resource availability. BioControl 2017, 62, 423–434. [Google Scholar] [CrossRef]
- Moniz, J.; Santos, A.M.C.; Mendonça, E.; Borges, P.A.V. Os artrópodes auxiliares generalistas associados às Macieiras na ilha Terceira (Açores). In Problemas Fitossanitários e Fauna Auxiliar Das Macieiras na Ilha Terceira; Pimentel, R., Lopes, D.H., Cabrera, R., Borges, P.A.V., Machado, A.C., Mumford, J.D., Mexia, A., Eds.; Centro de Biotecnologia dos Açores: Angra do Heroísmo, Portugal, 2009; pp. 61–77. [Google Scholar]
- Borges, I. Life History Evolution in Aphidophagous and Coccidophagous Coccinellidae (Coleoptera). Ph.D. Dissertation, University of the Azores, Ponta Delgada, Portugal, 2008. [Google Scholar]
- Borges, I.; Soares, A.O.; Magro, A.; Hemptinne, J.L. Prey availability in time and space is a driving force in life history evolution of predatory insects. Evol. Ecol. 2011, 25, 1307–1319. [Google Scholar] [CrossRef]
- Uygun, N.; Atlihan, R. The effect of temperature on development and fecundity of Scymnus levaillanti. BioControl 2000, 45, 453–462. [Google Scholar] [CrossRef]
- Takabayashi, J.; Takahashi, S. Role of the scale wax of Ceroplastes ceriferus Anderson (Hemiptera: Coccidae) as a defense against the parasitic wasp Anicetus ceroplastis Ishii (Hymenoptera: Encyrtidae). J. Insect. Behav. 1993, 6, 107–115. [Google Scholar] [CrossRef]
- Eisner, T. Integumental slime and wax secretion: Defensive adaptations of sawfly larvae. J. Chem. Ecol. 1994, 20, 2743–2749. [Google Scholar] [CrossRef]
- Schwartzberg, E.G.; Haynes, K.F.; Johnson, D.W.; Brown, G.C. Wax structures of Scymnus louisianae attenuate aggression from aphid-tending ants. Environ. Entomol. 2010, 39, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Majerus, M.E.N.; Sloggett, J.J.; Godeau, J.F.; Hemptinne, J.L. Interactions between ants and aphidophagous and coccidophagous ladybirds. Popul. Ecol. 2007, 49, 15–27. [Google Scholar] [CrossRef]
- Seago, A.E.; Giorgi, J.A.; Li, J.; Ślipiński, A. Phylogeny, classification and evolution of ladybird beetles (Coleoptera: Coccinellidae) based on simultaneous analysis of molecular and morphological data. Mol. Phylogenet. Evol. 2011, 60, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Lucas, E.; Coderre, E.; Brodeur, J. Intraguild predation among aphid predators: Characterization and influence of extraguild prey density. Ecology 1998, 79, 1084–1092. [Google Scholar] [CrossRef]
- Félix, S.; Soares, A.O. Intraguild predation between the aphidophagous ladybird beetles Harmonia axyridis and Coccinella undecimpunctata (Coleoptera: Coccinellidae): The role of body weight. Eur. J. Entomol. 2004, 101, 237–242. [Google Scholar] [CrossRef]
- Kaneko, S. Predator and parasitoid attacking ant-attended aphids: Effects of predator presence and attending ant species on emerging parasitoid numbers. Ecol. Res. 2007, 22, 451–458. [Google Scholar] [CrossRef]
- Stearns, S.C. The Evolution of Life Historie; Oxford University Press Inc.: New York, NY, USA, 1992; p. 264. [Google Scholar]
- Blanckenhorn, W.U.; Hoeck, P.E.A.; Reim, C.; Teuschl, Y. A cost of being large: Genetically large yellow dung flies lose out in intra-specific food competition. Evol. Ecol. 2011, 25, 875–884. [Google Scholar] [CrossRef][Green Version]
- Harrison, J.F.; Cease, A.J.; Vandenbrooks, J.M.; Albert, T.; Davidowitz, G. Caterpillars selected for large body size and short development time are more susceptible to oxygen-related stress. Ecol. Evol. 2013, 3, 1305–1316. [Google Scholar] [CrossRef]
- Borges, I.; Hemptinne, J.L.; Soares, A.O. Contrasting population growth parameters of the aphidophagous Scymnus nubilus and the coccidophagous Nephus reunioni. BioControl 2013, 58, 351–357. [Google Scholar] [CrossRef]
- Raimundo, A.A.C.; Alves, M.L.L.G. Revisão dos Coccinelídeos de Portugal; Universidade de Évora: Évora, Portugal, 1986; p. 103. [Google Scholar]
- SPSS. Base Version 27.0.1.0; SPSS Inc.: Chicago, IL, USA, 2020. [Google Scholar]
- Rhamhalinghan, M. Intraspecific variation in ovariole number/ovary in Coccinella septempunctata L. (Coleoptera: Coccinellidae). Indian Zool. 1985, 9, 91–97. [Google Scholar]
- Sato, S.; Kushibuchi, K.; Yasuda, H. Effect of reflex bleeding of a predatory ladybird beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), as a means of avoiding intraguild predation and its cost. Appl. Entomol. Zool. 2009, 44, 203–206. [Google Scholar] [CrossRef][Green Version]
- Vargas, G.; Michaud, J.P.; Nechols, J.R. Trajectories of reproductive effort in Coleomegilla maculata and Hippodamia convergens (Coleoptera: Coccinellidae) respond to variation in both income and capital. Environ. Entomol. 2013, 42, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Hodek, H.; Evans, E.W. Food relationships. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., Honěk, A., van Emden, H.F., Eds.; Wiley-Blackwell Publishing: London, UK, 2012; pp. 141–274. [Google Scholar]
- Sebastião, D.; Borges, I.; Soares, A.O. Effect of temperature and prey in the biology of Scymnus subvillosus. BioControl 2015, 60, 241–249. [Google Scholar] [CrossRef]
- Beltrà, A.; Wäckers, F.L.; Nedvěd, O.; Pekas, A. Predation rate and performance of three ladybirds against the green peach aphid Myzus persicae in sweet pepper. Entomol. Exp. Appl. 2018, 66, 491–499. [Google Scholar] [CrossRef]
- Lucas, E.; Brodeur, J. A fox in sheep’s clothing: Furtive predators benefit from the communal defense of their prey. Ecology 2001, 82, 3246–3250. [Google Scholar] [CrossRef]
- Fréchette, B.; Larouche, F.; Lucas, E. Leucopis annulipes larvae (Diptera: Chamaemyiidae) use a furtive predation strategy within aphid colonies. Eur. J. Entomol. 2008, 105, 399–403. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacheco, P.; Borges, I.; Branco, B.; Lucas, E.; Soares, A.O. Costs and Benefits of Wax Production in the Larvae of the Ladybeetle Scymnus nubilus. Insects 2021, 12, 458. https://doi.org/10.3390/insects12050458
Pacheco P, Borges I, Branco B, Lucas E, Soares AO. Costs and Benefits of Wax Production in the Larvae of the Ladybeetle Scymnus nubilus. Insects. 2021; 12(5):458. https://doi.org/10.3390/insects12050458
Chicago/Turabian StylePacheco, Paulo, Isabel Borges, Beatriz Branco, Eric Lucas, and António Onofre Soares. 2021. "Costs and Benefits of Wax Production in the Larvae of the Ladybeetle Scymnus nubilus" Insects 12, no. 5: 458. https://doi.org/10.3390/insects12050458
APA StylePacheco, P., Borges, I., Branco, B., Lucas, E., & Soares, A. O. (2021). Costs and Benefits of Wax Production in the Larvae of the Ladybeetle Scymnus nubilus. Insects, 12(5), 458. https://doi.org/10.3390/insects12050458