
The microRNA miR-14 Regulates Egg-Laying by Targeting EcR in Honeybees (Apis mellifera)


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sampling
2.3. Oversupply/Inhibition of miR-14 and EcR in Honeybee Queens
2.4. RNA Isolation and Real Time Quantitative PCR
2.5. S2 Cell Culture, Luciferase Reporter Assay and Western-Blot
3. Results
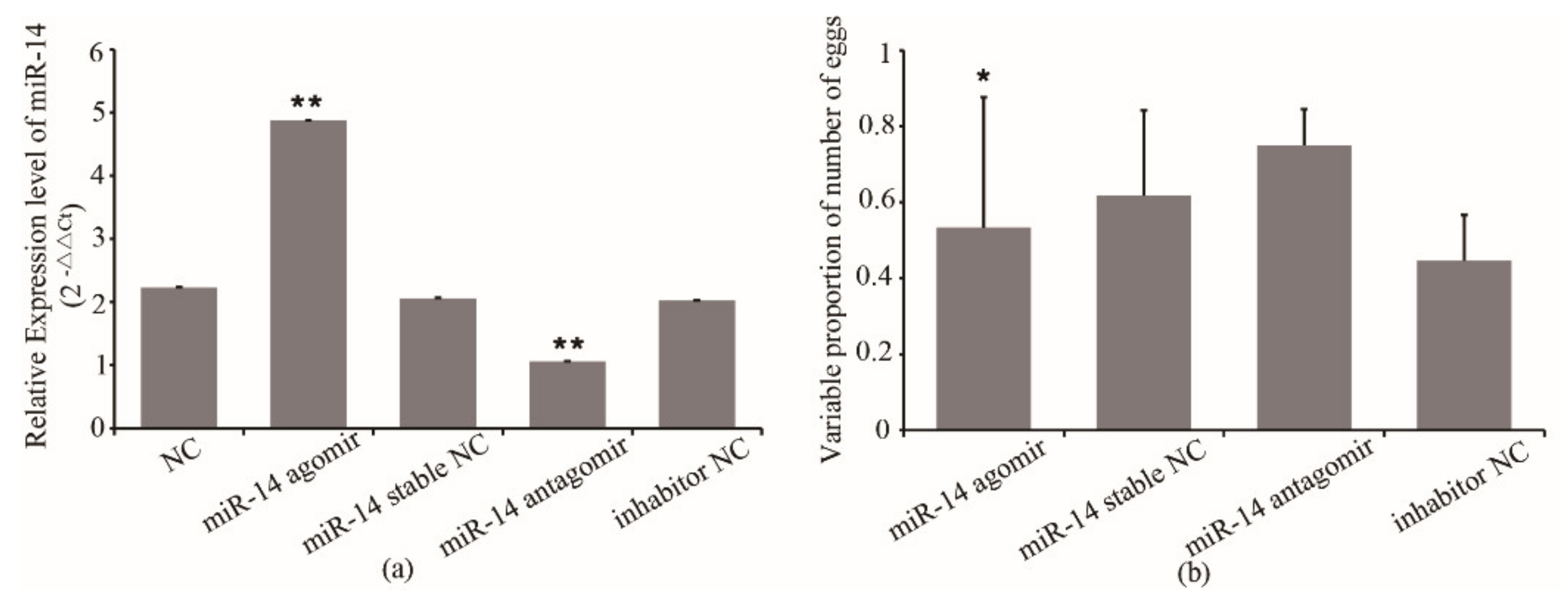
3.1. miR-14 Expression and Its Effect on Egg Number of Queens
3.2. EcR Expression and Its Effect on the Egg number of Queens
3.3. Confirmation of the Interaction of miR-14 with EcR Using a Luciferase Reporter Assay
3.4. The Regulation Mechanism of miR-14 on EcR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Woodward, D.R. Queen Bee: Biology, Rearing and Breeding; Northern Bee Books: Mytholmroyd, UK, 2010. [Google Scholar]
- Manfredini, F.; Brown, M.J.F.; Vergoz, V.; Oldroyd, B.P. RNA-sequencing elucidates the regulation of behavioural transitions associated with the mating process in honey bee queens. BMC Genom. 2015, 16, 563. [Google Scholar] [CrossRef]
- Mello, T.R.P.; Aleixo, A.C.; Pinheiro, D.G.; Nunes, F.M.F.; Bitondi, M.M.G.; Hartfelder, K.; Barchuk, A.R.; Simões, Z.L.P. Developmental regulation of ecdysone receptor (EcR) and EcR-controlled gene expression during pharate-adult development of honeybees (Apis mellifera). Front. Genet. 2014, 5, 445. [Google Scholar] [CrossRef]
- Kramarenko, A.S.; Lopukchov, A.A.; Gladyr, E.A.; Singina, G.N.; Ermilov, A.N.; Yanchukov, I.N.; Brem, G.; Zinovieva, N.A. 206 genome-wide associations for reproductive traits in Russian holstein population. Reprod. Fertil. Dev. 2014, 27, 194. [Google Scholar] [CrossRef]
- Spotter, A.; Gupta, P.; Nurnberg, G.; Reinsch, N.; Bienefeld, K. Development of a 44K SNP assay focussing on the analysis of a varroa-specific defence behaviour in honey bees (Apis mellifera carnica). Mol. Ecol. Resour. 2012, 12, 323–332. [Google Scholar] [CrossRef]
- Macedo, L.M.F.; Nunes, F.M.F.; Freitas, F.C.P.; Pires, C.V.; Tanaka, E.D.; Martins, J.R.; Piulachs, M.D.; Cristino, A.S.; Pinheiro, D.G.; Simões, Z.L.P. MicroRNA signatures characterizing caste-independent ovarian activity in queen and worker honeybees (Apis mellifera L.). Insect Mol. Biol. 2016, 25, 216–226. [Google Scholar] [CrossRef]
- Koywiwattrakul, P.; Sittipraneed, S. Expression of vitellogenin and transferrin in activated ovaries of worker honey bees, Apis mellifera. Biochem. Genet. 2009, 47, 19–26. [Google Scholar] [CrossRef]
- Thompson, G.J.; Yockey, H.; Lim, J.; Oldroyd, B.P. Experimental manipulation of ovary activation and gene expression in honey bee (Apis mellifera) queens and workers: Testing hypotheses of reproductive regulation. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2007, 307, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Vergoz, V.; Lim, J.; Oldroyd, B.P. Biogenic amine receptor gene expression in the ovarian tissue of the honey bee Apis mellifera. Insect Mol. Biol. 2012, 21, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Cardoen, D.; Wenseleers, T.; Ernst, U.R.; Danneels, E.L.; Laget, D.; Graaf, D.C.D.; Schoofs, L.; Verleyen, P. Genome-wide analysis of alternative reproductive phenotypes in honeybee workers. Mol. Ecol. 2011, 20, 4070–4084. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Zheng, H.; Corona, M.; Lu, Y.; Chen, X.; Cao, L.; Sohr, A.; Hu, F. Transcriptome comparison between inactivated and activated ovaries of the honey bee Apis mellifera L. Insect Mol. Biol. 2014, 23, 668–681. [Google Scholar] [CrossRef]
- Oxley, P.R.; Thompson, G.J.; Oldroyd, B.P. Four quantitative trait loci that influence worker sterility in the honeybee (Apis mellifera). Genetics 2008, 179, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.J.; Kucharski, R.; Maleszka, R.; Oldroyd, B.P. Towards a molecular definition of worker sterility: Differential gene expression and reproductive plasticity in honey bees. Insect Mol. Biol. 2006, 15, 537–644. [Google Scholar] [CrossRef]
- Pandey, A.; Bloch, G. Juvenile hormone and ecdysteroids as major regulators of brain and behavior in bees. Curr. Opin. Insect Sci. 2015, 12, 26–37. [Google Scholar] [CrossRef]
- Bloch, G.; Hefetz, A.; Hartfelder, K. Ecdysteroid titer, ovary status, and dominance in adult worker and queen bumble bees (Bombus terrestris). J. Insect Physiol. 2000, 46, 1033–1040. [Google Scholar] [CrossRef]
- Geva, S.; Hartfelder, K.; Bloch, G. Reproductive division of labor, dominance, and ecdysteroid levels in hemolymph and ovary of the bumble bee Bombus terrestris. J. Insect Physiol. 2005, 51, 811–823. [Google Scholar] [CrossRef]
- Gruntenko, N.E.; Rauschenbach, I.Y. Interplay of JH, 20E and biogenic amines under normal and stress conditions and its effect on reproduction. J. Insect Physiol. 2008, 54, 902–908. [Google Scholar] [CrossRef]
- Ashby, R.; Forêt, S.; Searle, I.; Maleszka, R. MicroRNAs in Honey Bee Caste Determination. Sci. Rep. 2016, 6, 18794. [Google Scholar] [CrossRef] [PubMed]
- Boecking, O.; Bienefeld, K.; Drescher, W. Heritability of the Varroa-specific hygienic behaviour in honey bees (Hymenoptera: Apidae). J. Anim. Breed. Genet. 2000, 117, 417–424. [Google Scholar] [CrossRef]
- Flatt, T.; Tu, M.P.; Tatar, M. Hormonal pleiotropy and the juvenile hormone regulation of Drosophila development and life history. Bioessays 2005, 27, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Hartfelder, K.; Emlen, D.J. 3.13–Endocrine Control of Insect Polyphenism. Compr. Mol. Insect Sci. 2005, 651–703. [Google Scholar] [CrossRef]
- Hoover, S.E.R.; Keeling, C.I.; Winston, M.L.; Slessor, K.N. The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften 2003, 90, 477–480. [Google Scholar] [CrossRef]
- Riddiford, L.M. Cellular and Molecular Actions of Juvenile Hormone I. General Considerations and Premetamorphic Actions. Adv. Insect Physiol. 1994, 24, 213–274. [Google Scholar]
- Wyatt, G.R.; Davey, K.G. Cellular and Molecular Actions of Juvenile Hormone. II. Roles of Juvenile Hormone in Adult Insects. Adv. Insect Physiol. 1996, 26, 1–155. [Google Scholar]
- Goodman, W.G.; Cusson, M. 8–The Juvenile Hormones; Plenum Press: New York, NY, USA, 2012; pp. 310–365. [Google Scholar]
- Chen, X.; Ma, C.; Chen, C.; Lu, Q.; Shi, W.; Liu, Z.; Wang, H.; Guo, H. Integration of lncRNA–miRNA–mRNA reveals novel insights into oviposition regulation in honey bees. PeerJ 2017, 5, e3881. [Google Scholar] [CrossRef] [PubMed]
- Varghese, J.; Cohen, S.M. microRNA miR-14 acts to modulate a positive autoregulatory loop controlling steroid hormone signaling in Drosophila. Genes Dev. 2007, 21, 2277–2282. [Google Scholar] [CrossRef]
- Hartfelder, K.; Bitondi, M.M.; Santana, W.C.; Simões, Z.L. Ecdysteroid titer and reproduction in queens and workers of the honey bee and of a stingless bee: Loss of ecdysteroid function at increasing levels of sociality? Insect Biochem. Mol. Biol. 2002, 32, 211–216. [Google Scholar] [CrossRef]
- Harbo, J.R. Propagation and Instrumental Insemination—Bee Genetics and Breeding—CHAPTER 15. Bee Genet. Breed. 1986, 1, 361–389. [Google Scholar]
- Amdam, G.V.; Simões, Z.L.P.; Guidugli, K.R.; Norberg, K.; Omholt, S.W. Disruption of vitellogenin gene function in adult honeybees by intra-abdominal injection of double-stranded RNA. BMC Biotechnol. 2003, 3, 1. [Google Scholar] [CrossRef]
- Tiscornia, G.; Singer, O.; Verma, I.M. Production and purification of lentiviral vectors. Nat. Protoc. 2006, 1, 241–245. [Google Scholar] [CrossRef]
- Hagedorn, H.H. The Role of Ecdysteroids in Reproduction. Endocrinology II; Pergamon Press: Oxford, UK, 1985; pp. 205–262. [Google Scholar]
- Nunes, F.M.; Ihle, K.E.; Mutti, N.S.; Simões, Z.L.; Amdam, G.V. The gene vitellogenin affects microRNA regulation in honey bee (Apis mellifera) fat body and brain. J. Exp. Biol. 2013, 216, 3724. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Page, R.E.; Fondrk, M.K.; Brent, C.S. Hormone response to bidirectional selection on social behavior. Evol. Dev. 2010, 12, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kaftanoglu, O.; Siegel, A.J.; Page, R.E.; Amdam, G.V. Surgically increased ovarian mass in the honey bee confirms link between reproductive physiology and worker behavior. J. Insect Physiol. 2010, 56, 1816–1824. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Kiuchi, M.; Takeuchi, H.; Kubo, T. Ecdysteroid biosynthesis in workers of the European honeybee Apis mellifera L. Insect Biochem. Mol. Biol. 2011, 41, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Bryant, B.; Macdonald, W.; Raikhel, A.S. microRNA miR-275 is indispensable for blood digestion and egg development in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2010, 107, 22391–22398. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Shi, T.; Yin, W.; Su, X.; Qi, L.; Huang, Z.Y.; Zhang, S.; Yu, L. The microRNA ame-miR-279a regulates sucrose responsiveness of forager honey bees (Apis mellifera). Insect Biochem. Mol. Biol. 2017, 90, 34–42. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | 5′ to 3′ |
|---|---|
| miR-14-F | GCGCTCAGTCTTTTTCTCT |
| U6 | CTTGCTTCGGCAGAACATAT |
| EcR-F | GCCTCCGGTTACCACTACAA |
| EcR-R | CTCGCAATTGTTCCCGTATT |
| β-actin-F | CTGCTGCATCATCCTCAAGC |
| β-actin-R | GAAAAGAGCCTCGGGACAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Fu, J. The microRNA miR-14 Regulates Egg-Laying by Targeting EcR in Honeybees (Apis mellifera). Insects 2021, 12, 351. https://doi.org/10.3390/insects12040351
Chen X, Fu J. The microRNA miR-14 Regulates Egg-Laying by Targeting EcR in Honeybees (Apis mellifera). Insects. 2021; 12(4):351. https://doi.org/10.3390/insects12040351
Chicago/Turabian StyleChen, Xiao, and Jinluan Fu. 2021. "The microRNA miR-14 Regulates Egg-Laying by Targeting EcR in Honeybees (Apis mellifera)" Insects 12, no. 4: 351. https://doi.org/10.3390/insects12040351
APA StyleChen, X., & Fu, J. (2021). The microRNA miR-14 Regulates Egg-Laying by Targeting EcR in Honeybees (Apis mellifera). Insects, 12(4), 351. https://doi.org/10.3390/insects12040351