
Small Bait Traps May Not Accurately Reflect the Composition of Necrophagous Diptera Associated to Remains


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling Procedure
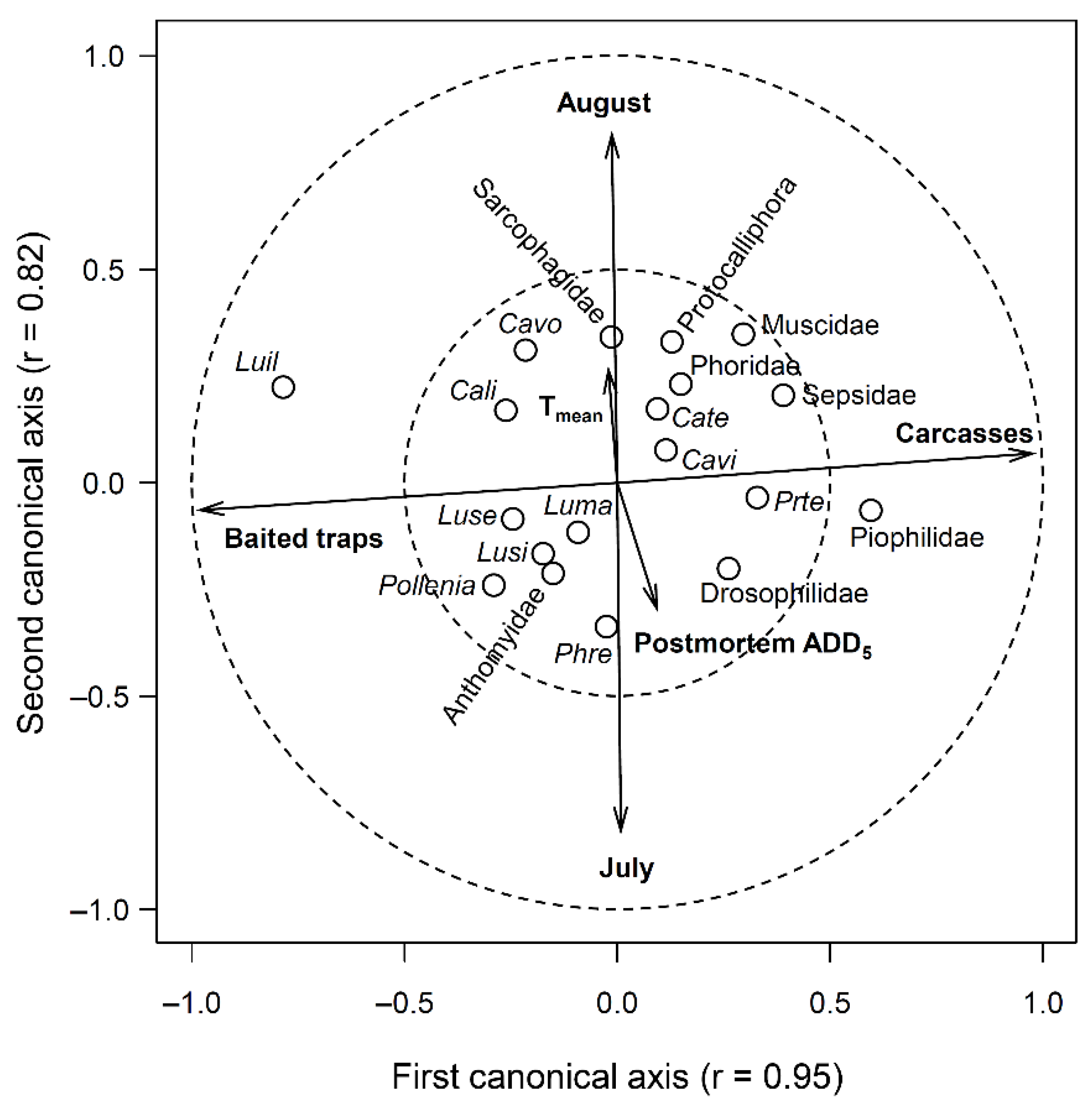
2.3. Statistical Analyses
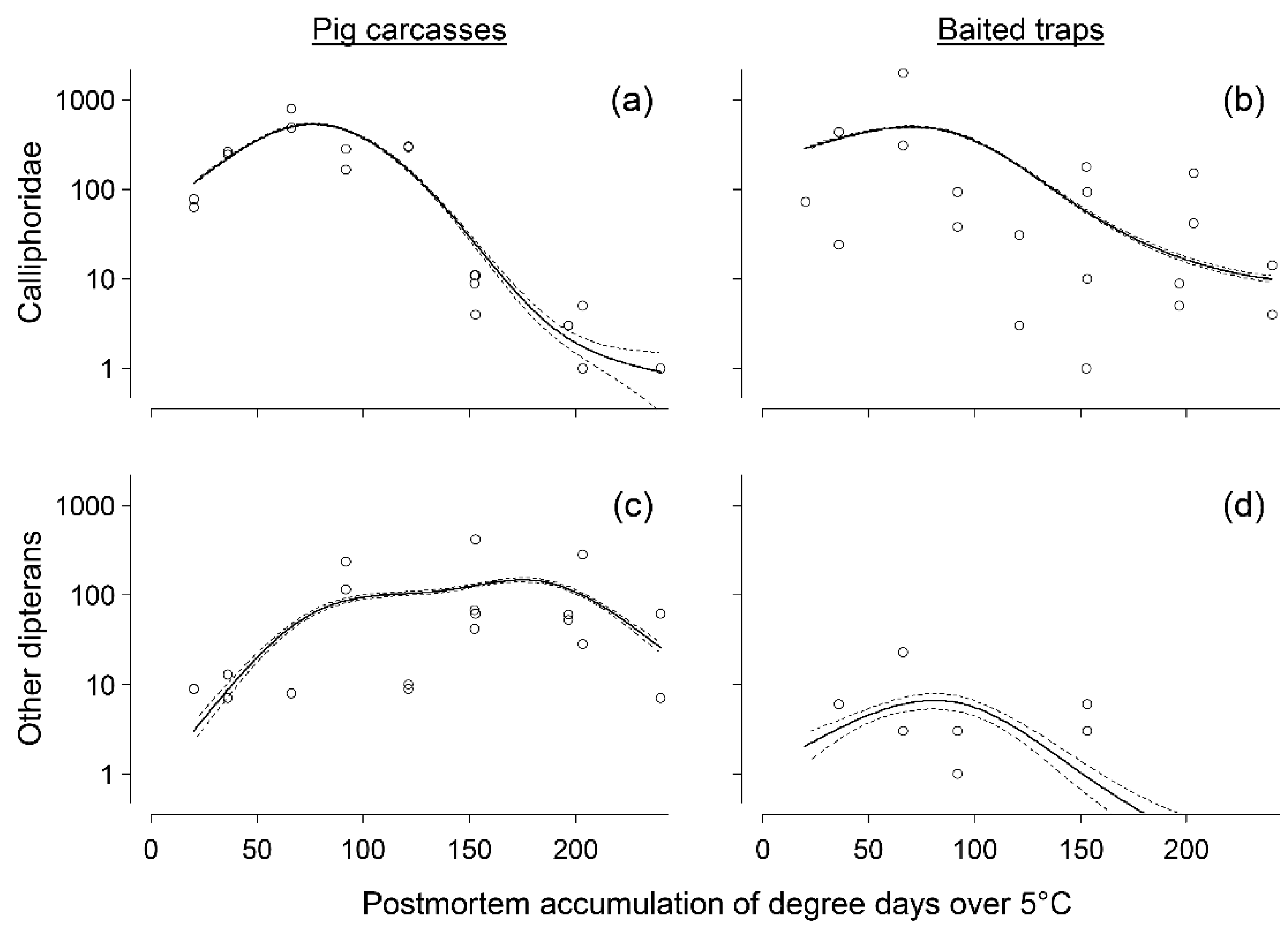
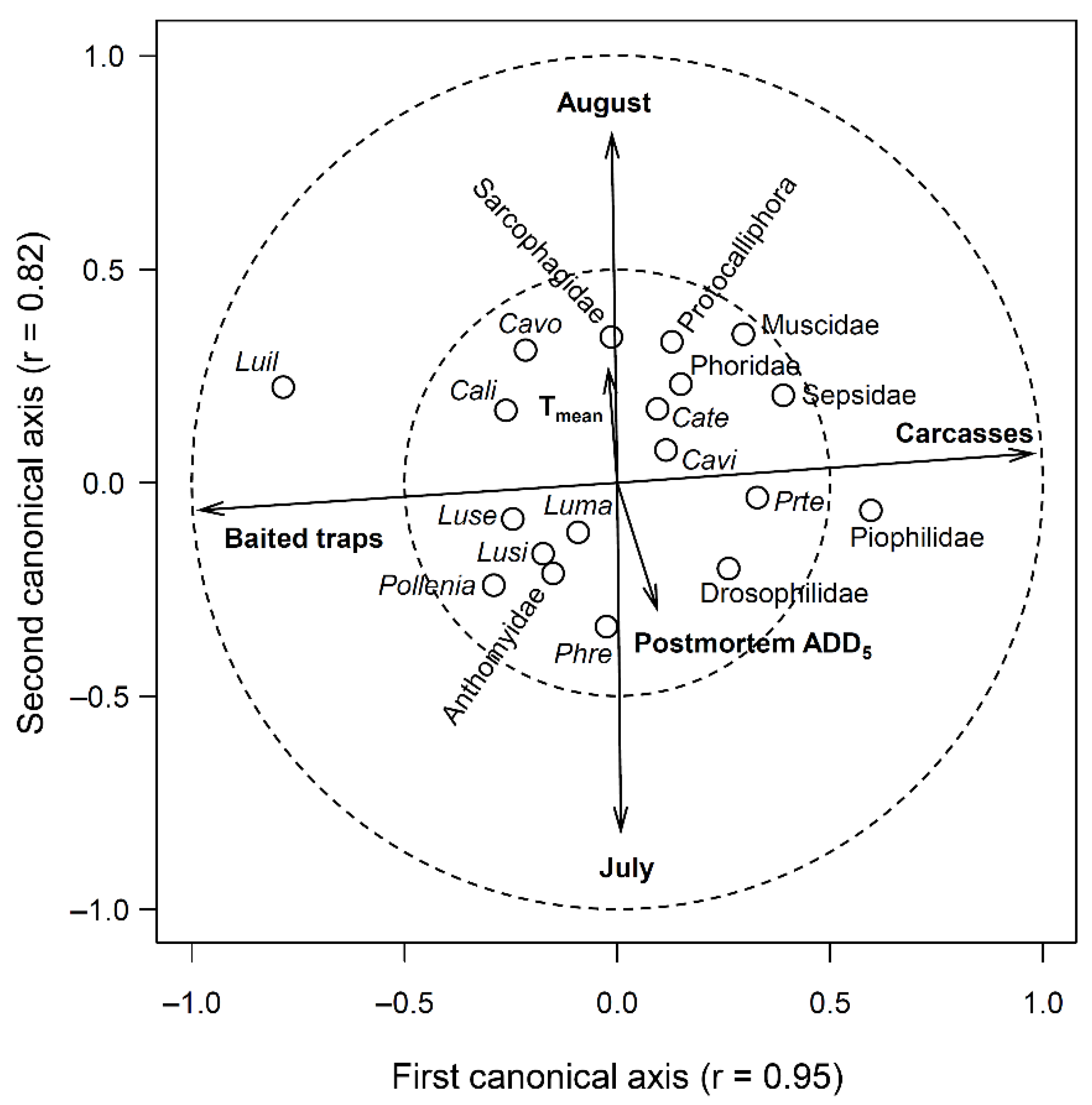
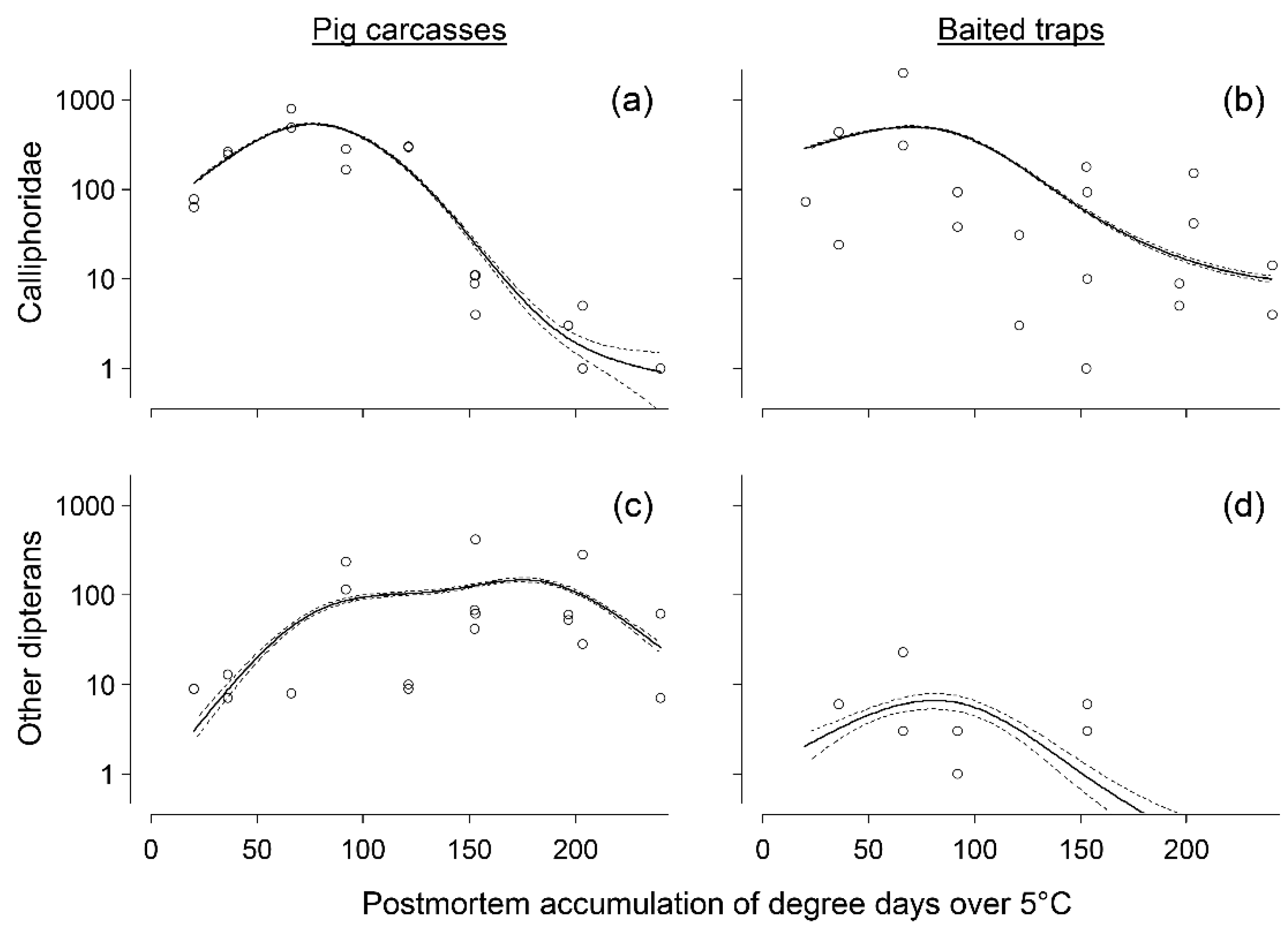
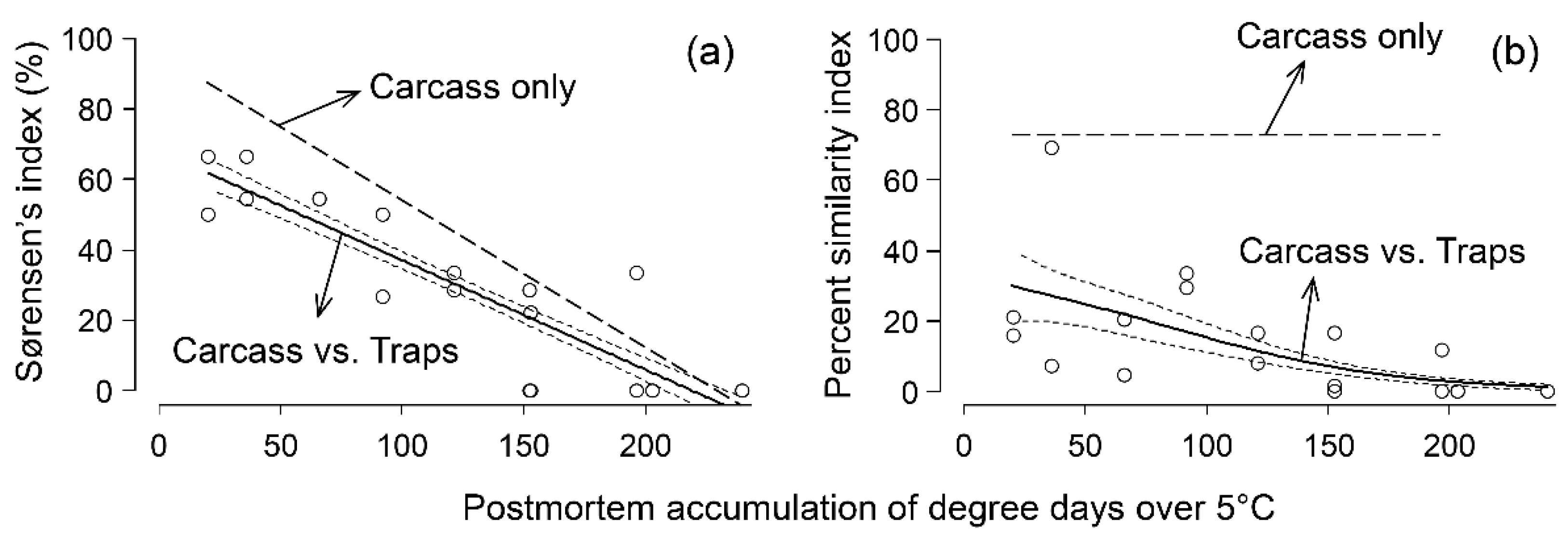
3. Results
3.1. Abundance Data
3.2. Comparison of Species Composition
4. Discussion
4.1. Discrepancies between Small Bait Traps and Remains
4.2. Other Considerations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farinha, A.; Dourado, C.G.; Centeio, N.; Oliveira, A.R.; Dias, D.; Rebelo, M.T. Small bait traps as accurate predictors of dipteran early colonizers in forensic studies. J. Insect Sci. 2014, 14, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, T.A.; Munger, L.; Thomas, L.; Wiggins, S.; Hildebrand, J.A. Estimating North Pacific right whale Eubalaena japonica density using passive acoustic cue counting. Endanger. Species Res. 2011, 13, 163–172. [Google Scholar] [CrossRef]
- Schoenly, K. Demographic bait trap. Environ. Entomol. 1981, 10, 615–617. [Google Scholar] [CrossRef]
- Wiener, J.G.; Smith, M.H. Relative efficiencies of four small mammal traps. J. Mammal. 1972, 53, 868–873. [Google Scholar] [CrossRef]
- Amendt, J.; Krettek, R.; Zehner, R. Forensic entomology. Naturwissenschaften 2004, 91, 51–65. [Google Scholar] [CrossRef]
- Benecke, M. A brief history of forensic entomology. Forensic Sci. Int. 2001, 120, 2–14. [Google Scholar] [CrossRef]
- Gennard, D. Forensic Entomology: An Introduction, 1st ed.; John Wiley & Sons: Chinchester, West Sussex, UK, 2012; p. 1. [Google Scholar]
- Hall, M.J.R. Trapping the flies that cause myiasis: Their responses to host-stimuli. Ann. Trop. Med. Parasitol. 1995, 89, 333–357. [Google Scholar] [CrossRef]
- Ordóñez, A.; García, M.D.; Fagua, G. Evaluation of efficiency of Schoenly trap for collecting adult sarcosaprophagous dipterans. J. Med. Entomol. 2008, 45, 522–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenly, K.G.; Haskell, N.H.; Hall, R.D.; Gbur, J.R. Comparative performance and complementarity of four sampling methods and arthropod preference tests from human and porcine remains at the Forensic Anthropology Center in Knoxville, Tennessee. J. Med. Entomol. 2007, 44, 881–894. [Google Scholar] [CrossRef]
- Hwang, C.; Turner, B.D. Spatial and temporal variability of necrophagous Diptera from urban to rural areas. Med. Vet. Entomol. 2005, 19, 379–391. [Google Scholar] [CrossRef]
- Baz, A.; Cifrián, B.; Díaz-äranda, L.M.; Martín-Vega, D. The distribution of adult blow-flies (Diptera: Calliphoridae) along an altitudinal gradient in Central Spain. Ann. Société Entomol. Fr. 2007, 43, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Aak, A.; Knudsen, G.K.; Soleng, A. Wind tunnel behavioural response and field trapping of the blowfly Calliphora vicina. Med. Vet. Entomol. 2010, 24, 250–257. [Google Scholar] [CrossRef]
- Payne, J.A. A Summer Carrion Study of the Baby Pig Sus Scrofa Linnaeus. Ecology 1965, 46, 592–602. [Google Scholar] [CrossRef]
- Brundage, A.; Bros, S.; Honda, J.Y. Seasonal and habitat abundance and distribution of some forensically important blow flies (Diptera: Calliphoridae) in Central California. Forensic Sci. Int. 2011, 212, 115–120. [Google Scholar] [CrossRef]
- Lutz, L.; Verhoff, M.A.; Amendt, J. Environmental factors influencing flight activity of forensically important female blow flies in Central Europe. Int. J. Legal Med. 2019, 133, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.R. Daily patterns of flight activity of blowflies (Calliphoridae: Diptera) in the Canberra district as indicated by trap catches. Aust. J. Zool. 1966, 14, 865–853. [Google Scholar] [CrossRef]
- Açikgöz, H.N.; Köse, S.K.; Divrak, D. Chemical meat bait traps versus basic meat bait traps in collection of adult flies. Entomol. Res. 2017, 47, 21–27. [Google Scholar] [CrossRef] [Green Version]
- George, K.A.; Archer, M.S.; Toop, T. Abiotic environmental factors influencing blowfly colonisation patterns in the field. Forensic Sci. Int. 2013, 229, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Weidner, L.M.; Gemmellaro, M.D.; Tomberlin, J.K.; Hamilton, G.C. Evaluation of bait traps as a means to predict initial blow fly (Diptera: Calliphoridae) communities associated with decomposing swine remains in New Jersey, USA. Forensic Sci. Int. 2017, 278, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Schoenly, K.; Reid, W. Community structure of carrion arthropods in the Chihuahuan Desert. J. Arid Environ. 1983, 6, 253–263. [Google Scholar] [CrossRef]
- Davies, L. Species composition and larval habitats of blowfly (Calliphoridae) populations in upland areas in England and Wales. Med. Vet. Entomol. 1990, 4, 61–68. [Google Scholar] [CrossRef]
- Anderson, G.; VanLaerhoven, S. Initial Studies on Insect Succession on Carrion in Southwestern British Columbia. J. Forensic Sci. 1996, 41, 617–625. [Google Scholar] [CrossRef]
- Matuszewski, S.; Hall, M.J.R.; Moreau, G.; Schoenly, K.G.; Tarone, A.M.; Villet, M.H. Pigs vs people: The use of pigs as analogues for humans in forensic entomology and taphonomy research. Int. J. Legal Med. 2020, 134, 793–810. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.J.; Benbow, M.E. When entomological evidence crawls away: Phormia regina en masse larval dispersal. J. Med. Entomol. 2011, 48, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.E.; Haskell, N.H.; Wells, J.D. Commonly used intercarcass distances appear to be sufficient to ensure independence of carrion insect succession pattern. Ann. Entomol. Soc. Am. 2016, 109, 72–80. [Google Scholar] [CrossRef]
- Michaud, J.-P.; Moreau, G. Facilitation may not be an adequate mechanism of community succession on carrion. Oecologia 2017, 183, 1143–1153. [Google Scholar] [CrossRef]
- Michaud, J.-P.; Moreau, G. Effect of variable rates of daily sampling of fly larvae on decomposition and carrion insect community assembly: Implications for forensic entomology field study protocols. J. Med. Entomol. 2013, 50, 890–897. [Google Scholar] [CrossRef] [PubMed]
- Marshall, S.A.; Whitworth, T.; Roscoe, L. Blow flies (Diptera: Calliphoridae) of eastern Canada with a key to Calliphoridae subfamilies and genera of eastern North America, and a key to the eastern Canadian species of Calliphorinae, Luciliinae and Chrysomyiinae. Can. J. Arthropod Identif. 2011, 11, 11. [Google Scholar] [CrossRef]
- Moreau, G. The pitfalls in the path of probabilistic inference in forensic entomology: A review. Insects 2021, 12, 240. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Oliveira, T.C.; Vasconcelos, S.D. Insects (Diptera) associated with cadavers at the Institute of Legal Medicine in Pernambuco, Brazil: Implications for forensic entomology. Forensic Sci. Int. 2010, 198, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Davies, L. Seasonal and spatial changes in blowfly production from small and large carcasses at Durham in lowland northeast England. Med. Vet. Entomol. 1999, 13, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Grassberger, M.; Friedrich, E.; Reiter, C. The blowfly Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae) as a new forensic indicator in Central Europe. Int. J. Legal Med. 2003, 117, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Baz, A.; Cifrián, B.; Martín-Vega, D. Patterns of diversity and abundance of carrion insect assemblages in the Natural Park ‘Hoces del Río Riaza’ (Central Spain). J. Insect Sci. 2014, 14, 162. [Google Scholar] [CrossRef] [Green Version]
- Goff, M.L. Comparison of insect species associated with decomposing remains recovered inside dwellings and outdoors on the Island of Oahu, Hawaii. J. Forensic Sci. 1991, 36, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Gruner, S.V.; Slone, D.H.; Capinera, J.L. Forensically important Calliphoridae (Diptera) associated with pig carrion in rural North-Central Florida. J. Med. Entomol. 2007, 44, 509–515. [Google Scholar] [CrossRef]
- Leccese, A. Insects as forensic indicators: Methodological aspects. Anil Aggrawals Internet J. Forensic Med. Toxicol. 2004, 5, 26–32. [Google Scholar]
- Martín-Vega, D.; Baz, A. Sex-biased captures of sarcosaprophagous Diptera in carrion-baited traps. J. Insect Sci. 2013, 13, 14. [Google Scholar] [CrossRef] [Green Version]
- Hayes, E.J.; Wall, R.; Smith, K.E. Mortality rate, reproductive output, and trap response bias in populations of the blowfly Lucilia sericata. Ecol. Entomol. 1999, 24, 300–307. [Google Scholar] [CrossRef]
- Sontigun, N.; Sukontason, K.L.; Klong-Klaew, T.; Sanit, S.; Samerjai, C.; Somboon, P.; Thanapornpoonpong, S.; Amendt, J.; Sukontason, K. Bionomics of the oriental latrine fly Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae): Temporal fluctuation and reproductive potential. Parasites Vectors 2018, 11, 415. [Google Scholar] [CrossRef]
- Nuorteva, P. The flying activity of Phormia terrae-novae R.-D. (Dipt, Calliphoridae) in subarctic conditions. Ann. Zool. Fenn. 1966, 3, 73–81. [Google Scholar]
- Prinkkilá, M.-L.; Hanski, I. Complex competitive interactions in four species of Lucilia blowflies. Ecol. Entomol. 1995, 20, 261–272. [Google Scholar] [CrossRef]
- Mashaly, A.; Mahmoud, A.; Ebaid, H.; Sammour, R. Effect of height to ground level on the insect attraction to exposed rabbit carcasses. J. Environ. Biol. 2020, 41, 73–78. [Google Scholar] [CrossRef]
- Kirk, A.A. The effect of the dung pad fauna on the emergence of Musca tempestiva [Dipt: Muscidae] from dung pads in southern France. Entomophaga 1992, 37, 507–514. [Google Scholar] [CrossRef]
- Vogt, W.G. Trap catches of Musca vetustissima Walker (Diptera: Muscidae) and other arthropods associated with cattle dung in relation to height above ground level. Austral Entomol. 1988, 27, 143–147. [Google Scholar] [CrossRef]
- Prado e Castro, C.; Cunha, E.; Serrano, A.; García, M.D. Piophila megastigmata (Diptera: Piophilidae): First records on human corpses. Forensic Sci. Int. 2012, 214, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Rochefort, S.; Wheeler, T. Diversity of Piophilidae (Diptera) in northern Canada and description of a new Holarctic species of Parapiophila McAlpine. Zootaxa 2015, 3925, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Micozzi, M.S. Experimental study of postmortem change under field conditions: Effects of freezing, thawing, and mechanical injury. J. Forensic Sci. 1986, 31, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Bugajski, K.N.; Seddon, C.C.; Williams, R.E. A comparison of blow fly (Diptera: Calliphoridae) and beetle (Coleoptera) activity on refrigerated only versus frozen-thawed pig carcasses in Indiana. J. Med. Entomol. 2011, 48, 1231–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepburn, G.A. Sheep blowfly research I—A survey of maggot collections from live sheep and a note on the trapping of blowflies. Onderstepoort J. Vet. Anim. Ind. 1943, 18, 13–18. [Google Scholar]
- Lobo, J.M.; Hortal, J.; Cabrero-Sañudo, F.J. Regional and local influence of grazing activity on the diversity of a semi-arid dung beetle community. Divers. Distrib. 2006, 12, 111–123. [Google Scholar] [CrossRef]
- Macedo, R.; Audino, L.D.; Korasaki, V.; Louzada, J. Conversion of Cerrado savannas into exotic pastures: The relative importance of vegetation and food resources for dung beetle assemblages. Agric. Ecosyst. Environ. 2020, 288, 106709. [Google Scholar] [CrossRef]
- Müller, J.; Wende, B.; Strobl, C.; Eugster, M.; Gallenberger, I.; Floren, A.; Steffan-Dewenter, I.; Linsenmair, K.E.; Weisser, W.W.; Gossner, M.M. Forest management and regional tree composition drive the host preference of saproxylic beetle communities. J. Appl. Ecol. 2015, 52, 753–762. [Google Scholar] [CrossRef] [Green Version]
- Thibault, M.; Moreau, G. Enhancing bark- and wood-boring beetle colonization and survival in vertical deadwood during thinning entries. J. Insect Conserv. 2016, 20, 789–796. [Google Scholar] [CrossRef]
- Zito, P.; Guarino, S.; Peri, E.; Sajeva, M.; Colazza, S. Electrophysiological and behavioural responses of the housefly to “sweet” volatiles of the flowers of Caralluma europaea (Guss.) NE Br. Arthropod-Plant Int. 2013, 7, 485–489. [Google Scholar] [CrossRef]
- Hoffman, E.M.; Curran, A.M.; Dulgerian, N.; Stockham, R.A.; Eckenrode, B.A. Characterization of the volatile organic compounds present in the headspace of decomposing human remains. Forensic Sci. Int. 2009, 186, 6–13. [Google Scholar] [CrossRef]
- Arnaldos, M.I.; García, M.D.; Romera, E.; Presa, J.J.; Luna, A. Estimation of postmortem interval in real cases based on experimentally obtained entomological evidence. Forensic Sci. Int. 2005, 149, 57–65. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Experimental Unit | Carcasses | Traps | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Month | July | August | Average | July | August | Average | ||||||||||||||||||
| Postmortem Interval (PMI) | 0–2 | 3–5 | 6–8 | 9–11 | 12–14 | 0–2 | 3–5 | 6–8 | 9–11 | 12–14 | 0–2 | 0–14 | 0–2 | 3–5 | 6–8 | 9–11 | 12–14 | 0–2 | 3–5 | 6–8 | 9–11 | 12–14 | 0–2 | 0–14 |
| Lucilia illustris (Meigen) | 35.7 | 13.1 | ‒ | 5.0 | ‒ | 36.0 | ‒ | ‒ | ‒ | ‒ | 35.9 | 9.0 | 87.7 | 42.4 | 74.3 | 75.7 | 57.1 | 89.8 | 52.9 | 57.8 | 94.3 | 88.9 | 88.8 | 72.1 |
| Phormia regina (Meigen) | 14.3 | 81.0 | 84.6 | 10.0 | 7.1 | 22.0 | 34.8 | 2.4 | ‒ | ‒ | 18.1 | 25.6 | 1.4 | 54.2 | 14.3 | 16.4 | 42.9 | 5.9 | 37.0 | 28.4 | 4.2 | 5.6 | 3.7 | 21.0 |
| Pollenia sp. Robineau-Desvoidy | 14.3 | ‒ | 2.6 | 5.0 | 7.1 | 2.0 | 1.1 | ‒ | ‒ | ‒ | 8.1 | 3.2 | 11.0 | 2.0 | 5.7 | 7.3 | ‒ | 2.5 | 2.2 | 3.4 | 1.0 | 5.6 | 6.8 | 4.1 |
| Protophormia terraenovae (Robineau-Desvoidy) | ‒ | 2.4 | 5.1 | ‒ | ‒ | 2.0 | 2.2 | ‒ | ‒ | ‒ | 1.0 | 1.2 | ‒ | 0.2 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.0 |
| Calliphora terraenovae Macquart | 7.1 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 3.6 | 0.7 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 1.4 | ‒ | ‒ | ‒ | ‒ | 0.1 |
| Calliphora vicina Robineau-Desvoidy | 3.6 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 1.8 | 0.4 | ‒ | 0.0 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.0 |
| Lucilia sericata (Meigen) | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 2.9 | ‒ | ‒ | ‒ | 0.7 | ‒ | ‒ | ‒ | ‒ | 0.4 |
| Calliphora livida Hall | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.0 | ‒ | ‒ | ‒ | ‒ | 1.4 | ‒ | 0.5 | ‒ | ‒ | 0.2 |
| Protocalliphora sp. Hough | ‒ | ‒ | ‒ | ‒ | ‒ | 2.0 | ‒ | ‒ | ‒ | ‒ | 1.0 | 0.2 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
| Lucilia silvarum (Meigen) | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.6 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.1 |
| Calliphora vomitoria (Linnaeus) | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.2 | ‒ | ‒ | ‒ | ‒ | 0.1 | 0.0 |
| Lucilia magnicornis (Siebke) | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.0 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 0.0 |
| Piophilidae—Stearibia nigriceps (Meigen) | 3.6 | 3.6 | ‒ | 60.0 | 78.6 | 14.0 | 33.7 | 92.9 | 100.0 | 100.0 | 8.8 | 48.6 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
| Muscidae | 10.7 | ‒ | 2.6 | 5.0 | 7.1 | 16.0 | 7.9 | ‒ | ‒ | ‒ | 13.4 | 4.9 | ‒ | 1.1 | ‒ | ‒ | ‒ | 0.8 | 2.2 | 4.3 | ‒ | ‒ | 0.4 | 0.8 |
| Sepsidae | 3.6 | ‒ | 2.6 | 10.0 | ‒ | 2.0 | 18.0 | 4.8 | ‒ | ‒ | 2.8 | 4.1 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
| Sarcophagidae | 3.6 | ‒ | ‒ | ‒ | ‒ | 4.0 | ‒ | ‒ | ‒ | ‒ | 3.8 | 0.8 | ‒ | 0.0 | ‒ | ‒ | ‒ | 0.4 | 0.7 | 3.4 | ‒ | ‒ | 0.2 | 0.5 |
| Anthomyiidae | 3.6 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 1.8 | 0.4 | ‒ | ‒ | 2.9 | ‒ | ‒ | ‒ | 1.4 | 2.6 | ‒ | ‒ | ‒ | 0.7 |
| Drosophilidae | ‒ | ‒ | 2.6 | 5.0 | ‒ | ‒ | 1.1 | ‒ | ‒ | ‒ | ‒ | 0.9 | ‒ | ‒ | ‒ | ‒ | ‒ | 0.2 | ‒ | ‒ | ‒ | ‒ | 0.1 | 0.0 |
| Phoridae | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | 1.1 | ‒ | ‒ | ‒ | ‒ | 0.1 | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ | ‒ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
LeBlanc, K.; Boudreau, D.R.; Moreau, G. Small Bait Traps May Not Accurately Reflect the Composition of Necrophagous Diptera Associated to Remains. Insects 2021, 12, 261. https://doi.org/10.3390/insects12030261
LeBlanc K, Boudreau DR, Moreau G. Small Bait Traps May Not Accurately Reflect the Composition of Necrophagous Diptera Associated to Remains. Insects. 2021; 12(3):261. https://doi.org/10.3390/insects12030261
Chicago/Turabian StyleLeBlanc, Kathleen, Denis R. Boudreau, and Gaétan Moreau. 2021. "Small Bait Traps May Not Accurately Reflect the Composition of Necrophagous Diptera Associated to Remains" Insects 12, no. 3: 261. https://doi.org/10.3390/insects12030261
APA StyleLeBlanc, K., Boudreau, D. R., & Moreau, G. (2021). Small Bait Traps May Not Accurately Reflect the Composition of Necrophagous Diptera Associated to Remains. Insects, 12(3), 261. https://doi.org/10.3390/insects12030261