
Mapping Invasion Potential of the Pest from Central Asia, Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae), in the Shelter Forests of Northwest China

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
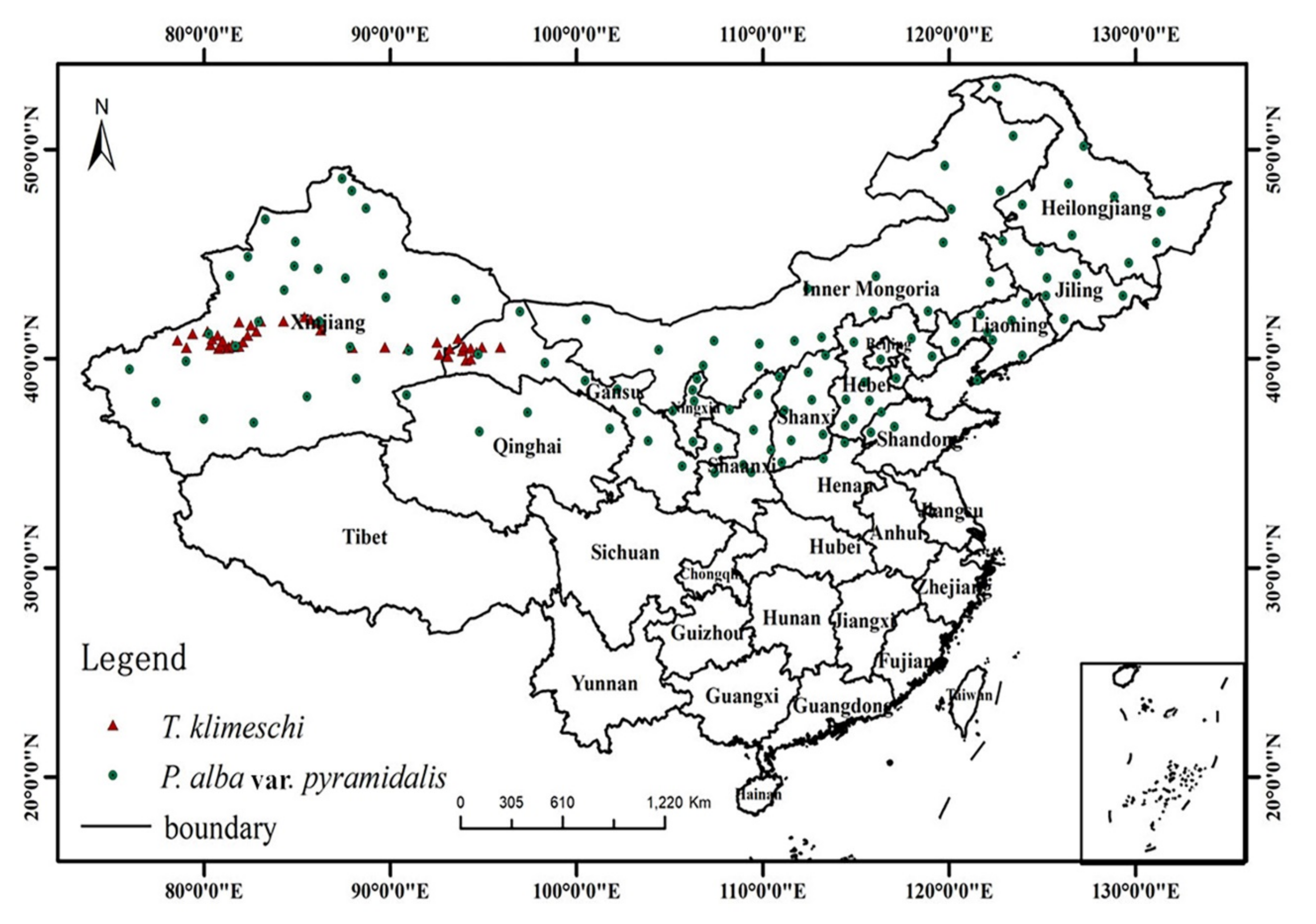
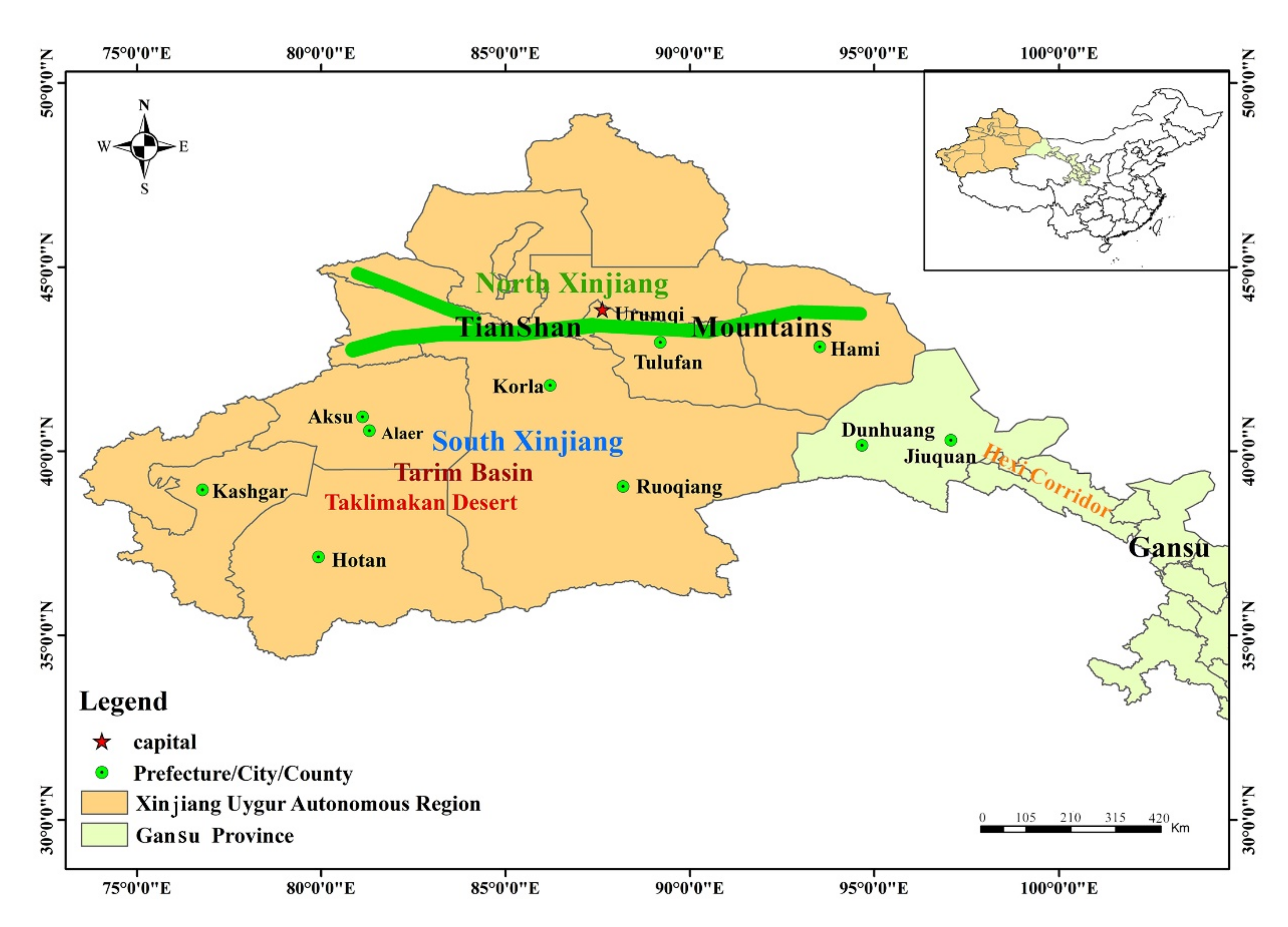
2.1. Study Area
2.2. Occurrence Collection
2.3. Environmental Variables
2.4. Species Distribution Modeling
2.5. The Centroid Changes Using SDM Tool-Box
3. Results
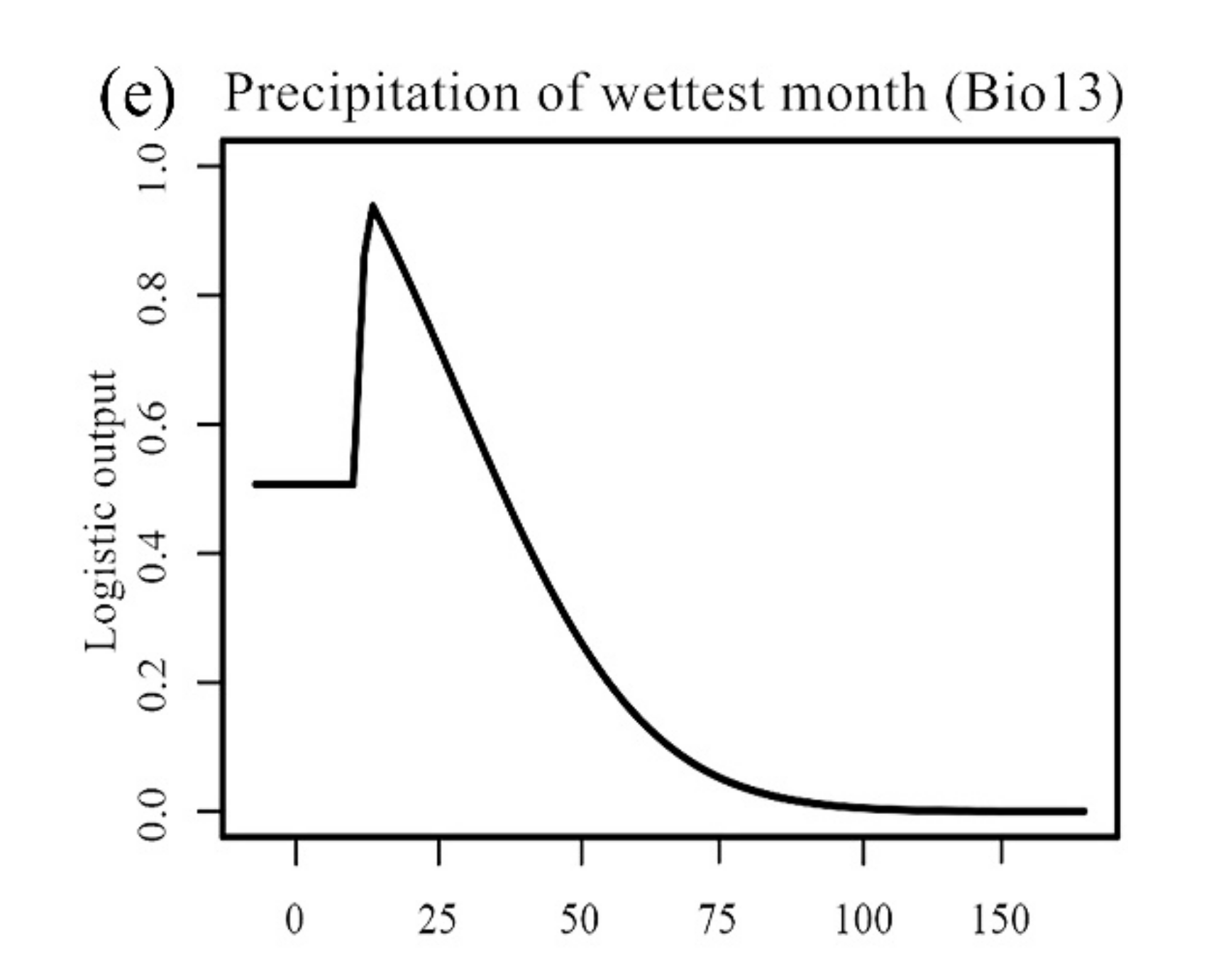
3.1. Model Performance and Variables’ Response Curves
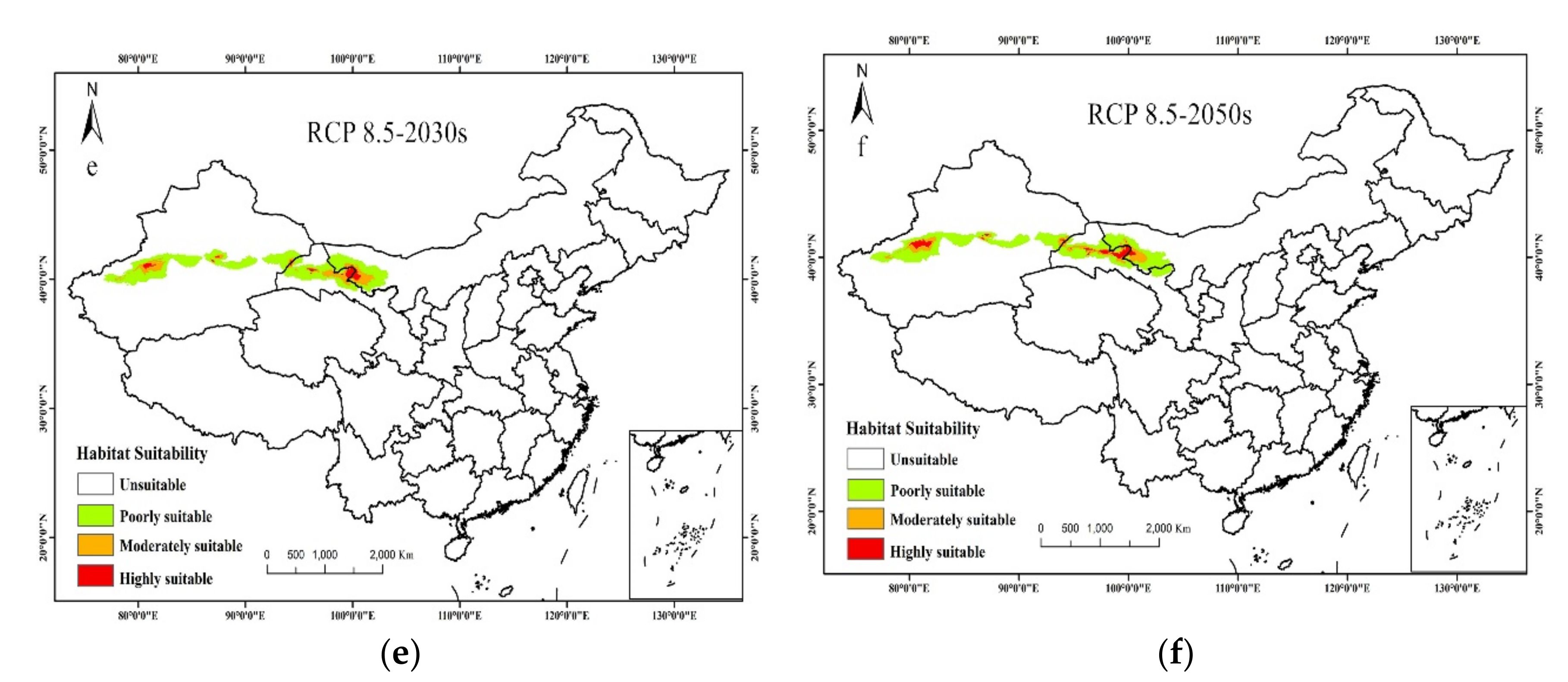
3.2. Current Potential Distribution of T. Klimeschi
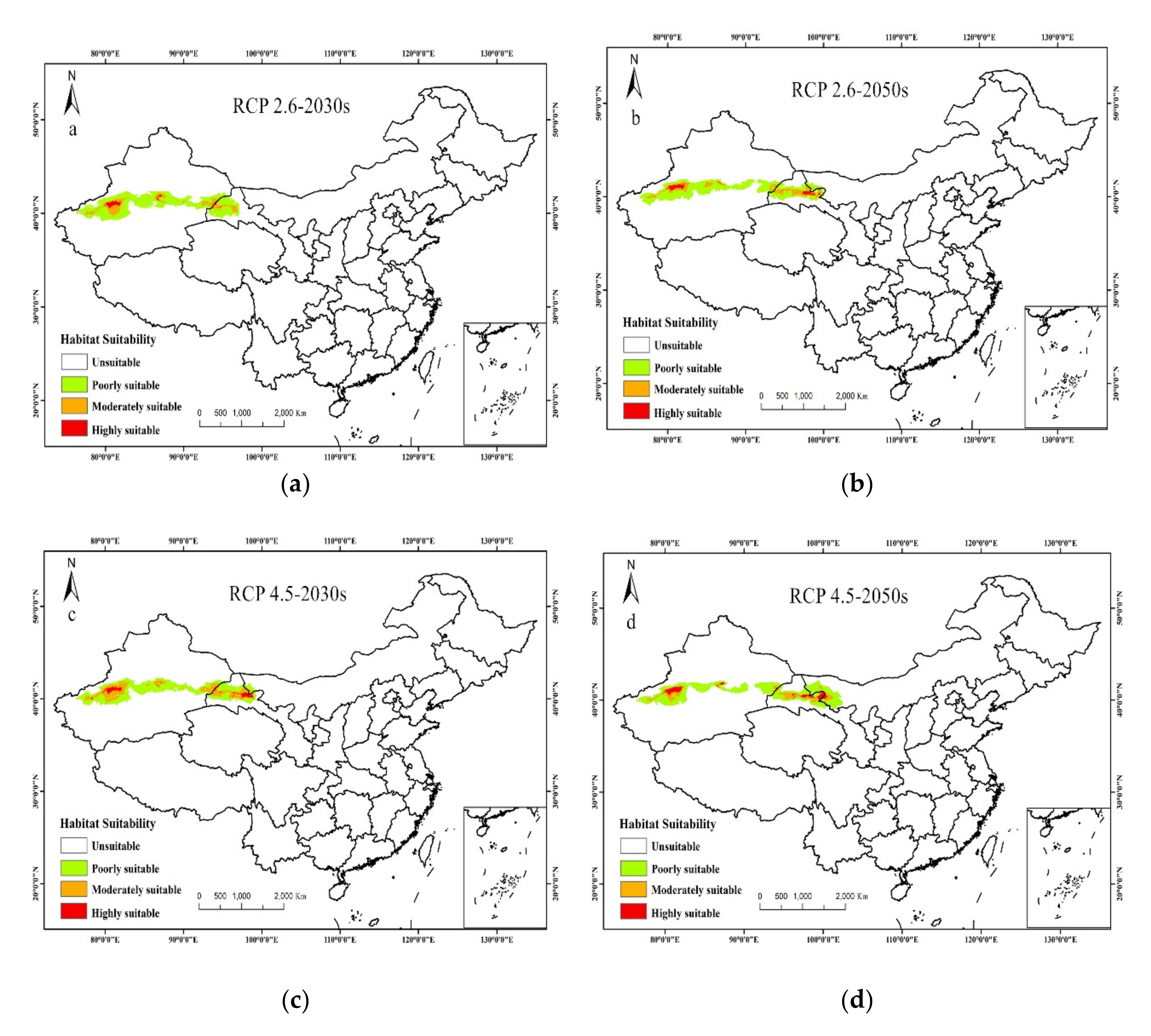
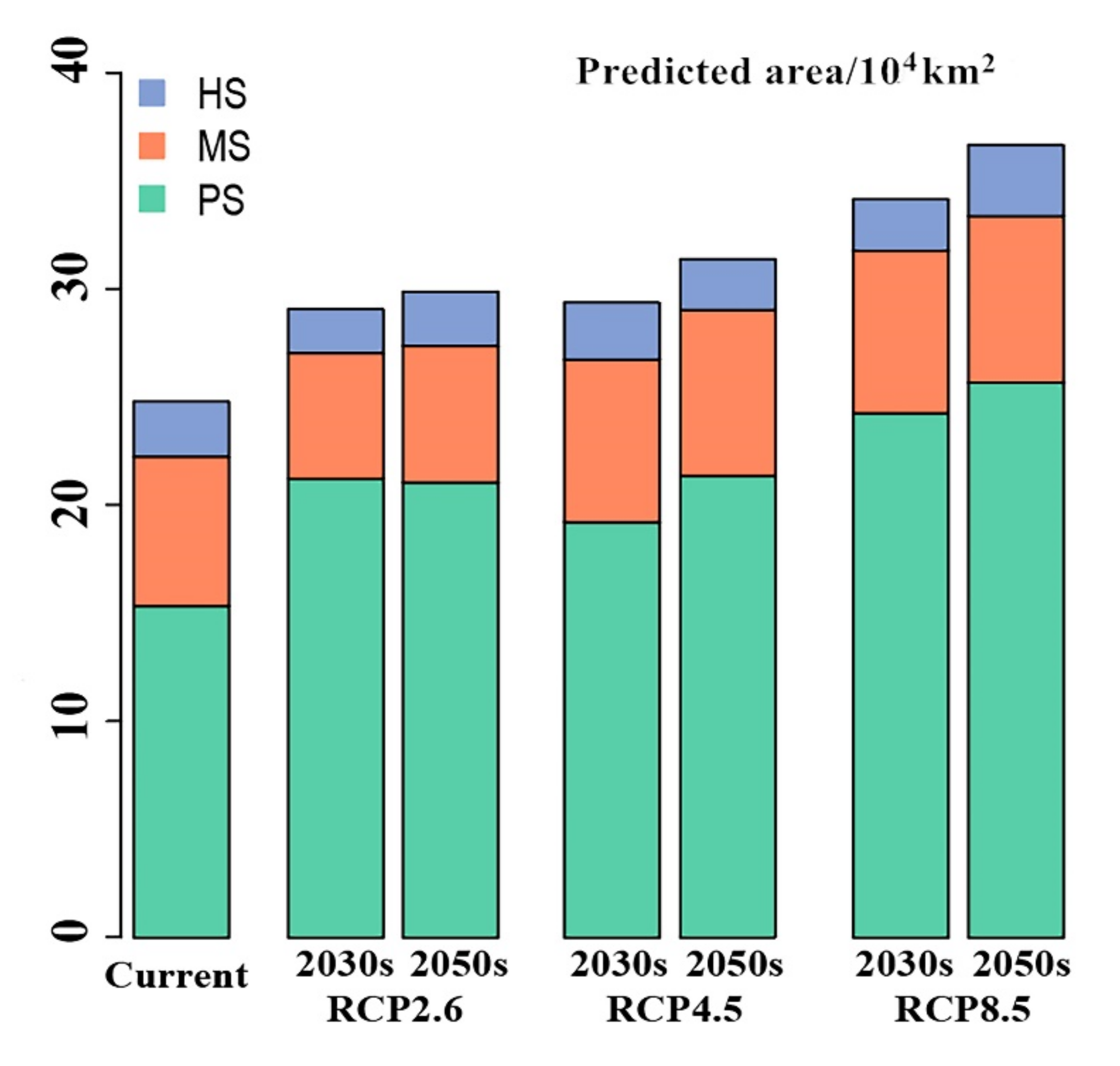
3.3. Future Potential Distribution of T. Klimeschi
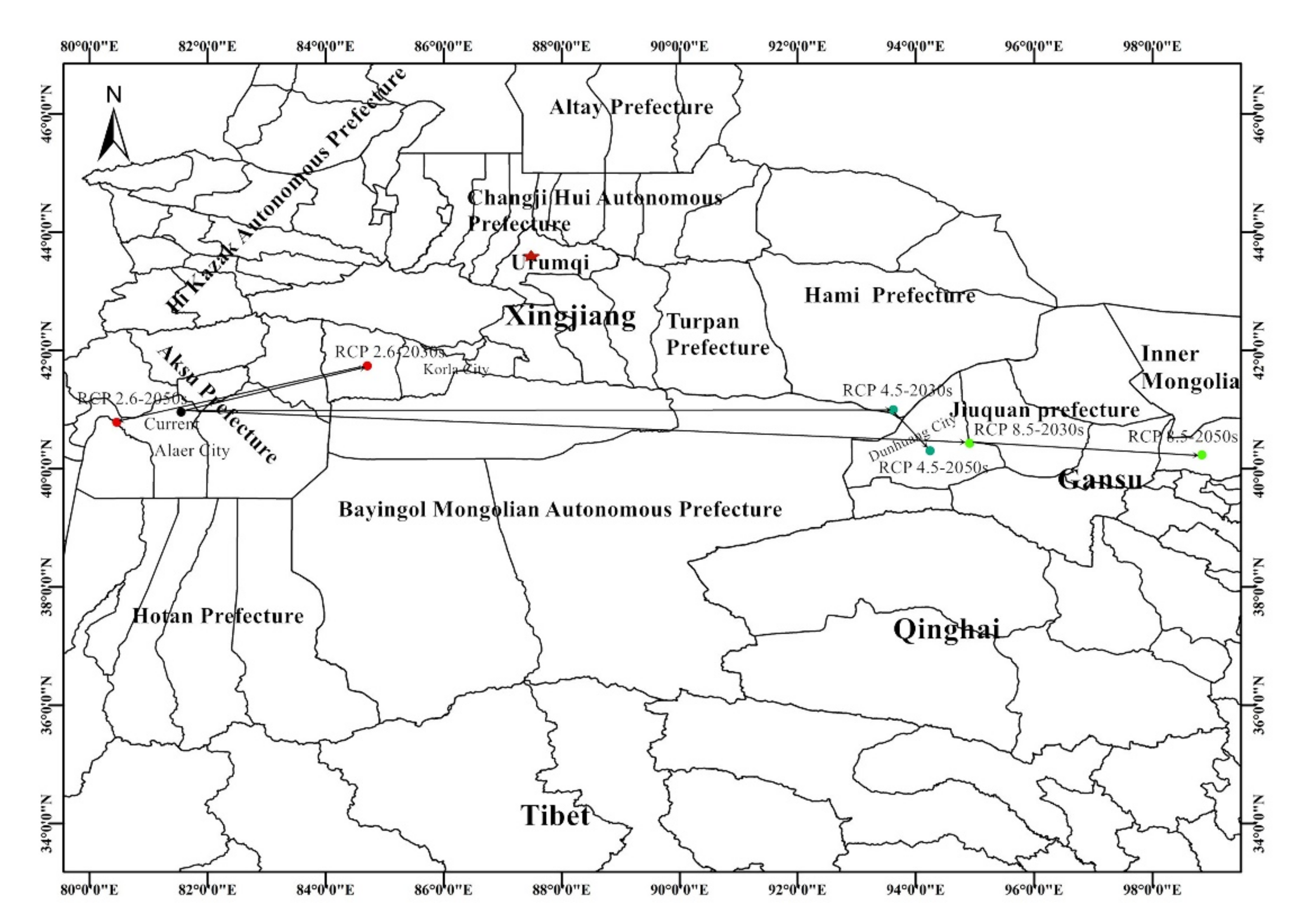
3.4. The Distributional Centroid Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, Y.; Luo, Z.B.; Wang, S.S.; Zhang, P. A new pest that harms Populus alba var. pyramidalis-Trypophloeus klimeschi. J. Tarim. Univers. Agric. 2003, 15, 9–10. [Google Scholar]
- Eggers, V.O.H. Trypophloeus klimeschi nov. spec. Entomol. Blätter 1915, 25, 7–9. [Google Scholar]
- Cao, Y.; Luo, Z.B.; Wang, S.S.; Wang, X.H.; Zhang, P. Investigation on the death of Populus alba var. pyramidalis and its cause. For. Pest Dis. 2003, 22, 12–13. [Google Scholar]
- Gao, G.Q.; Dai, L.L.; Gao, J.; Wang, J.X.; Chen, H. Volatile organic compound analysis of host and non-host poplars for Try-pophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae). Russ. J. Plant Physiol. 2018, 65, 916–925. [Google Scholar] [CrossRef]
- Gao, G.; Gao, J.; Hao, C.; Dai, L.; Chen, H. Biodiversity and Activity of Gut Fungal Communities across the Life History of Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae). Int. J. Mol. Sci. 2018, 19, 2010. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.Q.; Dai, L.L.; Gao, J.; Wang, J.X.; Chen, H. Electroantennogram, behavioural responses, and field trapping of Try-pophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae) to eight host volatiles. Can. Entomol. 2019, 151, 1–15. [Google Scholar] [CrossRef]
- Luo, W.X.; Liu, G.Q.; Li, J.J. Cultivation Technology of Main Tree Species in Northwest China; China Forestry Press: Beijing, China, 2007; pp. 679–681. [Google Scholar]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nat. Cell Biol. 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Ge, F. Challenges facing entomologists in a changing global climate. Chin. Bull. Entomol. 2011, 48, 1–1122. [Google Scholar]
- Zhang, H.; Lin, J.T.; Zhong, K. Responses of insects to global warming. J. Environ. Entomol. 2015, 37, 1280–1286. [Google Scholar]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Heads, M. The relationship between biogeography and ecology: Envelopes, models, predictions. Biol. J. Linn. Soc. 2015, 115, 456–468. [Google Scholar] [CrossRef]
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. bioclim: The first species distribution modelling package, its early appli-cations and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Stockwell, D.; Peters, D.P. The GARP modelling system: Problems and solutions to automated spatial prediction. Int. J. Geogr. Inf. Sci. 1999, 13, 143–158. [Google Scholar] [CrossRef]
- Hirzel, A.; Guisan, A. Which is the optimal sampling strategy for habitat suitability modelling. Ecol. Model. 2002, 157, 331–341. [Google Scholar] [CrossRef]
- Carpenter, G.; Gillison, A.N.; Winter, J. DOMAIN: A flexible modelling procedure for mapping potential distributions of plants and animals. Biodivers. Conserv. 1993, 2, 667–680. [Google Scholar] [CrossRef]
- Yee, T.W.; Mitchell, N.D. Generalized additive models in plant ecology. J. Veg. Sci. 1991, 2, 587–602. [Google Scholar] [CrossRef]
- Lehmann, A.; Overton, J.M.; Leathwick, J.R. GRASP: Generalized regression analysis and spatial prediction. Ecol. Model. 2002, 160, 189–207. [Google Scholar] [CrossRef]
- Kriticos, D.J.; De Barro, P.J.; Yonow, T.; Ota, N.; Sutherst, R.W. The potential geographical distribution and phenology of Bemisia tabaci Middle East/Asia Minor 1, considering irrigation and glasshouse production. Bull. Èntomol. Res. 2020, 110, 567–576. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Li, G.Q.; Liu, C.C.; Liu, Y.G.; Yang, J.; Zhang, X.S.; Guo, K. Advances in theoretical issues of species distribution models. Acta Ecol. Sin. 2013, 33, 4827–4835. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occur-rence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Domrös, M.; Gongbing, P. Climate Zones of China. In The Climate of China; Springer International Publishing: Berlin/Heidelberg, Germany, 1988; pp. 258–280. [Google Scholar]
- IPCC. Climate Change 2014: Fifth Assessment Synthesis Report. Available online: http://www.ipcc/report/ar5/syr/ (accessed on 13 April 2015).
- Van Vliet, J.; Eitelberg, D.A.; Verburg, P.H. A global analysis of land take in cropland areas and production displacement from urbanization. Glob. Environ. Chang. 2017, 43, 107–115. [Google Scholar] [CrossRef]
- Sánchez-Lugo, A.; Berrisford, P.; Morice, C.; Argüez, A. Temperature [in State of the Climate in 2018]. Bull. Am. Meteorol. Soc. 2018, 99, S11–S12. [Google Scholar]
- Xin, X.; Gao, F.; Wei, M.; Wu, T.; Fang, Y.; Zhang, J. Decadal prediction skill of BCC-CSM1.1 climate model in East Asia. Int. J. Clim. 2017, 38, 584–592. [Google Scholar] [CrossRef]
- Duque-Lazo, J.; Van Gils, H.; Groen, T.; Navarro-Cerrillo, R. Transferability of species distribution models: The case of Phytophthora cinnamomi in Southwest Spain and Southwest Australia. Ecol. Model. 2016, 320, 62–70. [Google Scholar] [CrossRef]
- Tibshirani, R. Regression Shrinkage and Selection Via the Lasso. J. R. Stat. Soc. Ser. B Stat. Methodol. 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Kukreja, S.L.; Löfberg, J.; Brenner, M.J. A least absolute shrinkage and selection operator (LASSO) for nonlinear system iden-tification. IPV 2006, 39, 814–819. [Google Scholar]
- Duque-Lazo, J.; Navarro-Cerrillo, R. What to save, the host or the pest? The spatial distribution of xylophage insects within the Mediterranean oak woodlands of Southwestern Spain. For. Ecol. Manag. 2017, 392, 90–104. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Zhu, G.P.; Liu, Q.; Gao, Y.B. Improving ecological niche model transferability to predict the potential distribution of invasive exotic species. Biodivers. Sci. 2014, 22, 223–230. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Bean, W.T.; Stafford, R.; Brashares, J.S. The effects of small sample size and sample bias on threshold selection and accuracy assessment of species distribution models. Ecography 2012, 35, 250–258. [Google Scholar] [CrossRef]
- Brown, J.L. SDM toolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Régnière, J.; Powell, J.; Bentz, B.; Nealis, V. Effects of temperature on development, survival and reproduction of insects: Ex-perimental design, data analysis and modeling. J. Insect Physiol. 2012, 58, 634–647. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Fettig, C.J.; Hansen, E.M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate Change and Bark Beetles of the Western United States and Canada: Direct and Indirect Effects. Bioscience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Logan, J.A.; Bentz, B.J. Model Analysis of Mountain Pine Beetle (Coleoptera: Scolytidae) Seasonality. Environ. Èntomol. 1999, 28, 924–934. [Google Scholar] [CrossRef]
- Jaworski, T.; Hilszczański, J. The effect of temperature and humidity changes on insects development their impact on forest ecosystems in the expected climate change. For. Res. Pap. 2013, 74, 345–355. [Google Scholar] [CrossRef]
- Ungerer, M.J.; Ayres, M.P.; Lombardero, M.J. Climate and the northern distribution limits of Dendroctonus frontalis Zimmer-mann (Coleoptera: Scolytidae). J. Biogeogr. 1999, 26, 1133–1145. [Google Scholar] [CrossRef]
- Carroll, A.L.; Taylor, S.W.; Régnière, J.; Safranyik, L. Effect of climate change on range expansion by the mountain pine beetle in British Columbia. In Mountain Pine Beetle Symposium: Challenges and Solutions, Information Report BC-X.-399; Natural Resources Canada: Victoria, BC, Canada, 2003; p. 298. [Google Scholar]
- Mendoza, M.G.; Salinas-Moreno, Y.; Olivo-Martínez, A.; Zúñiga, G. Factors influencing the geographical distribution of Den-droctonus rhizophagus (Coleoptera: Curculionidae: Scolytinae) in the Sierra Madre Occidental, Mexico. Environ. Entomol. 2011, 40, 549–559. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Forrest, J.R. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef]
- Burkett, V.R.; Wilcox, D.A.; Stottlemyer, R.; Barrow, W.; Fagre, D.; Baron, J.; Price, J.; Nielsen, J.L.; Allen, C.D.; Peterson, D.L.; et al. Nonlinear dynamics in ecosystem response to climatic change: Case studies and policy implications. Ecol. Complex. 2005, 2, 357–394. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Elith, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A. Novel methods improve prediction of species’ dis-tributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of consensus methods in predictive species distribution modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Bentz, B.J.; Jönsson, A.M. Modeling Bark Beetle Responses to Climate Change. In Bark Beetles; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 533–553. [Google Scholar]
- Sømme, L. Effects of glycerol on cold-hardiness in insects. Can. J. Zool. 1964, 42, 87–101. [Google Scholar] [CrossRef]
- Saunders, D.S. Insect photoperiodism: Effects of temperature on the induction of insect diapause and diverse roles for the circadian system in the photoperiodic response. Èntomol. Sci. 2013, 17, 25–40. [Google Scholar] [CrossRef]
- Davis, C.C.; Willis, C.G.; Primack, R.B.; Miller-Rushing, A.J. The importance of phylogeny to the study of phenological response to global climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 3201–3213. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climate Belt | Environmental Overview | Distribution Records |
|---|---|---|
| Northern warm temperate deciduous broad-leaved forest | The annual average temperature is 7–12 °C, the average temperature of the coldest month is −10∼−3 °C, the average temperature of the warmest month is 18∼27 °C, and the annual precipitation is 500–700 mm. | Shenyang, Huludao, Dalian, Dandong, Anshan, Liaoyang, Jinzhou, Yingkou, Panjin, Beijing, Tianjin, Taiyuan, Linfen, Changzhi, Shijiazhuang, Qinhuangdao, Baoding, Tangshan, Handan, Xingtai, Chengde, Jinan, Dezhou, Yan’an, Baoji, Tianshui |
| Temperate grassland | The annual average temperature is −3–−9 °C, the accumulated temperature of ≥ 10 °C is 1600–3200 °C, and the average temperature of coldest month is −7–−29 °C. The annual precipitation is 150–500 mm, mostly below 350 mm, mainly in summer. | Lanzhou, Pingliang, Altay, Hailar, Manzhouli, Qiqihar, Fuxin, Dandong, Daqing, Xining, Yinchuan, Tongliao, Yulin, Hohhot, Baotou, Zhangjiakou, Jining, Chifeng, Datong, Xilinhot |
| Temperate desert | This climate belt is distributed mainly in Xinjiang. The annual average temperature is 4–9 °C in northern Xinjiang and 7–14 °C in southern Xinjiang. The average temperature in January is −20–15 °C in northern Xinjiang and −10 °C–5 °C in southern Xinjiang. Most of the annual rainfall is below 50–100 mm, and the least is only 10–20 mm. | Urumqi, Shihezi, Karamay, Hami Kashgar, Wuwei, Jiuquan, Yumen, Jiayuguan, Golmud, Korla, Jinchang, Wuhai |
| Data Source | Category | Environmental Variables (unit) | Abbreviation |
|---|---|---|---|
| WorldClim | Bioclimatic | Annual mean temperature (°C) | Bio1 |
| Mean diurnal range (°C) | Bio2 | ||
| Isothermality (%) | Bio3 | ||
| Temperature seasonality (°C) | Bio4 | ||
| Maximum temperature of warmest month (°C) | Bio5 | ||
| Minimum temperature of coldest month (°C) | Bio6 | ||
| Temperature annual range (°C) | Bio7 | ||
| Mean temperature of wettest quarter (°C) | Bio8 | ||
| Mean temperature of driest quarter (°C) | Bio9 | ||
| Mean temperature of warmest quarter (°C) | Bio10 | ||
| Mean temperature of coldest quarter (°C) | Bio11 | ||
| Annual precipitation (mm) | Bio12 | ||
| Precipitation of wettest month (mm) | Bio13 | ||
| Precipitation of driest month (mm) | Bio14 | ||
| Precipitation seasonality | Bio15 | ||
| Precipitation of wettest quarter (mm) | Bio16 | ||
| Precipitation of driest quarter (mm) | Bio17 | ||
| Precipitation of warmest quarter (mm) | Bio18 | ||
| Precipitation of coldest quarter (mm) | Bio19 | ||
| USGS | Terrain | Altitude (m) | Alt. |
| Aspect (degree) | Asp. | ||
| Slope (degree) | Slop. | ||
| GBIF, CVH, Field investigations | Host | P. alba var. pyramidalis distribution | H |
| Rank | Environmental Variables | Regression Coefficients in LASSO | Contribution (%) | Probability of Selection |
|---|---|---|---|---|
| 1 | Mean temperature of coldest quarter | −0.8217 | 36.3 | 1.00 |
| 2 | Precipitation of wettest month | 0.5534 | 25.1 | 0.98 |
| 3 | Mean temperature of warmest quarter | −0.4035 | 17.7 | 0.96 |
| 4 | Mean temperature of driest quarter | −0.1864 | 9.6 | 0.93 |
| 5 | Mean diurnal range | −0.0569 | 7.4 | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ning, H.; Tang, M.; Chen, H. Mapping Invasion Potential of the Pest from Central Asia, Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae), in the Shelter Forests of Northwest China. Insects 2021, 12, 242. https://doi.org/10.3390/insects12030242
Ning H, Tang M, Chen H. Mapping Invasion Potential of the Pest from Central Asia, Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae), in the Shelter Forests of Northwest China. Insects. 2021; 12(3):242. https://doi.org/10.3390/insects12030242
Chicago/Turabian StyleNing, Hang, Ming Tang, and Hui Chen. 2021. "Mapping Invasion Potential of the Pest from Central Asia, Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae), in the Shelter Forests of Northwest China" Insects 12, no. 3: 242. https://doi.org/10.3390/insects12030242
APA StyleNing, H., Tang, M., & Chen, H. (2021). Mapping Invasion Potential of the Pest from Central Asia, Trypophloeus klimeschi (Coleoptera: Curculionidae: Scolytinae), in the Shelter Forests of Northwest China. Insects, 12(3), 242. https://doi.org/10.3390/insects12030242