
Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii

 ,
,  , ,
, ,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Assessment of the Attractiveness of Droskidrink® Food Baits Inoculated with Different Lactic Acid Bacteria
2.2. Assessment of The Performance of Oenococcus oeni Strains in Droskidrink® Food Bait
2.3. Electroantennography Responses of Drosophila suzukii Females to Volatile Collection of Oenococcus oeni Strains in Droskidrink® Food Bait
2.4. Data Analysis
3. Results
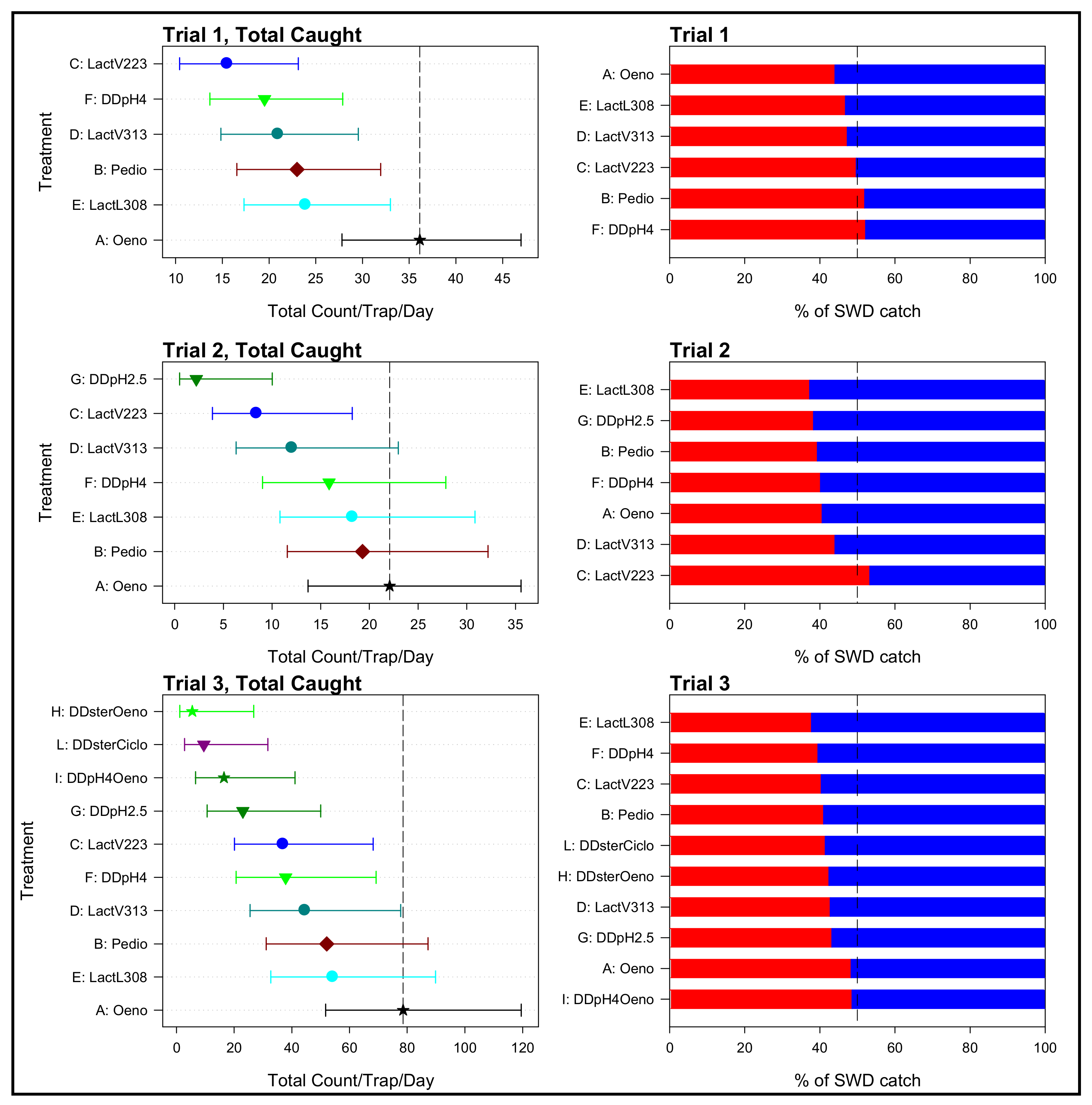
3.1. Field Assessment of the Attractiveness of Droskidrink® Food Baits Inoculated with Different Lactic Acid Bacteria
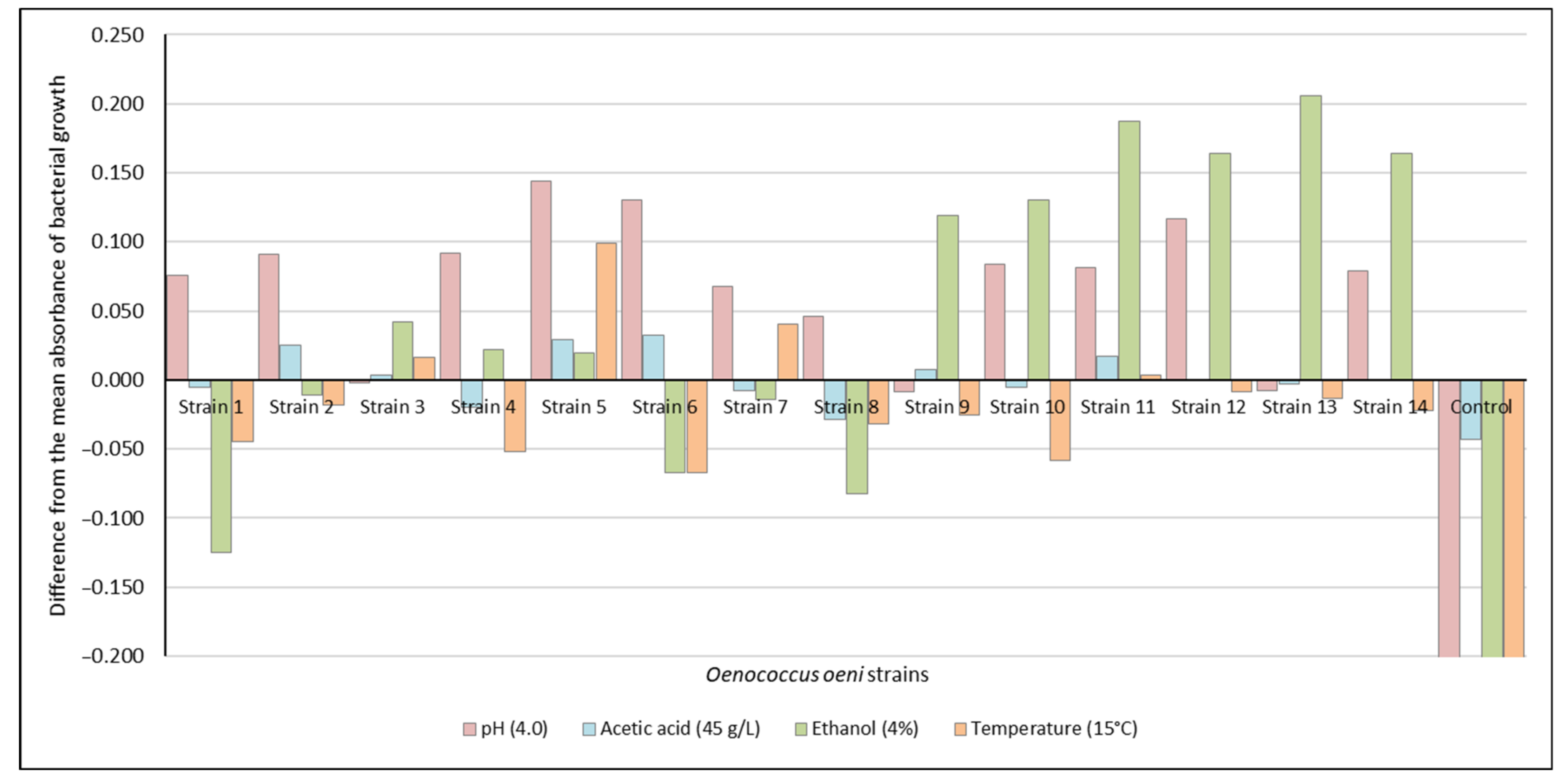
3.2. Assessment of the Performance of Oenococcus oeni Strains in Droskidrink® Food Bait
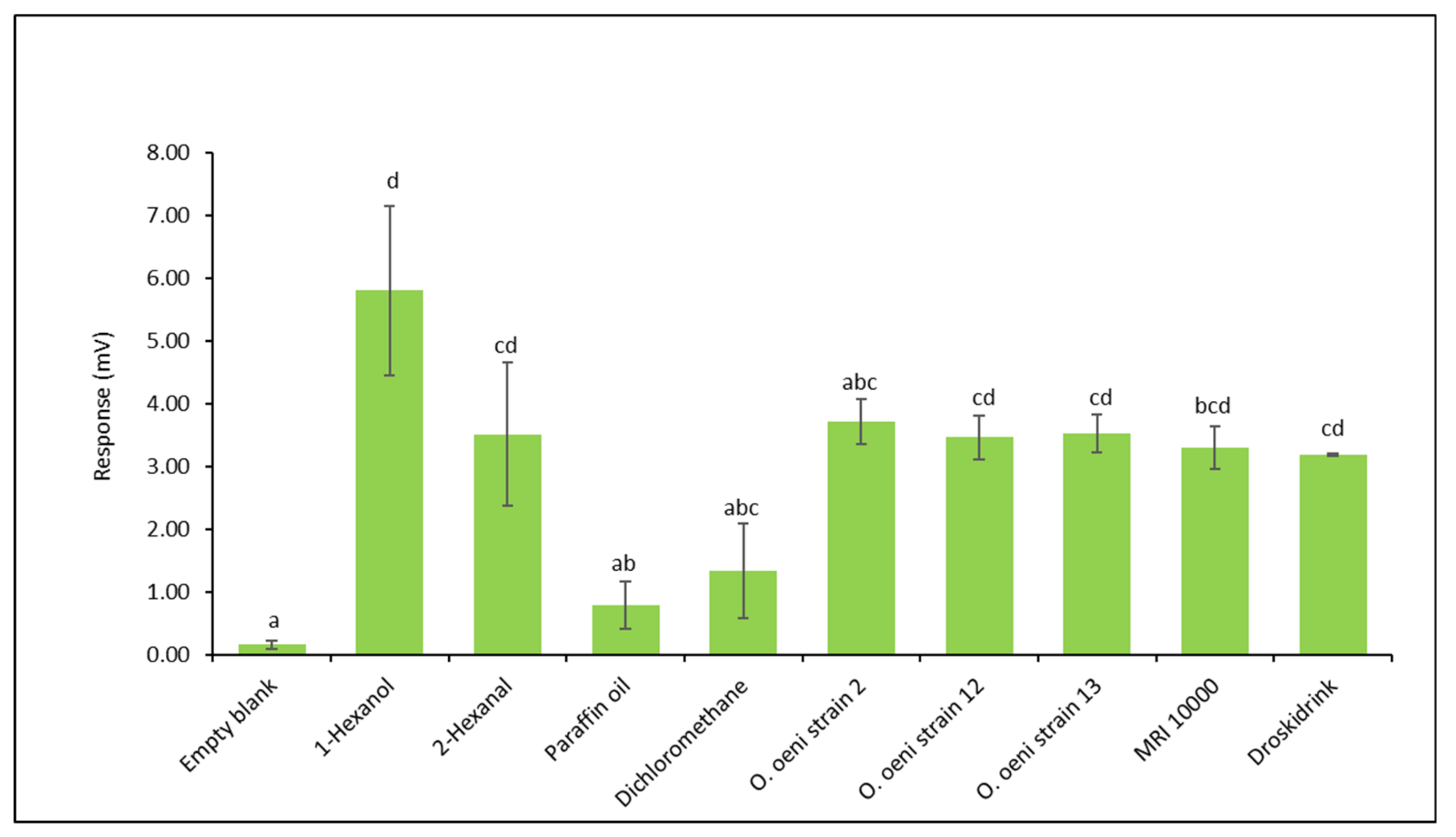
3.3. Electroantennography Responses of Drosophila suzukii Females to Volatile Collection of Oenococcus oeni Strains in Droskidrink® Food Bait
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In Focus: Spotted wing drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Rota-Stabelli, O.; Blaxter, M.; Anfora, G. Drosophila suzukii. Curr. Biol. 2013, 23, R8. [Google Scholar] [CrossRef]
- Pavlova, A.K.; Dahlmann, M.; Hauck, M.; Reineke, A. Laboratory bioassays with three different substrates to test the efficacy of insecticides against various stages of Drosophila suzukii (Diptera: Drosophilidae). J. Insect Sci. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Cahenzli, F.; Strack, T.; Daniel, C. Screening of 25 different natural crop protection products against Drosophila suzukii. J. Appl. Entomol. 2018, 142, 563–577. [Google Scholar] [CrossRef]
- Shawer, R.; Tonina, L.; Tirello, P.; Duso, C.; Mori, N. Laboratory and field trials to identify effective chemical control strategies for integrated management of Drosophila suzukii in European cherry orchards. Crop Prot. 2018, 103, 73–80. [Google Scholar] [CrossRef]
- Rice, K.B.; Short, B.D.; Leskey, T.C. Development of an attract-and-kill strategy for Drosophila suzukii (Diptera: Drosophilidae): Evaluation of attracticidal spheres under laboratory and field conditions. J. Econ. Entomol. 2017, 110, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Spies, J.M.; Liburd, O.E. Comparison of attractants, insecticides, and mass trapping for managing Drosophila suzukii (Diptera: Drosophilidae) in Blueberries. Fla. Entomol. 2019, 102, 315–321. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Grassi, A.; Dalton, D.T.; Miller, B.; Ouantar, M.; Loni, A.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. First field records of Pachycrepoideus vindemiae as a parasitoid of Drosophila suzukii in European and Oregon small fruit production areas. Entomologia 2013, 1, 11–16. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Audsley, N. Efficacy of commercially available invertebrate predators against Drosophila suzukii. Insects 2014, 5, 952–960. [Google Scholar] [CrossRef]
- Girod, P.; Rossignaud, L.; Haye, T.; Kenis, M. First results of the testing of Asian parasitoids as potential biological control agents of the Spotted Wing Drosophila, Drosophila suzukii. IOBC-WPRS Bull. 2017, 123, 186–187. [Google Scholar]
- Ibouh, K.; Oreste, M.; Bubici, G.; Tarasco, E.; Rossi-Stacconi, M.V.; Ioriatti, C.; Verrastro, V.; Anfora, G.; Baser, N. Biological control of Drosophila suzukii: Efficacy of parasitoids, entomopathogenic fungi, nematodes and deterrents of oviposition in laboratory assays. Crop Prot. 2019, 125. [Google Scholar] [CrossRef]
- Lasa, R.; Tadeo, E.; Toledo-Hérnandez, R.A.; Carmona, L.; Lima, I.; Williams, T. Improved capture of Drosophila suzukii by a trap baited with two attractants in the same device. PLoS ONE 2017, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Baroffio, C.A.; Richoz, P.; Fischer, S.; Kuske, S.; Linder, C.; Kehrli, P. Monitoring Drosophila suzukii in Switzerland in 2012. J. Berry Res. 2014, 4, 47–52. [Google Scholar] [CrossRef]
- Iglesias, L.E.; Nyoike, T.W.; Liburd, O.E. Effect of trap design, bait type, and age on captures of Drosophila suzukii (Diptera: Drosophilidae) in berry crops. J. Econ. Entomol. 2014, 107, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Vaccari, G.; Caruso, S.; Nouhaud, C.; Maistrello, L. Comparison of different trap types for Drosophila suzukii in cherry fields. IOBC-WPRS Bull. 2015, 109, 211–214. [Google Scholar]
- Mazzetto, F.; Pansa, M.G.; Ingegno, B.L.; Tavella, L.; Alma, A. Monitoring of the exotic fly Drosophila suzukii in stone, pome and soft fruit orchards in NW Italy. J. Asia Pac. Entomol. 2015, 18, 321–329. [Google Scholar] [CrossRef]
- Cha, D.H.; Loeb, G.M.; Linn, C.E.; Hesler, S.P.; Landolt, P.J. A Multiple-choice bioassay approach for rapid screening of key attractant volatiles. Environ. Entomol. 2018, 47, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Tonina, L.; Grassi, A.; Caruso, S.; Mori, N.; Gottardello, A.; Anfora, G.; Giomi, F.; Vaccari, G.; Ioriatti, C. Comparison of attractants for monitoring Drosophila suzukii in sweet cherry orchards in Italy. J. Appl. Entomol. 2018, 142, 18–25. [Google Scholar] [CrossRef]
- Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Beliën, T.; Bylemans, D. Olfactory preference of Drosophila suzukii shifts between fruit and fermentation cues over the season: Effects of physiological status. Insects 2019, 10, 200. [Google Scholar] [CrossRef]
- Mazzetto, F.; Gonella, E.; Crotti, E.; Vacchini, V.; Syrpas, M.; Pontini, M.; Mangelinckx, S.; Daffonchio, D.; Alma, A. Olfactory attraction of Drosophila suzukii by symbiotic acetic acid bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Hayek, A.S.; Ibrahim, A.S. Current limitations and challenges with lactic acid bacteria: A review. Food Nutr. Sci. 2013, 73–87. [Google Scholar] [CrossRef]
- Hoefnagel, M.H.N.; Starrenburg, M.J.C.; Martens, D.E.; Hugenholtz, J.; Kleerebezem, M.; Van Swam, I.I.; Bongers, R.; Westerhoff, H.V.; Snoep, J.L. Metabolic engineering of lactic acid bacteria, the combined approach: Kinetic modelling, metabolic control and experimental analysis. Microbiology 2002, 148, 1003–1013. [Google Scholar] [CrossRef]
- Endo, A.; Shintaro, M.; Yasuhiro, T.; Wolfgang, K.; Masanori, A.; Leon, D.; Salminenf, S. Fructophilic lactic acid bacteria, a unique group of fructose-fermenting microbes. Appl. Environ. Microbiol. 2018, 84, 1–14. [Google Scholar] [CrossRef]
- Capozzi, V.; Russo, P.; Beneduce, L.; Weidmann, S.; Grieco, F.; Guzzo, J.; Spano, G. Technological properties of Oenococcus oeni strains isolated from typical southern Italian wines. Lett. Appl. Microbiol. 2010, 50, 327–334. [Google Scholar] [CrossRef]
- Venu, I.; Durisko, Z.; Xu, J.; Dukas, R. Social attraction mediated by fruit flies’ microbiome. J. Exp. Biol. 2014, 217, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut microbiota affects development and olfactory behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef]
- Vacchini, V.F. Microbial Ecology of the Spotted Wing Fly Drosophila suzukii. Ph.D. Thesis, Università Degli Studi di Milano, Milano, Italy, 2014. [CrossRef]
- Grassi, A.; Anfora, G.; Maistri, S.; Maddalena, G.; De Gristofaro, A.; Savini, G.; Savini, G.; Ioriatti, C. Development and efficacy of Droskidrink, a food bait for trapping Drosophila suzukii. IOBC-WPRS Bull. 2015, 109, 26–28. [Google Scholar]
- Solieri, L.; Genova, F.; De Paola, M.; Giudici, P. Characterization and technological properties of Oenococcus oeni strains from wine spontaneous malolactic fermentations: A framework for selection of new starter cultures. J. Appl. Microbiol. 2010, 108, 285–298. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman & Hall: London, UK, 1989; p. 511. [Google Scholar]
- Guzzon, R.; Poznanski, E.; Conterno, L.; Vagnoli, P.; Krieger-Weber, S.; Cavazza, A. Selection of a new highly resistant strain for malolactic fermentation under difficult conditions. S. Afr. J. Enol. Vitic. 2009, 30, 133–141. [Google Scholar] [CrossRef][Green Version]
- Salvagnin, U.; Malnoy, M.; Thöming, G.; Tasin, M.; Carlin, S.; Martens, S.; Vrhovsek, U.; Angeli, S.; Anfora, G. Adjusting the scent ratio: Using genetically modified Vitis vinifera plants to manipulate European grapevine moth behaviour. Plant Biotechnol. J. 2018, 16, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Revadi, S.; Vitagliano, S.; Rossi-Stacconi, M.V.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Payne, R.; Murray, D.; Baird, D. The Guide to the Genstat Command Language (Release 19); VSN International, Hemel Hempsted: Hertfordshire, UK, 2017. [Google Scholar]
- Lonvaud-Funel, A. Biogenic amines in wines: Role of lactic acid bacteria. FEMS Microbiol. Lett. 2001, 199, 9–13. [Google Scholar] [CrossRef]
- Liu, S.Q. Malolactic fermentation in wine—beyond deacidification. J. Appl. Microbiol. 2002, 92, 589–601. [Google Scholar] [CrossRef]
- Spano, G.; Massa, S. Environmental stress response in wine lactic acid bacteria: Beyond Bacillus subtilis. Crit. Rev. Microbiol. 2006, 32, 77–86. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef]
- Succi, M.; Pannella, G.; Tremonte, P.; Tipaldi, L.; Coppola, R.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E. Sub-optimal pH preadaptation improves the survival of Lactobacillus plantarum strains and the malic acid consumption in wine-like medium. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Nisiotou, A.A.; Dourou, D.; Filippousi, M.E.; Diamantea, E.; Fragkoulis, P.; Tassou, C.; Banilas, G. Genetic and technological characterisation of vineyard- and winery-associated lactic acid bacteria. BioMed Res. Int. 2015, 1–8. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; du Toit, M. Malolactic fermentation: The ABC’s of MLF. S. Afr. J. Enol. Vitic. 2010, 31, 186–212. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguez-Saona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Ðurović, D.; Alawamleh, A.; Carlin, S.; Maddalena, G.; Guzzon, R.; Mazzoni, V.; Dalton, D.T.; Walton, V.M.; Suckling, D.M.; Butler, R.C.; et al. Liquid Baits with Oenococcus oeni increase captures of Drosophila suzukii. Insects 2021, 12, 66. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Composition |
|---|---|
| Bait A | DD (pH 4.0) inoculated with Oenococcus oeni ATCCBAA-331 |
| Bait B | DD (pH 4.0) inoculated with Pediococcus spp. |
| Bait C | DD (pH 4.0) inoculated with Lactobacillus spp. V223 |
| Bait D | DD (pH 4.0) inoculated with Lactobacillus spp. V313 |
| Bait E | DD (pH 4.0) inoculated with Lactobacillus spp. L308 |
| Bait F | DD at pH 4.0 |
| Bait G | DD at pH 2.5 (commercial version) |
| Bait H | DD at pH 2.5 pasteurized at 70 °C for 30 min and inoculated with Oenococcus oeni |
| Bait I | DD at pH 4.0 pasteurized at 70 °C for 30 min, mixed with 10 mL/L of cycloheximide aqueous solution (0.01%), and inoculated with Oenococcus oeni |
| Bait L | DD at pH 4.0 mixed with 1 g of tetracycline and inoculated with Oenococcus oeni |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alawamleh, A.; Ðurović, G.; Maddalena, G.; Guzzon, R.; Ganassi, S.; Hashmi, M.M.; Wäckers, F.; Anfora, G.; Cristofaro, A.D. Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii. Insects 2021, 12, 153. https://doi.org/10.3390/insects12020153
Alawamleh A, Ðurović G, Maddalena G, Guzzon R, Ganassi S, Hashmi MM, Wäckers F, Anfora G, Cristofaro AD. Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii. Insects. 2021; 12(2):153. https://doi.org/10.3390/insects12020153
Chicago/Turabian StyleAlawamleh, Amani, Gordana Ðurović, Giuseppe Maddalena, Raffaele Guzzon, Sonia Ganassi, Maaz Maqsood Hashmi, Felix Wäckers, Gianfranco Anfora, and Antonio De Cristofaro. 2021. "Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii" Insects 12, no. 2: 153. https://doi.org/10.3390/insects12020153
APA StyleAlawamleh, A., Ðurović, G., Maddalena, G., Guzzon, R., Ganassi, S., Hashmi, M. M., Wäckers, F., Anfora, G., & Cristofaro, A. D. (2021). Selection of Lactic Acid Bacteria Species and Strains for Efficient Trapping of Drosophila suzukii. Insects, 12(2), 153. https://doi.org/10.3390/insects12020153