
The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms



{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
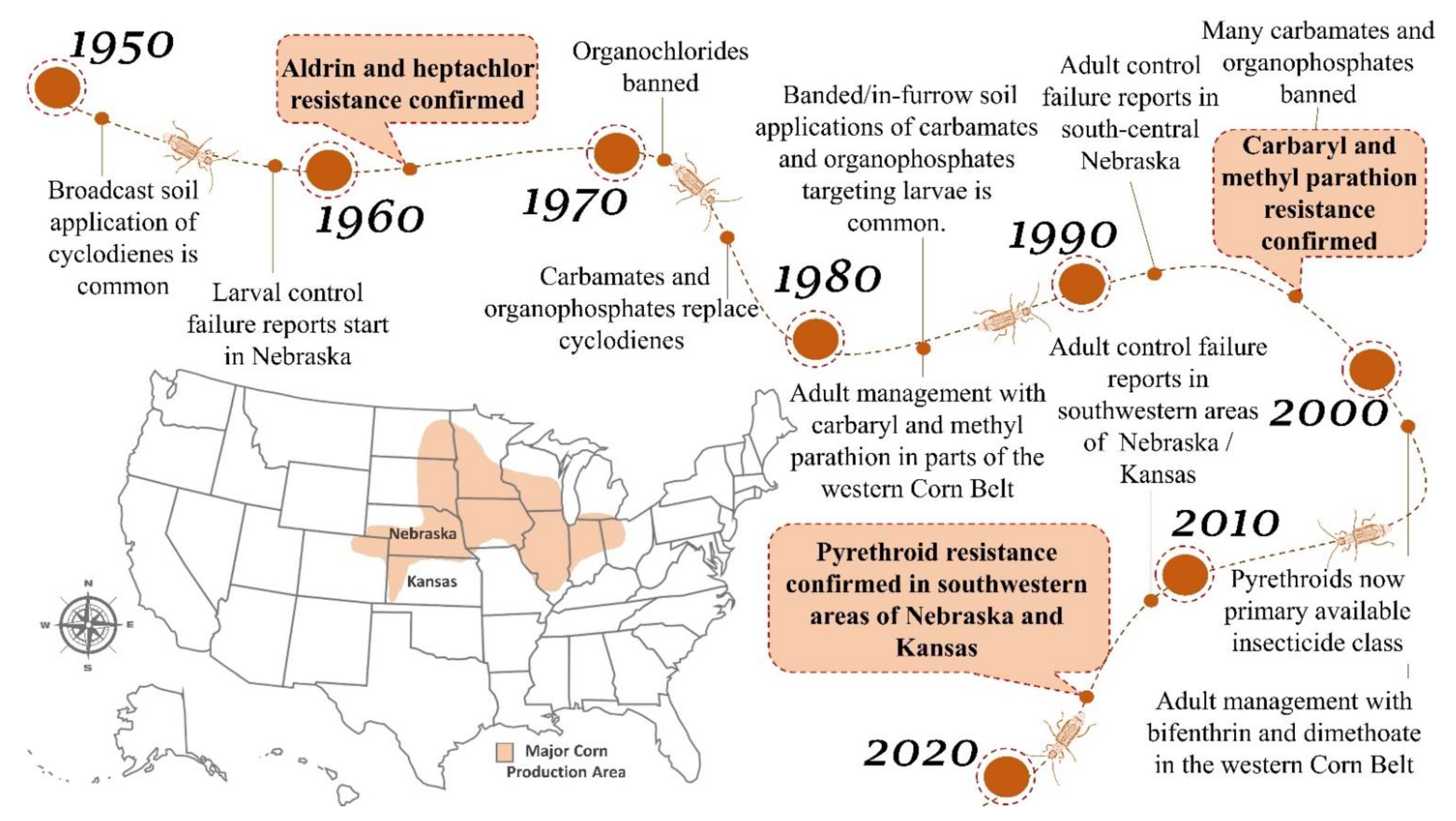
2. Western Corn Rootworm—Insecticide History
2.1. Soil Insecticides
2.2. Seed Treatments
2.3. Foliar Insecticides
3. Mechanisms of Dvv Resistance to Insecticides
3.1. Organochlorides
3.2. Carbamates and Organophosphates
3.3. Pyrethroids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sappington, T.W.; Siegfried, B.D.; Guillemaud, T. Coordinated Diabrotica genetics research: Accelerating progress on an urgent insect pest problem. Am. Entomol. 2006, 52, 90–97. [Google Scholar] [CrossRef]
- Wechsler, S.; Smith, D. Has resistance taken root in U.S. corn fields? Demand for insect control. Am. J. Agric. Econ. 2018, 100, 1136–1150. [Google Scholar] [CrossRef]
- Branson, T.F.; Ortman, E.E. Host range of larvae of the western corn rootworm. J. Econ. Entomol. 1967, 60, 201–203. [Google Scholar] [CrossRef]
- Branson, T.F.; Ortman, E.E. The host range of larvae of the western corn rootworm: Further studies. J. Econ. Entomol. 1970, 63, 800–803. [Google Scholar] [CrossRef]
- Branson, T.F. Resistance in the grass tribe Maydeae to larvae of the western corn rootworm. Ann. Entomol. Soc. Am. 1971, 64, 861–863. [Google Scholar] [CrossRef]
- Clark, T.L.; Hibbard, B.E. Comparison of nonmaize hosts to support western corn rootworm (Coleoptera: Chrysomelidae) larval biology. Environ. Entomol. 2004, 33, 681–689. [Google Scholar] [CrossRef]
- Oyediran, I.O.; Hibbard, B.E.; Clark, T.L. Prairie grasses as hosts of the western corn rootworm (Coleoptera: Chrysomelidae). Environ. Entomol. 2004, 33, 740–747. [Google Scholar] [CrossRef]
- Moeser, J.; Vidal, S. Do Alternative host plants enhance the invasion of the maize pest Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae, Galerucinae) in Europe? Environ. Entomol. 2004, 33, 1169–1177. [Google Scholar] [CrossRef]
- Campbell, L.A.; Meinke, L.J. Seasonality and adult habitat use by four Diabrotica species at prairie-corn interfaces. Environ. Entomol. 2006, 35, 922–936. [Google Scholar] [CrossRef]
- Moeser, J.; Hibbard, B.E. A synopsis of the nutritional ecology of larvae and adults of Diabrotica virgifera virgifera (LeConte) in the new and old world—Nouvelle cuisine for the invasive maize pest Diabrotica virgifera virgifera in Europe? In Western Corn Rootworm: Ecology and Management; Vidal, S., Kuhlmann, U., Edwards, C.R., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 41–65. [Google Scholar]
- Branson, T.F.; Krysan, J.L. Feeding and oviposition behavior and life cycle strategies of Diabrotica: An evolutionary view with implications for pest management. Environ. Entomol. 1981, 10, 826–831. [Google Scholar] [CrossRef]
- Prystupa, B.; Ellis, C.R.; Teal, P.E.A. Attraction of adult Diabrotica (Coleoptera: Chrysomelidae) to corn silks and analysis of the host-finding response. J. Chem. Ecol. 1988, 14, 635–651. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, S.E. Flight orientation of Diabrotica virgifera virgifera and D. barberi (Coleoptera: Chrysomelidae) at habitat interfaces. Ann. Entomol. Soc. Am. 1994, 87, 383–394. [Google Scholar] [CrossRef]
- Darnell, S.J.; Meinke, L.J.; Young, L.J. Influence of corn phenology on adult western corn rootworm (Coleoptera: Chrysomelidae) distribution. Environ. Entomol. 2000, 29, 587–595. [Google Scholar] [CrossRef]
- LeConte, J.L. New Coleoptera collected on the survey for the extension of the Union Pacific Railway, E. D. from Kansas to Fort Craig, New Mexico. In Transactions of the American Entomological Society; The Society: Philadelphia, PA, USA, 1868; Volume 2, pp. 58–59. [Google Scholar]
- Segura-Leon, O.L. Phylogeography of Diabrotica virgifera virgifera LeConte and Diabrotica virgifera zeae Krysan and Smith (Coleoptera: Chrysomelidae). Ph.D Thesis, The University of Nebraska-Lincoln, Lincoln, NE, USA, 2004; pp. 1–143. [Google Scholar]
- Lombaert, E.; Ciosi, M.; Miller, N.J.; Sappington, T.W.; Blin, A.; Guillemaud, T. Colonization history of the western corn rootworm (Diabrotica virgifera virgifera) in North America: Insights from random forest ABC using microsatellite data. Biol. Invasions 2018, 20, 665–677. [Google Scholar] [CrossRef]
- Meinke, L.J.; Sappington, T.W.; Onstad, D.W.; Guillemaud, T.; Miller, N.J.; Komáromi, J.; Levay, N.; Furlan, L.; Kiss, J.; Toth, F. Western corn rootworm (Diabrotica virgifera virgifera LeConte) population dynamics. Agric. For. Entomol. 2009, 11, 29–46. [Google Scholar] [CrossRef]
- Gillette, C.P. Diabrotica virgifera Lec. as a corn root-worm. J. Econ. Entomol. 1912, 5, 364–366. [Google Scholar] [CrossRef]
- Bare, O.S. Corn Rootworm Does Damage in Southwestern Nebraska; Annual Report of Cooperative Extension Work in Agriculture and Home Economics, Entomology, State of Nebraska; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1930; p. 21. [Google Scholar]
- Bare, O.S. Corn Rootworms; University Nebraska Cooperative Extension Circular 1506; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1931. [Google Scholar]
- Tate, H.D.; Bare, O.S. Corn Rootworms; Bulletin of the Agricultural Experiment Station of Nebraska; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1946; pp. 1–12. [Google Scholar]
- Hill, R.E.; Hixson, E.; Muma, M.H. Corn rootworm control tests with benzene hexachloride DDT, nitrogen fertilizers and crop rotations. J. Econ. Entomol. 1948, 41, 392–401. [Google Scholar] [CrossRef]
- Lilly, J.H. Soil insects and their control. Annu. Rev. Entomol. 1956, 1, 203–222. [Google Scholar] [CrossRef]
- Ball, H.J. Larval and Adult Control Recommendations and Insecticide Resistance Data for Corn Rootworms in Nebraska (1948–1981); Department of Entomology Report; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1981; pp. 1–16. [Google Scholar]
- Wangila, D.S.; Gassmann, A.J.; Petzold-Maxwell, J.L.; French, B.W.; Meinke, L.J. Susceptibility of Nebraska western corn rootworm (Coleoptera: Chrysomelidae) populations to Bt corn events. J. Econ. Entomol. 2015, 108, 742–751. [Google Scholar] [CrossRef]
- Gray, M.E.; Sappington, T.W.; Miller, N.J.; Moeser, J.; Bohn, M.O. Adaptation and invasiveness of western corn rootworm: Intensifying research on a worsening pest. Annu. Rev. Entomol. 2009, 54, 303–321. [Google Scholar] [CrossRef]
- Levine, E.; Oloumi-Sadeghi, H. Management of diabroticite rootworms in corn. Annu. Rev. Entomol. 1991, 36, 229–255. [Google Scholar] [CrossRef]
- Muma, M.H.; Hill, R.E.; Hixson, E. Soil treatments for corn rootworm control. J. Econ. Entomol. 1949, 42, 822–824. [Google Scholar] [CrossRef]
- Munson, J.D.; Hill, R.E. Insecticides applied at planting time for control of corn rootworm larvae in Nebraska. J. Econ. Entomol. 1970, 63, 1614–1617. [Google Scholar] [CrossRef]
- Mayo, Z.B. A Five-Year Comparison of Insecticides Applied to Control Larvae of Western and Northern Corn Rootworms; Agricultural Experiment Station, University of Nebraska-Lincoln: Lincoln, NE, USA, 1975; pp. 1–15. [Google Scholar]
- Mayo, Z.B. Aerial Suppression of Rootworm Adults for Larval Control; Agricultural Experiment Station, University of Nebraska-Lincoln: Lincoln, NE, USA, 1976; pp. 1–12. [Google Scholar]
- Mayo, Z.B. Field evaluation of insecticides for control of larvae of corn rootworms. In Methods for the Study of Pest Diabrotica; Springer Series in Experimental Entomology; Springer: New York, NY, USA, 1986; pp. 183–203. ISBN 978-1-4612-9338-5. [Google Scholar]
- Pruess, K.P.; Witkowski, J.F.; Raun, E.S. Population suppression of western corn rootworm by adult control with ULV malathion. J. Econ. Entomol. 1974, 67, 651–655. [Google Scholar] [CrossRef]
- Chio, H.; Chang, C.-S.; Metcalf, R.L.; Shaw, J. Susceptibility of four species of Diabrotica to insecticides. J. Econ. Entomol. 1978, 71, 389–393. [Google Scholar] [CrossRef]
- Sutter, G.R. Comparative toxicity of insecticides for corn rootworm (Coleoptera: Chrysomelidae) larvae in a soil bioassay. J. Econ. Entomol. 1982, 75, 489–491. [Google Scholar] [CrossRef]
- Fuller, B.W.; Boetel, M.A.; Walgenbach, D.D.; Grundler, J.A.; Hein, G.L.; Jarvi, K.J.; Keaster, A.J.; Landis, D.A.; Meinke, L.J.; Oleson, J.D.; et al. Optimization of soil insecticide rates for managing corn rootworm (Coleoptera: Chrysomelidae) larvae in the North Central United States. J. Econ. Entomol. 1997, 90, 1332–1340. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Wilde, G.E.; Sloderbeck, P.E.; Buschman, L.L.; Higgins, R.A.; Whitworth, R.J.; Bowling, R.A.; Starkey, S.R.; He, F. Comparative susceptibility of western corn rootworm (Coleoptera: Chrysomelidae) adults to selected insecticides in Kansas. J. Econ. Entomol. 2005, 98, 2181–2187. [Google Scholar] [CrossRef]
- Douglas, M.R.; Tooker, J.F. Large-scale deployment of seed treatments has driven rapid increase in use of neonicotinoid insecticides and preemptive pest management in U.S. field crops. Environ. Sci. Technol. 2015, 49, 5088–5097. [Google Scholar] [CrossRef]
- USEPA. Current & Previously Registered Section 3 PIP Registrations. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/pip/pip_list.htm (accessed on 16 December 2020).
- Andow, D.A.; Pueppke, S.G.; Schaafsma, A.W.; Gassmann, A.J.; Sappington, T.W.; Meinke, L.J.; Mitchell, P.D.; Hurley, T.M.; Hellmich, R.L.; Porter, R.P. Early detection and mitigation of resistance to Bt maize by western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2016, 109, 1–12. [Google Scholar] [CrossRef]
- USDA-NASS. United States: Corn Production. Available online: https://ipad.fas.usda.gov/rssiws/al/crop_production_maps/us/USA_Corn_Total_Lev2_Prod.png (accessed on 30 December 2020).
- USDA-NASS. Corn for Grain 2019 Production by County for Selected States. Available online: https://www.nass.usda.gov/Charts_and_Maps/Crops_County/cr-pr.php (accessed on 30 December 2020).
- Cox, H.C.; Lilly, J.H. Chemical control of the corn rootworm. J. Econ. Entomol. 1953, 46, 217–224. [Google Scholar] [CrossRef]
- Ball, H.J.; Hill, R.E. You can control corn rootworm. Neb. Exp. Stn. Quart. 1953, 1, 3–4. [Google Scholar]
- Ball, H.J.; Roselle, R.E. You Can Control Corn Rootworms; University Nebraska College of Agricultural Extension Circular; University of Nebraska-Lincoln: Lincoln, NE, USA, 1954. [Google Scholar]
- Ball, H.J. Aldrin and dieldrin residues in Nebraska soils as related to decreasing LD50 values for the adult western corn rootworm—1962–1981. J. Environ. Sci. Health B 1983, 18, 735–744. [Google Scholar] [CrossRef]
- Roselle, R.E.; Anderson, L.W.; Simpson, R.C.; Webb, M.C. Annual Report for 1959; Cooperative Extension Work in Entomology; University of Nebraska-Lincoln: Lincoln, NE, USA, 1959. [Google Scholar]
- Roselle, R.E.; Anderson, L.W.; Simpson, R.C.; Bergman, P.W. Annual Report for 1960; Cooperative Extension Work in Entomology; University of Nebraska-Lincoln: Lincoln, NE, USA, 1960. [Google Scholar]
- Roselle, R.E.; Anderson, L.W.; Bergman, P.W. Annual Report for 1961; Cooperative Extension Work in Entomology; University of Nebraska-Lincoln: Lincoln, NE, USA, 1961. [Google Scholar]
- Weekman, G.T. Problem in rootworm control. Proc. North Cent. Branch Entomol. Soc. Am. 1961, 16, 32–34. [Google Scholar]
- Ball, H.J.; Weekman, G.T. Insecticide resistance in the adult western corn rootworm in Nebraska. J. Econ. Entomol. 1962, 55, 439–441. [Google Scholar] [CrossRef]
- Ball, H.J.; Weekman, G.T. Differential resistance of corn rootworms to insecticides in Nebraska and adjoining states. J. Econ. Entomol. 1963, 56, 553–555. [Google Scholar] [CrossRef]
- Ball, H.J. On the biology and egg-laying habits of the western corn rootworm. J. Econ. Entomol. 1957, 50, 126–128. [Google Scholar] [CrossRef]
- Hamilton, E.W. Aldrin resistance in corn rootworm beetles. J. Econ. Entomol. 1965, 58, 296–300. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Mullin, C.A. Influence of alternative host plant feeding on aldrin susceptibility and detoxification enzymes in western and northern corn rootworms. Pestic. Biochem. Physiol. 1989, 35, 155–164. [Google Scholar] [CrossRef]
- Metcalf, R.L. Foreword. In Methods for the Study of Pest Diabrotica; Krysan, J.L., Miller, T.A., Eds.; Springer: New York, NY, USA, 1986; pp. vii–xv. [Google Scholar]
- Metcalf, R.L. The ecology of insecticides and the chemical control of insects. In Ecological Theory and Integrated Pest Management Practice; Kogan, M., Ed.; Wiley: New York, NY, USA, 1986; pp. 251–297. [Google Scholar]
- Felsot, A.S. Enhanced biodegradation of insecticides in soil: Implications for agroecosystems. Ann. Rev. Entomol. 1989, 34, 453–476. [Google Scholar] [CrossRef]
- Turpin, F.T.; Thieme, J.M. Impact of soil insecticide usage on corn production in Indiana: 1972–19741. J. Econ. Entomol. 1978, 71, 83–86. [Google Scholar] [CrossRef]
- Stamm, D.E.; Mayo, Z.B.; Campbell, J.B.; Witkowski, J.F.; Andersen, L.W.; Kozub, R. Western corn rootworm (Coleoptera: Chrysomelidae) beetle counts as a means of making larval control recommendations in Nebraska. J. Econ. Entomol. 1985, 78, 794–798. [Google Scholar] [CrossRef]
- Gray, M.E.; Steffey, K.L.; Oloumi-Sadeghi, H. Participatory on-farm research in Illinois cornfields: An evaluation of established soil insecticide rates and prevalence of com rootworm (Coleoptera: Chrysomelidae) injury. J. Econ. Entomol. 1993, 86, 1473–1482. [Google Scholar] [CrossRef]
- USEPA. EPA Pesticide Fact Sheet: Tefluthrin. Available online: https://nepis.epa.gov/Exe/ZyPDF.cgi/P100XZAX.PDF?Dockey=P100XZAX.PDF (accessed on 16 December 2020).
- Johnson, M.; Luukinen, B.; Gervais, J.; Buhl, K.; Stone, D. Bifenthrin General Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services: Corvallis, OR, USA, 2010. [Google Scholar]
- Harris, C.R. Factors influencing the effectiveness of soil insecticides. Annu. Rev. Entomol. 1972, 17, 177–198. [Google Scholar] [CrossRef]
- Mayo, Z.B. Influences of rainfall and sprinkler irrigation on the residual activity of insecticides applied to corn for control of adult western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1984, 77, 190–193. [Google Scholar] [CrossRef]
- Sutter, G.R.; Gustin, R.D. Environmental factors influencing corn rootworm biology and control. In Proceedings of the Illinois Agricultural Pesticides Conference; University of Illinois Cooperative Extension Service: Urbana-Champaign, IL, USA, 1989; pp. 43–48. [Google Scholar]
- Sutter, G.R.; Branson, T.F.; Fisher, J.R.; Elliott, N.C.; Jackson, J.J. Effect of insecticide treatments on root damage ratings of maize in controlled infestations of western corn rootworms (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1989, 82, 1792–1798. [Google Scholar] [CrossRef]
- Riedell, W.E.; Sutter, G.R. Soil moisture and survival of western corn rootworm larvae in field plots. J. Kans. Entomol. Soc. 1995, 68, 80–84. [Google Scholar]
- Gorder, G.W.; Dahm, P.A.; Tollefson, J.J. Carbofuran persistence in cornfield soils. J. Econ. Entomol. 1982, 75, 637–642. [Google Scholar] [CrossRef]
- Harris, C.R.; Chapman, R.A.; Harris, C.; Tu, C.M. Biodegradation of pesticides in soil: Rapid induction of carbamate degrading factors after carbofuran treatment. J. Environ. Sci. Health B 1984, 19, 1–11. [Google Scholar] [CrossRef]
- Walker, A.; Suett, D.L. Enhanced degradation of pesticides in soil: A potential problem for continued pest, disease and weed control. Asp. Appl. Biol. 1986, 12, 95–103. [Google Scholar]
- Tollefson, J.J. Accelerated degradation of soil insecticides used for corn rootworm control. In Proceedings of the British Crop Protection Conference Pests and Diseases, Brighton Metropole, UK, 17–20 November 1986; pp. 1131–1136. [Google Scholar]
- Williams, I.H.; Pepin, H.S.; Brown, M.J. Degradation of carbofuran by soil microorganisms. Bull. Environ. Contam. Toxicol. 1976, 15, 244–249. [Google Scholar] [CrossRef]
- Felsot, A.S.; Wilson, J.G.; Kuhlman, D.E.; Steffey, K.L. Rapid dissipation of carbofuran as a limiting factor in corn rootworm (Coleoptera: Chrysomelidae) control in fields with histories of continous carbofuran use. J. Econ. Entomol. 1982, 75, 1098–1103. [Google Scholar] [CrossRef]
- Abou-Assaf, N.; Coats, J.R.; Gray, M.E.; Tollefson, J.J. Degradation of isofenphos in cornfields with conservation tillage practices. J. Environ. Sci. Health B 1986, 21, 425–446. [Google Scholar] [CrossRef]
- Racke, K.D.; Coats, J.R. Enhanced degradation of isofenphos by soil microorganisms. J. Agric. Food Chem. 1987, 35, 94–99. [Google Scholar] [CrossRef]
- Hills, T.M.; Peters, D.C. Methods of applying insecticides for controlling western corn rootworm larvae. J. Econ. Entomol. 1972, 65, 1714–1718. [Google Scholar] [CrossRef]
- Mayo, Z.B. Influence of planting dates on the efficacy of soil insecticides applied to control larvae of the western and northern corn rootworm. J. Econ. Entomol. 1980, 73, 211–212. [Google Scholar] [CrossRef]
- Hoffmann, M.P.; Kirkwyland, J.J.; Gardner, J. Impact of western corn rootworm (Coleoptera: Chrysomelidae) on sweet corn and evaluation of insecticidal and cultural control options. J. Econ. Entomol. 2000, 93, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Mayo, Z.B.; Peters, L.L. Planting vs. cultivation time applications of granular soil insecticides to control larvae of corn rootworms in Nebraska. J. Econ. Entomol. 1978, 71, 801–803. [Google Scholar] [CrossRef]
- Felsot, A.S.; Bruce, W.N.; Steffey, K.L. Degradation of terbufos (Counter) soil insecticide in cornfields under conservation tillage practices. Bull. Environ. Contam. Toxicol. 1987, 38, 369–376. [Google Scholar] [CrossRef]
- Gray, M.E.; Felsot, A.S.; Steffey, K.L.; Levine, E. Planting time application of soil insecticides and western corn rootworm (Coleoptera: Chrysomelidae) emergence: Implications for long-term management programs. J. Econ. Entomol. 1992, 85, 544–553. [Google Scholar] [CrossRef]
- Furlan, L.; Canzi, S.; Di Bernardo, A.; Edwards, C.R. The ineffectiveness of insecticide seed coatings and planting-time soil insecticides as Diabrotica virgifera virgifera LeConte population suppressors. J. Appl. Entomol. 2006, 130, 485–490. [Google Scholar] [CrossRef]
- Boetel, M.A.; Fuller, B.W.; Evenson, P.D. Emergence of adult northern and western corn rootworms (Coleoptera: Chrysomelidae) following reduced soil insecticide applications. J. Econ. Entomol. 2003, 96, 714–729. [Google Scholar] [CrossRef] [PubMed]
- Souza, D.; Peterson, J.A.; Wright, R.J.; Meinke, L.J. Field efficacy of soil insecticides on pyrethroid-resistant western corn rootworms (Diabrotica virgifera virgifera LeConte). Pest Manag. Sci. 2020, 76, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Parimi, S.; Meinke, L.J.; Wade French, B.; Chandler, L.D.; Siegfried, B.D. Stability and persistence of aldrin and methyl-parathion resistance in western corn rootworm populations (Coleoptera: Chrysomelidae). Crop Prot. 2006, 25, 269–274. [Google Scholar] [CrossRef]
- Ball, H.J.; Su, P.P. Effect of sublethal dosages of carbofuran and carbaryl on fecundity and longevity of the female western corn rootworm. J. Econ. Entomol. 1979, 72, 873–876. [Google Scholar] [CrossRef]
- Sutter, G.R.; Branson, T.F.; Fisher, J.R.; Elliott, N.C. Effect of insecticides on survival, development, fecundity, and sex ratio in controlled infestations of western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1991, 84, 1905–1912. [Google Scholar] [CrossRef]
- Boetel, M.A.; Fuller, B.W.; Chandler, L.D.; Hovland, D.G.; Evenson, P.D. Fecundity and egg viability of northern and western corn rootworm (Coleoptera: Chrysomelidae) beetles surviving labeled and reduced soil insecticide applications. J. Econ. Entomol. 1998, 91, 274–279. [Google Scholar] [CrossRef]
- Petzold-Maxwell, J.L.; Meinke, L.J.; Gray, M.E.; Estes, R.E.; Gassmann, A.J. Effect of Bt maize and soil insecticides on yield, injury, and rootworm survival: Implications for resistance management. J. Econ. Entomol. 2013, 106, 1941–1951. [Google Scholar] [CrossRef][Green Version]
- Weiss, M.J.; Seevers, K.P.; Mayo, Z.B. Influence of western corn rootworm larval densities and damage on corn rootworm survival, developmental time, size, and sex ratio (Coleoptera: Chrysomelidae). J. Kans. Entomol. Soc. 1985, 58, 397–402. [Google Scholar]
- Elliott, N.C.; Sutter, G.R.; Branson, T.F.; Fisher, J.R. Effect of population density of immatures on survival and development of the western corn rootworm (Coleoptera: Chrysomelidae). J. Entomol. Sci. 1989, 24, 209–213. [Google Scholar] [CrossRef]
- Mayo, Z.B. Control of larvae of the western and northern corn rootworm with liquid starter fertilizer-insecticide combinations and the influence of depth of placement. J. Econ. Entomol. 1977, 70, 234–236. [Google Scholar] [CrossRef]
- Peters, L.L.; Lowry, S.R. Western corn rootworm (Coleoptera: Chrysomelidae) larval control with chlorpyrifos applied at planting time versus a post-planting chemigation application to corn growth under two different tillage systems. J. Kans. Entomol. Soc. 1991, 64, 451–454. [Google Scholar]
- USEPA. Pesticide Fact Sheet: Clothianidin. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/fs_PC-044309_30-May-03.pdf (accessed on 16 December 2020).
- Beyond Pesticides Appendix B. Thiamethoxam Pesticide Registrations. 2013. Available online: https://www.beyondpesticides.org/assets/media/documents/pollinators/documents/2013-03-21AppendixBThiamethoxamII.pdf (accessed on 16 December 2020).
- Syngenta. Syngenta: Pioneering Seed Treatment Innovation. Available online: https://www.syngenta-us.com/prodrender/imagehandler.ashx?imid=a0209de7-27cf-4e31-8d4a-10c19f3b1194&fty=0&et=8 (accessed on 16 December 2020).
- Mullin, C.A.; Saunders, M.C., II; Leslie, T.W.; Biddinger, D.J.; Fleischer, S.J. Toxic and behavioral effects to Carabidae of seed treatments used on Cry3Bb1- and Cry1Ab/c-protected corn. Environ. Entomol. 2005, 34, 1626–1636. [Google Scholar] [CrossRef]
- Magalhaes, L.C.; French, B.W.; Hunt, T.E.; Siegfried, B.D. Baseline susceptibility of western corn rootworm (Coleoptera: Chrysomelidae) to clothianidin. J. Appl. Entomol. 2007, 131, 251–255. [Google Scholar] [CrossRef]
- Steffey, K.; Gray, M.; Estes, R. Insecticidal seed treatments and soil insecticides for corn rootworm control. In Proceedings of the Illinois Crop Protection Technology Conference; University of Illinois Extension: Urbana-Champaign, IL, USA, 2005; pp. 35–40. [Google Scholar]
- Van Rozen, K.; Ester, A. Chemical control of Diabrotica virgifera virgifera LeConte. J. Appl. Entomol. 2010, 134, 376–384. [Google Scholar] [CrossRef]
- Krupke, C.H.; Long, E.Y. Intersections between neonicotinoid seed treatments and honey bees. Curr. Opin. Insect Sci. 2015, 10, 8–13. [Google Scholar] [CrossRef]
- Schaafsma, A.; Limay-Rios, V.; Baute, T.; Smith, J.; Xue, Y. Neonicotinoid insecticide residues in surface water and soil associated with commercial maize (Corn) fields in southwestern ontario. PLoS ONE 2015, 10, e0118139. [Google Scholar] [CrossRef]
- Douglas, M.R.; Rohr, J.R.; Tooker, J.F. Neonicotinoid insecticide travels through a soil food chain, disrupting biological control of non-target pests and decreasing soya bean yield. J. Appl. Ecol. 2015, 52, 250–260. [Google Scholar] [CrossRef]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef]
- European Commission. Neonicotinoids. 2020. Available online: https://ec.europa.eu/food/plant/pesticides/approval_active_substances/approval_renewal/neonicotinoids_en (accessed on 14 December 2020).
- Health Canada Update on the Neonicotinoid Pesticides. 2020. Available online: https://www.canada.ca/content/dam/hc-sc/documents/services/consumer-product-safety/reports-publications/pesticides-pest-management/fact-sheets-other-resources/update-neonicotinoid-pesticides-january-2020/neonic-update-sept2020-eng.pdf (accessed on 14 December 2020).
- USEPA. Schedule for Review of Neonicotinoid Pesticides. Available online: https://www.epa.gov/pollinator-protection/schedule-review-neonicotinoid-pesticides (accessed on 14 December 2020).
- Sappington, T.W.; Hesler, L.S.; Allen, K.C.; Luttrell, R.G.; Papiernik, S.K. Prevalence of sporadic insect pests of seedling corn and factors affecting risk of infestation. J. Integr. Pest Manag. 2018, 9. [Google Scholar] [CrossRef]
- Veres, A.; Wyckhuys, K.A.G.; Kiss, J.; Tóth, F.; Burgio, G.; Pons, X.; Avilla, C.; Vidal, S.; Razinger, J.; Bazok, R.; et al. An update of the worldwide integrated assessment (WIA) on systemic pesticides. Part 4: Alternatives in major cropping systems. Environ. Sci. Pollut. Res. 2020, 27, 29867–29899. [Google Scholar] [CrossRef] [PubMed]
- Meinke, L.J. Adult corn rootworm management. In University of Nebraska Agricultural Research Division Miscellaneous Publication 63-C; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1995. [Google Scholar]
- Meinke, L.J. Adult Corn Rootworm Suppression: Corn Rootworm Management in the Transgenic Era. Available online: http://www.plantmanagementnetwork.org/edcenter/seminars/corn/AdultRootworm/ (accessed on 20 December 2020).
- Beardmore, B.W. Evaluation of western corn rootworm, Diabrotica virgifera LeConte, adult control on an individual field basis as an alternative to the use of soil insecticides in eastern Nebraska. Master’s Thesis, University of Nebraska-Lincoln, Lincoln, NE, USA, 1975. [Google Scholar]
- Meinke, L.J.; Siegfried, B.; Wright, R.J.; Chandler, L. Adult susceptibility of Nebraska western corn rootworm (Coleoptera: Chrysomelidae) populations to selected insecticides. J. Econ. Entomol. 1998, 91, 594–600. [Google Scholar] [CrossRef]
- Wright, R.J.; Meinke, L.J.; Siegfried, B.D. Corn rootworm management. In Proceedings Crop Protection Clinics; The University of Nebraska-Lincoln: Lincoln, NE, USA, 1996; pp. 45–53. [Google Scholar]
- Meinke, L.J.; Siegfried, B.D.; Wright, R.J.; Chandler, L.D. Western corn rootworm resistance to insecticides: Current situation in Nebraska. In Proceedings of the Illinois Agricultural Pesticides Conference, Champaign, IL, USA, 8–9 January 1997; pp. 88–92. [Google Scholar]
- Siegfried, B.D.; Meinke, L.J.; Scharf, M.E. Resistance management concerns for areawide management programs. J. Agric. Entomol. 1998, 15, 359–369. [Google Scholar]
- Scharf, M.E.; Meinke, L.J.; Siegfried, B.D.; Wright, R.J.; Chandler, L.D. Carbaryl susceptibility, diagnostic concentration determination, and synergism for U.S. populations of western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1999, 92, 33–39. [Google Scholar] [CrossRef][Green Version]
- Zhou, X.; Scharf, M.E.; Parimi, S.; Meinke, L.J.; Wright, R.J.; Chandler, L.D.; Siegfried, B.D. Diagnostic assays based on esterase-mediated resistance mechanisms in western corn rootworms (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2002, 95, 1261–1266. [Google Scholar] [CrossRef][Green Version]
- Miller, N.J.; Guillemaud, T.; Giordano, R.; Siegfried, B.D.; Gray, M.E.; Meinke, L.J.; Sappington, T.W. Genes, Gene flow and adaptation of Diabrotica virgifera virgifera. Agric. For. Entomol. 2009, 11, 47–60. [Google Scholar] [CrossRef]
- Anderson, J.F.; Metcalf, R.L. Identification of a volatile attractant for Diabrotica and Acalymma spp. from blossoms of Cucurbita maxima Duchesne. J. Chem. Ecol. 1986, 12, 87–699. [Google Scholar]
- Lampman, R.L.; Metcalf, R.L.; Andersen, J.F. Semiochemical attractants of Diabrotica undecimpunctata howardi Barber, southern corn rootworm, and Diabrotica virgifera virgifera LeConte, the western corn rootworm (Coleoptera: Chrysomelidae). J. Chem. Ecol. 1987, 13, 959–975. [Google Scholar] [CrossRef]
- Lampman, R.L.; Metcalf, R.L. Multicomponent kairomonal lures for southern and western corn rootworms (Coleoptera: Chrysomelidae: Diabrotica spp.). J. Econ. Entomol. 1987, 80, 1137–1142. [Google Scholar] [CrossRef]
- Chambliss, O.L.; Jones, C.M. Cucurbitacins: Specific insect attractants in Cucurbitaceae. Science 1966, 153, 1392–1393. [Google Scholar] [CrossRef]
- Metcalf, R.L. Changing role of insecticides in crop protection. Ann. Rev. Entomol. 1980, 25, 219–256. [Google Scholar] [CrossRef]
- Metcalf, R.L.; Rhodes, A.M.; Metcalf, R.A.; Ferguson, J.; Metcalf, E.R.; Lu, P.-Y. Cucurbitacin contents and diabroticite (Coleoptera: Chrysomelidae) feeding upon Cucurbita spp. Environ. Entomol. 1982, 11, 931–937. [Google Scholar] [CrossRef]
- Metcalf, R.L.; Ferguson, J.E.; Lampman, R.; Andersen, J.F. Dry cucurbitacin-containing baits for controlling diabroticite beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1987, 80, 870–875. [Google Scholar] [CrossRef]
- Lance, D.R.; Sutter, G.R. Field-cage and laboratory evaluations of semiochemical-based baits for managing western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1990, 83, 1085–1090. [Google Scholar] [CrossRef]
- Weissling, T.J.; Meinke, L.J. Potential of starch encapsulated semiochemical-insecticide formulations for adult corn rootworm (Coleoptera: Chrysomelidae) control. J. Econ. Entomol. 1991, 84, 601–609. [Google Scholar] [CrossRef][Green Version]
- Lance, D.R.; Sutter, G.R. Field tests of a semiochemical-based toxic bait for suppression of corn rootworm beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1992, 85, 967–973. [Google Scholar] [CrossRef]
- Weissling, T.J.; Meinke, L.J. Semiochemical-insecticide bait placement and vertical distribution of corn rootworm (Coleoptera: Chrysomelidae) adults: Implications for management. Environ. Entomol. 1991, 20, 945–952. [Google Scholar] [CrossRef]
- Lance, D.R.; Sutter, G.R. Semiochemical-based toxic baits for Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae): Effects of particle size, location, and attractant content. J. Econ. Entomol. 1991, 84, 1861–1868. [Google Scholar] [CrossRef]
- Meinke, L.J. Behavior-based adult corn rootworm management. In Proceedings of the 1992 Crop Pest Management Update; University of Illinois Extension: Urbana-Champaign, IL, USA, 1992; pp. 61–65. [Google Scholar]
- Chandler, L.D. Comparison of insecticide-bait aerial application methods for management of corn rootworm (Coleoptera: Chrysomelidae). Southwest. Entomol. 1998, 23, 147–159. [Google Scholar]
- Wilde, G.E.; Whitworth, R.J.; Shufran, R.A.; Zhu, K.Y.; Sloderbeck, P.E.; Higgins, R.A.; Buschman, L.L. Rootworm areawide management project in Kansas. J. Agric. Entomol. 1998, 15, 335–349. [Google Scholar]
- Behle, R.W. Consumption of residue containing cucurbitacin feeding stimulant and reduced rates of carbaryl insecticide by western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2001, 94, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Chandler, L.D. Corn rootworm areawide management program: United States Department of Agriculture-Agricultural Research Service. Pest Manag. Sci. 2003, 59, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Chandler, L.D.; Coppedge, J.R.; Edwards, C.R.; Tollefson, J.J.; Wilde, G.R.; Faust, R.M. Corn rootworm areawide pest management in the midwestern USA. In Areawide Pest Management: Theory and Implementation; Koul, O., Cuperous, G., Elliott, N., Eds.; CAB International: Oxfordshire, UK, 2008; pp. 191–207. [Google Scholar]
- Boetel, M.A.; Fuller, B.W.; Chandler, L.D.; Tollefson, J.J.; Mcmanus, B.L.; Kadakia, N.D.; Evenson, P.D.; Mishra, T.P. Nontarget arthropod abundance in areawide-managed corn habitats treated with semiochemical-based bait insecticide for corn rootworm (Coleoptera: Chrysomelidae) control. J. Econ. Entomol. 2005, 98, 1957–1968. [Google Scholar] [CrossRef]
- Weissling, T.J.; Meinke, L.J.; Lytle, K.A. Effect of starch-based corn rootworm (Coleoptera: Chrysomelidae) baits on selected nontarget insect species: Influence of semiochemical composition. J. Econ. Entomol. 1991, 84, 1235–1241. [Google Scholar] [CrossRef][Green Version]
- Siegfried, B.D.; Meinke, L.J.; Parimi, S.; Scharf, M.E.; Nowatzki, T.J.; Zhou, X.; Chandler, L.D. Monitoring western corn rootworm (Coleoptera: Chrysomelidae) susceptibility to carbaryl and cucurbitacin baits in the areawide management pilot program. J. Econ. Entomol. 2004, 97, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.Y.; Wilde, G.E.; Higgins, R.A.; Sloderbeck, P.E.; Buschman, L.L.; Shufran, R.A.; Whitworth, R.J.; Starkey, S.R.; He, F. Evidence of evolving carbaryl resistance in western corn rootworm (Coleoptera: Chrysomelidae) in areawide-managed cornfields in north central Kansas. J. Econ. Entomol. 2001, 94, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Parimi, S.; Meinke, L.J.; Nowatzki, T.M.; Chandler, L.D.; Wade French, B.; Siegfried, B.D. Toxicity of insecticide-bait mixtures to insecticide resistant and susceptible western corn rootworms (Coleoptera: Chrysomelidae). Crop Prot. 2003, 22, 781–786. [Google Scholar] [CrossRef]
- Chandler, L.D.; Faust, R.M. Overview of areawide management of insects. J. Agric. Entomol. 1998, 15, 319–325. [Google Scholar]
- Rogers, E.M. Diffusion of Innovations, 3rd ed.; Free Press: New York, NY, USA, 1983. [Google Scholar]
- Rice, M.E. Transgenic rootworm corn: Assessing potential agronomic, economic, and environmental benefits. Plant Health Prog. 2004. [Google Scholar] [CrossRef]
- Souza, D.; Vieira, B.C.; Fritz, B.K.; Hoffmann, W.C.; Peterson, J.A.; Kruger, G.R.; Meinke, L.J. Western corn rootworm pyrethroid resistance confirmed by aerial application simulations of commercial insecticides. Sci. Rep. 2019, 9, 6713. [Google Scholar] [CrossRef]
- Pereira, A.E.; Wang, H.; Zukoff, S.N.; Meinke, L.J.; French, B.W.; Siegfried, B.D. Evidence of field-evolved resistance to bifenthrin in western corn rootworm (Diabrotica virgifera virgifera LeConte) populations in western Nebraska and Kansas. PLoS ONE 2015, 10, e0142299. [Google Scholar] [CrossRef] [PubMed]
- NASS-USDA. QuickStats Ad-Hoc Query Tool. Available online: https://quickstats.nass.usda.gov/ (accessed on 20 December 2020).
- Pereira, A.E.; Souza, D.; Zukoff, S.N.; Meinke, L.J.; Siegfried, B.D. Cross-resistance and synergism bioassays suggest multiple mechanisms of pyrethroid resistance in western corn rootworm populations. PLoS ONE 2017, 12, e0179311. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.J.; Scharf, M.E.; Meinke, L.J.; Zhou, X.; Siegfried, B.D.; Chandler, L.D. Larval susceptibility of an insecticide-resistant western corn rootworm (Coleoptera: Chrysomelidae) population to soil insecticides: Laboratory bioassays, assays of detoxification enzymes, and field performance. J. Econ. Entomol. 2000, 93, 7–13. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chio, H.; Metcalf, R.L. Detoxication mechanisms for aldrin, carbofuran, fonofos, phorate, and terbufos in four species of diabroticites. J. Econ. Entomol. 1979, 72, 732–738. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Mullin, C.A. Properties of a cytochrome P-450-dependent epoxidase in aldrin-resistant western corn rootworms, Diabrotica virgifera virgifera LeConte. Pestic. Biochem. Physiol. 1988, 31, 261–268. [Google Scholar] [CrossRef]
- Hamilton, E.W. Relative toxicity of aldrin and dieldrin. J. Agric. Food Chem. 1971, 19, 863–864. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Mullin, C.A. Metabolism, penetration, and partitioning of [14C]aldrin in aldrin-resistant and susceptible corn rootworms. Pestic. Biochem. Physiol. 1990, 36, 135–146. [Google Scholar] [CrossRef]
- Kadous, A.A.; Ghiasuddin, S.M.; Matsumura, F.; Scott, J.G.; Tanaka, K. Difference in the picrotoxinin receptor between the cyclodiene-resistant and susceptible strains of the german cockroach. Pestic. Biochem. Physiol. 1983, 19, 157–166. [Google Scholar] [CrossRef]
- Tanaka, K.; Scott, J.G.; Matsumura, F. Picrotoxinin receptor in the central nervous system of the american cockroach: Its role in the action of cyclodiene-type insecticides. Pestic. Biochem. Physiol. 1984, 22, 117–127. [Google Scholar] [CrossRef]
- Bloomquist, J.R.; Soderlund, D.M. Neurotoxic insecticides inhibit GABA-dependent chloride uptake by mouse brain vesicles. Biochem. Biophys. Res. Commun. 1985, 133, 37–43. [Google Scholar] [CrossRef]
- Narahashi, T.; Frey, J.M.; Ginsburg, K.S.; Roy, M.L. Sodium and GABA-activated channels as the targets of pyrethroids and cyclodienes. Toxicol. Lett. 1992, 64–65, 429–436. [Google Scholar] [CrossRef]
- Buckingham, S.D.; Biggin, P.C.; Sattelle, B.M.; Brown, L.A.; Sattelle, D.B. Insect GABA receptors: Splicing, editing, and targeting by antiparasitics and insecticides. Mol. Pharmacol. 2005, 68, 942–951. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Williamson, M.S.; Davies, T.G.E.; Bass, C. Ion channels as insecticide targets. J. Neurosci. 2016, 30, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Anthony, N.; Aronstein, K.; Rocheleau, T.; Stilwell, G. Cyclodiene insecticide resistance: From molecular to population genetics. Ann. Rev. Entomol. 2000, 45, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Raymond-Delpech, V.; Matsuda, K.; Sattelle, B.M.; Rauh, J.J.; Sattelle, D.B. Ion channels: Molecular targets of neuroactive insecticides. Invertebr. Neurosci. 2005, 3–4, 119–133. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Roush, R.T. Cloning of a locus associated with cyclodiene resistance in Drosophila. In Molecular Mechanisms of Insecticide Resistance; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 1992; Volume 505, pp. 90–98. ISBN 978-0-8412-2474-2. [Google Scholar]
- Taylor-Wells, J.; Jones, A.K. Variations in the insect GABA receptor, RDL, and their impact on receptor pharmacology. In Advances in Agrochemicals: Ion Channels and G Protein-Coupled Receptors (GPCRs) as Targets for Pest Control; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2017; Volume 1265, pp. 1–21. ISBN 978-0-8412-3260-0. [Google Scholar]
- Hosie, A.; Sattelle, D.; Aronstein, K.; Ffrench-Constant, R. Molecular biology of insect neuronal GABA receptors. Trends Neurosci. 1997, 20, 578–583. [Google Scholar] [CrossRef]
- Wang, H.; Coates, B.S.; Chen, H.; Sappington, T.W.; Guillemaud, T.; Siegfried, B.D. Role of a gamma-aminobutryic acid (GABA) receptor mutation in the evolution and spread of Diabrotica virgifera virgifera resistance to cyclodiene insecticides. Insect. Mol. Biol. 2013, 22, 473–484. [Google Scholar] [CrossRef]
- Chen, H.; Wang, H.; Siegfried, B.D. Genetic differentiation of western corn rootworm populations (Coleoptera: Chrysomelidae) relative to insecticide resistance. Ann. Entomol. Soc. Am. 2012, 105, 232–240. [Google Scholar] [CrossRef][Green Version]
- Coustau, C.; Chevillon, C.; Ffrench-Constant, R. Resistance to xenobiotics and parasites: Can we count the cost? Trends Ecol. Evol. 2000, 15, 378–383. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Bass, C. Does resistance really carry a fitness cost? Curr. Opin. Insect Sci. 2017, 21, 39–46. [Google Scholar] [CrossRef]
- Taylor, M.; Feyereisen, R. Molecular biology and evolution of resistance of toxicants. Mol. Biol. Evol. 1996, 13, 719–734. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mckenzie, J.A. Selection at the dieldrin resistance locus in overwintering populations of Lucilia cuprina (Wiedemann). Aust. J. Zool. 1990, 38, 493–501. [Google Scholar] [CrossRef]
- Rowland, M. Activity and mating competitiveness of ΓHCH/dieldrin resistant and susceptible male and virgin female Anopheles gambiae and An. stephensi mosquitoes, with assessment of an insecticide-rotation strategy. Med. Vet. Entomol. 1991, 5, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Aronstein, K.; Ode, P.; Ffrench-Constant, R.H. Direct comparison of PCR-based monitoring for cyclodiene resistance in Drosophila populations with insecticide bioassay. Pestic. Biochem. Physiol. 1994, 48, 229–233. [Google Scholar] [CrossRef]
- Hansen, K.K.; Kristensen, M.; Jensen, K.-M.V. Correlation of a resistance-associated Rdl mutation in the german cockroach, Blattella germanica (L), with persistent dieldrin resistance in two Danish field populations. Pest Manag. Sci. 2005, 61, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowska, R.M.; Platt, N.; Poupardin, R.; Irving, H.; Dabire, R.K.; Mitchell, S.; Jones, C.M.; Diabaté, A.; Ranson, H.; Wondji, C.S. Dissecting the mechanisms responsible for the multiple insecticide resistance phenotype in Anopheles gambiae s.s., M Form, from Vallée Du Kou, Burkina Faso. Gene 2013, 519, 98–106. [Google Scholar] [CrossRef]
- Aldridge, W.N. Some properties of specific cholinesterase with particular reference to the mechanism of inhibition by diethyl p-nitrophenyl thiophosphate (E 605) and analogues. Biochem. J. 1950, 46, 451–460. [Google Scholar] [CrossRef]
- Aldridge, W.N.; Reiner, E. Enzyme Inhibitors as Substrates: Interactions of Esterases with Esters of Organophosphorus and Carbamic Acids; North-Holland Publishing Co.: Amsterdam, The Netherland, 1972. [Google Scholar]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Čolović, M.B.; Krstić, D.Z.; Lazarević-Pašti, T.D.; Bondžić, A.M.; Vasić, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Montella, I.R.; Schama, R.; Valle, D. The classification of esterases: An important gene family involved in insecticide resistance—A Review. Mem. Inst. Oswaldo Cruz 2012, 107, 437–449. [Google Scholar] [CrossRef]
- Sogorb, M.A.; Vilanova, E. Enzymes involved in the detoxification of organophosphorus, carbamate and pyrethroid insecticides through hydrolysis. Toxicol. Lett. 2002, 128, 215–228. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Ann. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Fournier, D. Mutations of acetylcholinesterase which confer insecticide resistance in insect populations. Chem. Biol. Interact. 2005, 157–158, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kim, Y.H.; Kwon, D.H.; Cha, D.J.; Kim, J.H. Mutation and duplication of arthropod acetylcholinesterase: Implications for pesticide resistance and tolerance. Pestic. Biochem. Physiol. 2015, 120, 118–124. [Google Scholar] [CrossRef]
- Miota, F.; Scharf, M.E.; Ono, M.; Marçon, P.; Meinke, L.J.; Wright, R.J.; Chandler, L.D.; Siegfried, B.D. Mechanisms of methyl and ethyl parathion resistance in the western corn rootworm (Coleoptera: Chrysomelidae). Pestic. Biochem. Physiol. 1998, 61, 39–52. [Google Scholar] [CrossRef]
- Scharf, M.E.; Meinke, L.J.; Wright, R.J.; Chandler, L.D.; Siegfried, B.D. Metabolism of carbaryl by insecticide-resistant and -susceptible western corn rootworm populations (Coleoptera: Chrysomelidae). Pesti. Biochem. Physiol. 1999, 63, 85–96. [Google Scholar] [CrossRef]
- Scharf, M.E.; Parimi, S.; Meinke, L.J.; Chandler, L.D.; Siegfried, B.D. Expression and induction of three family 4 cytochrome P450 (CYP4)* genes identified from insecticide-resistant and susceptible western corn rootworms, Diabrotica virgifera virgifera. Insect Mol. Biol. 2001, 10, 139–146. [Google Scholar] [CrossRef]
- Zhou, X.; Scharf, M.E.; Meinke, L.J.; Chandler, L.D.; Siegfried, B.D. Characterization of general esterases from methyl parathion-resistant and -susceptible populations of western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2003, 96, 1855–1863. [Google Scholar] [CrossRef][Green Version]
- Zhou, X.; Scharf, M.E.; Sarath, G.; Meinke, L.J.; Chandler, L.D.; Siegfried, B.D. Partial purification and characterization of a methyl-parathion resistance-associated general esterase in Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). Pestic. Biochem. Physiol. 2004, 78, 114–125. [Google Scholar] [CrossRef]
- Parimi, S.; Scharf, M.E.; Meinke, L.J.; Chandler, L.D.; Siegfried, B.D. Inheritance of methyl-parathion resistance in Nebraska western corn rootworm populations (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2003, 96, 131–136. [Google Scholar] [CrossRef]
- Narahashi, T. Nerve membrane as a target of pyrethroids. Pestic. Sci. 1976, 7, 267–272. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Bloomquist, J.R. Neurotoxic actions of pyrethroid insecticides. Ann. Rev. Entomol. 1989, 34, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Vijverberg, H.P.M.; van der Zalm, J.M.; van den Bercken, J. Similar mode of action of pyrethroids and DDT on sodium channel gating in myelinated nerves. Nature 1982, 295, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Breckenridge, C.B.; Holden, L.; Sturgess, N.; Weiner, M.; Sheets, L.; Sargent, D.; Soderlund, D.M.; Choi, J.-S.; Symington, S.; Clark, J.M.; et al. Evidence for a separate mechanism of toxicity for the Type I and the Type II pyrethroid insecticides. Neurotoxicology 2009, 30, S17–S31. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, M.E.; McRory, J.E.; Snutch, T.P.; Stea, A. Mammalian voltage-gated calcium channels are potently blocked by the pyrethroid insecticide allethrin. J. Pharmacol. Exp. Ther. 2004, 308, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, L.J.; Casida, J.E. Stereospecific action of pyrethroid insecticides on the gamma-aminobutyric acid receptor-ionophore complex. Science 1983, 221, 1399–1401. [Google Scholar] [CrossRef] [PubMed]
- Ray, D.E.; Sutharsan, S.; Forshaw, P.J. Actions of pyrethroid insecticides on voltage-gated chloride channels in neuroblastoma cells. Neurotoxicology 1997, 18, 755–760. [Google Scholar]
- Soderlund, D.M. Molecular mechanisms of pyrethroid insecticide neurotoxicity: Recent advances. Arch. Toxicol. 2011, 86, 165–181. [Google Scholar] [CrossRef]
- Symington, S.B.; Clark, J.M. Action of deltamethrin on N-type (Cav2.2) voltage-sensitive calcium channels in rat brain. Pestic. Biochem. Physiol. 2005, 82, 1–15. [Google Scholar] [CrossRef]
- Vijverberg, H.P.M.; van den Bercken, J. Neurotoxicological effects and the mode of action of pyrethroid insecticides. Crit. Rev. Toxicol. 1990, 21, 105–126. [Google Scholar] [CrossRef]
- Gammon, D.W.; Brown, M.A.; Casida, J.E. Two classes of pyrethroid action in the cockroach. Pestic. Biochem. Physiol. 1981, 15, 181–191. [Google Scholar] [CrossRef]
- Lawrence, L.J.; Casida, J.E. Pyrethroid toxicology: Mouse intracerebral structure-toxicity relationships. Pestic. Biochem. Physiol. 1982, 18, 9–14. [Google Scholar] [CrossRef]
- Scott, J.G.; Matsumura, F. Evidence for two types of toxic actions of pyrethroids on susceptible and DDT-resistant german cockroaches. Pestic. Biochem. Physiol. 1983, 19, 141–150. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Xu, Q.; Zhu, F.; Zhang, L. Pyrethroid resistance in mosquitoes. Insect Sci. 2006, 13, 159–166. [Google Scholar] [CrossRef]
- Soderlund, D.M.; Knipple, D.C. The molecular biology of knockdown resistance to pyrethroid insecticides. Insect Biochem. Mol. Biol. 2003, 33, 563–577. [Google Scholar] [CrossRef]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; Mccarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med. Vet. Entomol. 2003, 17, 87–94. [Google Scholar] [CrossRef]
- Schuler, T.H.; Martinez-Torres, D.; Thompson, A.J.; Denholm, I.; Devonshire, A.L.; Duce, I.R.; Williamson, M.S. Toxicological, electrophysiological, and molecular characterisation of knockdown resistance to pyrethroid insecticides in the diamondback moth, Plutella Xylostella (L.). Pestic. Biochem. Physiol. 1998, 59, 169–182. [Google Scholar] [CrossRef]
- Williamson, M.S.; Denholm, I.; Bell, C.A.; Devonshire, A.L. Knockdown resistance (kdr) to DDT and pyrethroid insecticides maps to a sodium channel gene locus in the housefly (Musca domestica). Mol. Gen. Genet. 1993, 240, 17–22. [Google Scholar] [CrossRef]
- Li, F.-G.; Ai, G.-M.; Zou, D.-Y.; Ji, Y.; Bao-Gen, G.; Gao, X.-W. Characterization of activation metabolism activity of indoxacarb in insects by liquid chromatography-triple quadrupole mass spectrometry. Chin. J. Anal. Chem. 2014, 42, 463–468. [Google Scholar] [CrossRef]
- Wing, K.D.; Sacher, M.; Kagaya, Y.; Tsurubuchi, Y.; Mulderig, L.; Connair, M.; Schnee, M. Bioactivation and mode of action of the oxadiazine indoxacarb in insects. Crop Prot. 2000, 19, 537–545. [Google Scholar] [CrossRef]
- Wing, K.D.; Schnee, M.E.; Sacher, M.; Connair, M. A novel oxadiazine insecticide is bioactivated in lepidopteran larvae. Arch. Insect Biochem. Physiol. 1998, 37, 91–103. [Google Scholar] [CrossRef]
- Bisset, J.; Rodriguez, M.; Soca, A.; Pasteur, N.; Raymond, M. Cross-resistance to pyrethroid and organophosphorus insecticides in the southern house mosquito (Diptera: Culicidae) from Cuba. J. Med. Entomol. 1997, 34, 244–246. [Google Scholar] [CrossRef]
- Cahill, M.; Byrne, F.J.; Gorman, K.; Denholm, I.; Devonshire, A.L. Pyrethroid and organophosphate resistance in the tobacco whitefly Bemisia tabaci (Homoptera: Aleyrodidae). Bull. Entomol. Res. 1995, 85, 181–187. [Google Scholar] [CrossRef]
- Devonshire, A.L.; Moores, G.D. A carboxylesterase with broad substrate specificity causes organophosphorus, carbamate and pyrethroid resistance in peach-potato aphids (Myzus persicae). Pestic. Biochem. Physiol. 1982, 18, 235–246. [Google Scholar] [CrossRef]
- Gunning, R.V.; Devonshire, A.L. Negative cross resistance between indoxacarb and pyrethroids in the cotton bollworm, Helicoverpa armigera, in Australia: A tool for resistance management. In Proceedings of the BCPC International Congress, Glasgow, Scotland, UK, 10–12 November 2003; Volume 2, pp. 789–794. [Google Scholar]
- Ramasubramanian, T.; Regupathy, A. Pattern of cross-resistance in pyrethroid-selected populations of Helicoverpa armigera Hübner (Lep., Noctuidae) from India. J. Appl. Entomol. 2004, 128, 583–587. [Google Scholar] [CrossRef]
- Souza, D.; Jiménez, A.V.; Sarath, G.; Meinke, L.J.; Miller, N.J.; Siegfried, B.D. Enhanced metabolism and selection of pyrethroid-resistant western corn rootworms (Diabrotica virgifera virgifera LeConte). Pestic. Biochem. Physiol. 2020, 164, 165–172. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Taylor, C.E. Genetic and biological influences in the evolution of insecticide resistance. J. Econ. Entomol. 1977, 70, 319–323. [Google Scholar] [CrossRef]
- Roush, R.T.; McKenzie, J.A. Ecological genetics of insecticide and acaricide resistance. Ann. Rev. Entomol. 1987, 32, 361–380. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Resistance risk assessment: Realized heritability of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae), tobacco budworm (Lepidoptera: Noctuidae), and Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1992, 85, 1551–1559. [Google Scholar] [CrossRef]
- Souza, D.; Siegfried, B.D.; Meinke, L.J.; Miller, N.J. Molecular characterization of western corn rootworm pyrethroid resistance. Pest Manag. Sci. 2021, 77, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Crowder, D.W.; Onstad, D.W.; Gray, M.E.; Pierce, C.M.F.; Hager, A.G.; Ratcliffe, S.T.; Steffey, K.L. Analysis of the dynamics of adaptation to transgenic corn and crop rotation by western corn rootworm (Coleoptera: Chrysomelidae) using a daily time-step model. J. Econ. Entomol. 2005, 98, 534–551. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Clark, T.L.; Frank, D.L.; French, B.W.; Meinke, L.J.; Moellenbeck, D.; Vaughn, T.T.; Hibbard, B.E. Mortality impact of MON863 transgenic maize roots on western corn rootworm larvae in the field. J. Appl. Entomol. 2012, 136, 721–729. [Google Scholar] [CrossRef]
- Hitchon, A.J.; Smith, J.L.; French, B.W.; Schaafsma, A.W. Impact of the Bt corn proteins Cry34/35Ab1 and Cry3Bb1, alone or pyramided, on western corn rootworm (Coleoptera: Chrysomelidae) beetle emergence in the field. J. Econ. Entomol. 2015, 108, 1986–1993. [Google Scholar] [CrossRef]
- Keweshan, R.S.; Head, G.P.; Gassmann, A.J. Effects of pyramided Bt corn and blended refuges on western corn rootworm and northern corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2015, 108, 720–729. [Google Scholar] [CrossRef]
- Shields, E.J.; Testa, A.M.; O’Neil, W.J. Long-term persistence of native New York entomopathogenic nematode isolates across crop rotation. J. Econ. Entomol. 2018, 111, 2592–2598. [Google Scholar] [CrossRef]
- Shields, E.J. Biological Control of Corn Rootworm with Persistent Entomopathogenic Nematodes: An Opportunity to Try Them on Your Farm. Available online: https://blogs.cornell.edu/whatscroppingup/2019/03/22/biological-control-of-corn-rootworm-with-persistent-entomopathogenic-nematodes-an-opportunity-to-try-them-on-your-farm/ (accessed on 31 December 2020).
- USEPA. Framework to Delay Corn Rootworm Resistance. Available online: https://www.epa.gov/regulation-biotechnology-under-tsca-and-fifra/framework-delay-corn-rootworm-resistance (accessed on 16 December 2020).
- Levine, E.; Spencer, J.L.; Isard, S.A.; Onstad, D.W.; Gray, M.E. Adaptation of the western corn rootworm, Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae), to crop rotation: Evolution of a new strain in response to a cultural management practice. Am. Entomol. 2002, 48, 94–107. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Clifton, E.H.; Dunbar, M.W.; Hoffmann, A.M.; Ingber, D.A.; Keweshan, R.S. Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. Proc. Natl. Acad. Sci. USA 2014, 111, 5141–5146. [Google Scholar] [CrossRef]
- Reinders, J.D.; Hitt, B.D.; Stroup, W.W.; French, B.W.; Meinke, L.J. Spatial variation in western corn rootworm (Coleoptera: Chrysomelidae) susceptibility to Cry3 toxins in Nebraska. PLoS ONE 2018, 13, e0208266. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meinke, L.J.; Souza, D.; Siegfried, B.D. The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms. Insects 2021, 12, 112. https://doi.org/10.3390/insects12020112
Meinke LJ, Souza D, Siegfried BD. The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms. Insects. 2021; 12(2):112. https://doi.org/10.3390/insects12020112
Chicago/Turabian StyleMeinke, Lance J., Dariane Souza, and Blair D. Siegfried. 2021. "The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms" Insects 12, no. 2: 112. https://doi.org/10.3390/insects12020112
APA StyleMeinke, L. J., Souza, D., & Siegfried, B. D. (2021). The Use of Insecticides to Manage the Western Corn Rootworm, Diabrotica virgifera virgifera, LeConte: History, Field-Evolved Resistance, and Associated Mechanisms. Insects, 12(2), 112. https://doi.org/10.3390/insects12020112