Large-Scale Monitoring of Resistance to Coumaphos, Amitraz, and Pyrethroids in Varroa destructor


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mites
2.2. Bioassays with Acaricides
2.3. TaqMan Assays
2.4. Statistical Analysis
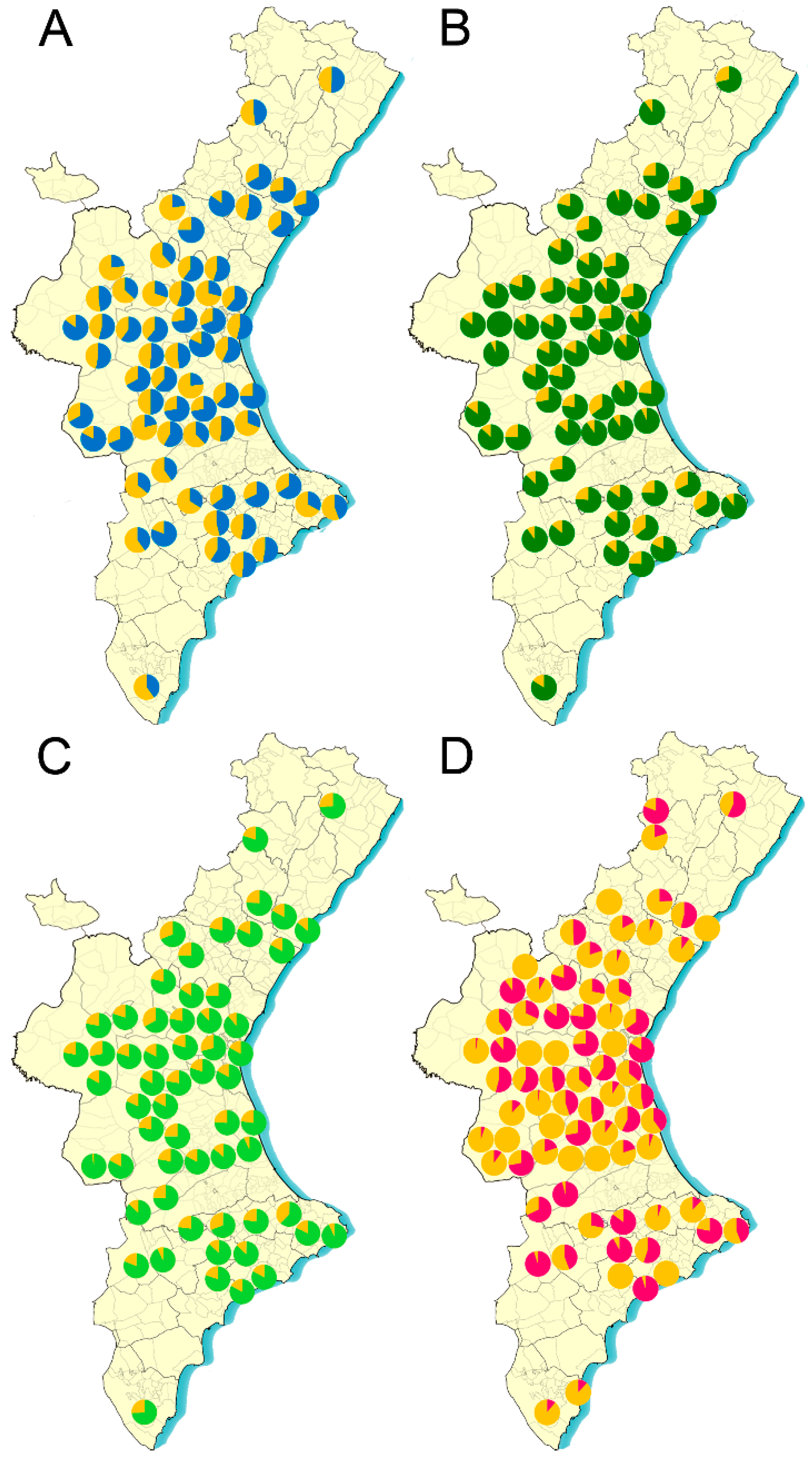
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Boecking, O.; Genersch, E. Varroosis—The ongoing crisis in bee keeping. J. Consum. Prot. Food Saf. 2008, 3, 221–228. [Google Scholar] [CrossRef]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E.; Von Der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The German bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Eccles, L.; Calvete, Y.; Mcgowan, J.; Kelly, P.G.; Correa-Benítez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2010, 41, 443–450. [Google Scholar] [CrossRef]
- Solignac, M.; Cornuet, J.M.; Vautrin, D.; Le Conte, Y.; Anderson, D.; Evans, J.; Cros-Arteil, S.; Navajas, M. The invasive Korea and Japan types of Varroa destructor, ectoparasitic mites of the Western honeybee (Apis mellifera), are two partly isolated clones. Proc. R. Soc. B-Biol. Sci. 2005, 272, 411–419. [Google Scholar] [CrossRef]
- Llorente, J. Principales Enfermedades de las Abejas; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2003. [Google Scholar]
- Muñoz, I.; Garrido-Bailón, E.; Martín-Hernández, R.; Meana, A.; Higes, M.; De la Rúa, P. Genetic profile of Varroa destructor infesting Apis mellifera iberiensis colonies. J. Apic. Res. 2008, 47, 310–313. [Google Scholar] [CrossRef]
- Underwood, R.; López-Uribe, M. Methods to Control. Varroa Mites: An. Integrated Pest. Management Approach; Penn State University: State College, PA, USA, 2020. [Google Scholar]
- FAO Specifications for Agricultural Pesticides in Agriculture. Available online: http://www.fao.org/agriculture/crops/core-themes/theme/pests/jmps/ps-new/en/ (accessed on 5 August 2020).
- Elzen, P.J.; Eischen, F.A.; Baxter, J.B.; Pettis, J.; Elzen, G.W.; Wilson, W.T. Fluvalinate resistance in Varroa jacobsoni from several geographic locations. Am. Bee J. 1998, 138, 674–676. [Google Scholar]
- González-Cabrera, J.; Bumann, H.; Rodríguez-Vargas, S.; Kennedy, P.J.; Krieger, K.; Altreuther, G.; Hertel, A.; Hertlein, G.; Nauen, R.; Williamson, M.S. A single mutation is driving resistance to pyrethroids in European populations of the parasitic mite, Varroa destructor. J. Pest Sci. 2018, 91, 1137–1144. [Google Scholar] [CrossRef]
- Gracia-Salinas, M.J.; Ferrer-Dufol, M.; Latorre-Castro, E.; Monero-Manera, C.; Castillo-Hernández, J.A.; Lucientes-Curd, J.; Peribanez-Lopez, M.A. Detection of fluvalinate resistance in Varroa destructor in Spanish apiaries. J. Apic. Res. 2006, 45, 101–105. [Google Scholar] [CrossRef]
- Milani, N. The resistance of Varroa-Jacobsoni Oud to pyrethroids—A laboratory assay. Apidologie 1995, 26, 415–429. [Google Scholar] [CrossRef]
- Sammataro, D.; Untalan, P.; Guerrero, F.; Finley, J. The resistance of varroa mites (Acari: Varroidae) to acaricides and the presence of esterase. Int. J. Acarol. 2005, 31, 67–74. [Google Scholar] [CrossRef]
- Elzen, P.J.; Westervelt, D. Detection of coumaphos resistance in Varroa destructor in Florida. Am. Bee J. 2002, 142, 291–292. [Google Scholar]
- Maggi, M.D.; Ruffinengo, S.R.; Damiani, N.; Sardella, N.H.; Eguaras, M.J. First detection of Varroa destructor resistance to coumaphos in Argentina. Exp. Appl. Acarol. 2009, 47, 317–320. [Google Scholar] [CrossRef]
- Maggi, M.D.; Ruffinengo, S.R.; Mendoza, Y.; Ojeda, P.; Ramallo, G.; Floris, I.; Eguaras, M.J. Susceptibility of Varroa destructor (Acari: Varroidae) to synthetic acaricides in Uruguay: Varroa mites’ potential to develop acaricide resistance. Parasitol. Res. 2011, 108, 815–821. [Google Scholar] [CrossRef]
- Kamler, M.; Nesvorna, M.; Stara, J.; Erban, T.; Hubert, J. Comparison of tau-fluvalinate, acrinathrin, and amitraz effects on susceptible and resistant populations of Varroa destructor in a vial test. Exp. Appl. Acarol. 2016, 69, 1–9. [Google Scholar] [CrossRef]
- Maggi, M.D.; Ruffinengo, S.R.; Negri, P.; Eguaras, M.J. Resistance phenomena to amitraz from populations of the ectoparasitic mite Varroa destructor of Argentina. Parasitol. Res. 2010, 107, 1189–1192. [Google Scholar] [CrossRef]
- Rinkevich, F.D. Detection of amitraz resistance and reduced treatment efficacy in the Varroa Mite, Varroa destructor, within commercial beekeeping operations. PLoS ONE 2020, 15, e0227264. [Google Scholar] [CrossRef]
- Rodríguez-Dehaibes, S.R.; Otero-Colina, G.; Sedas, V.P.; Jiménez, J.A.V. Resistance to amitraz and flumethrin in Varroa destructor populations from Veracruz, Mexico. J. Apic. Res. 2005, 44, 124–125. [Google Scholar] [CrossRef]
- González-Cabrera, J.; Davies, T.E.; Field, L.M.; Kennedy, P.J.; Williamson, M.S. An amino acid substitution (L925V) associated with resistance to pyrethroids in Varroa destructor. PLoS ONE 2013, 8, e82941. [Google Scholar] [CrossRef]
- González-Cabrera, J.; Rodriguez-Vargas, S.; Davies, T.E.; Field, L.M.; Schmehl, D.; Ellis, J.D.; Krieger, K.; Williamson, M.S. Novel Mutations in the voltage-gated sodium channel of pyrethroid-resistant Varroa destructor populations from the Southeastern USA. PLoS ONE 2016, 11, e0155332. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.; Nesvorna, M.; Kamler, M.; Kopecky, J.; Tyl, J.; Titera, D.; Stara, J. Point mutations in the sodium channel gene conferring tau-fluvalinate resistance in Varroa destructor. Pest Manag. Sci. 2014, 70, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Hernández-Rodríguez, C.S.; González-Cabrera, J. Assessing the resistance to acaricides in Varroa destructor from several Spanish locations. Parasitol. Res. 2020, 119, 3595–3601. [Google Scholar] [CrossRef] [PubMed]
- Püntener, W. Manual for Field Trials in Plant Protection Second Edition; Ciba-Geigy Limited: Basle, Switzerland, 1981. [Google Scholar]
- Davies, T.G.E.; Field, L.M.; Usherwood, P.N.R.; Williamson, M.S. DDT, pyrethrins, pyrethroids and insect sodium channels. IUBMB Life 2007, 59, 151–162. [Google Scholar] [CrossRef]
- Calatayud, F.; Simó, E.; Domingo, P. Hacia un control integrado y sostenible de Varroa (1). Apic. Ibérica 2018, 27, 15–25. [Google Scholar]
- Sánchez Escudero, M.D.; Fernández Tejedor, T. Lucha Frente a la Varroosis en Colmenas Layens; CSIC—Instituto de Recursos Naturales y Agrobiología de Salamanca (IRNASA): Salamanca, Spain, 2016. [Google Scholar]
- Mitton, G.A.; Szawarski, N.; Ramos, F.; Fuselli, S.; Meroi Arcerito, F.R.; Eguaras, M.J.; Ruffinengo, S.R.; Maggi, M.D. Varroa destructor: When reversion to coumaphos resistance does not happen. J. Apic. Res. 2018, 57, 536–540. [Google Scholar] [CrossRef]
- Calatayud-Vernich, P.; Calatayud, F.; Simó, E.; Picó, Y. Pesticide residues in honey bees, pollen and beeswax: Assessing beehive exposure. Environ. Pollut. 2018, 241, 106–114. [Google Scholar] [CrossRef]
- Milani, N.; Della Vedova, G. Decline in the proportion of mites resistant to fluvalinate in a population of Varroa destructor not treated with pyrethroids. Apidologie 2002, 33, 417–422. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sanitary Defence Groups (ADS) | Province | No. Apiaries Sampled in 2018 | No. Apiaries Sampled in 2019 |
|---|---|---|---|
| ADSAV | Valencia | 6 | 3 |
| AIXAM | Castellón | 5 | 4 |
| ALAPI | Alicante | 2 | 1 |
| APAC | Castellón | 17 | 13 |
| APIADS | Valencia | 23 | 27 |
| APICAL | Alicante | 15 | 12 |
| APIVAL | Valencia | 9 | 13 |
| CASAPI | Castellón | 5 | 4 |
| PROAPI | Valencia | 8 | 12 |
| TOTAL | 90 | 89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Rodríguez, C.S.; Marín, Ó.; Calatayud, F.; Mahiques, M.J.; Mompó, A.; Segura, I.; Simó, E.; González-Cabrera, J. Large-Scale Monitoring of Resistance to Coumaphos, Amitraz, and Pyrethroids in Varroa destructor. Insects 2021, 12, 27. https://doi.org/10.3390/insects12010027
Hernández-Rodríguez CS, Marín Ó, Calatayud F, Mahiques MJ, Mompó A, Segura I, Simó E, González-Cabrera J. Large-Scale Monitoring of Resistance to Coumaphos, Amitraz, and Pyrethroids in Varroa destructor. Insects. 2021; 12(1):27. https://doi.org/10.3390/insects12010027
Chicago/Turabian StyleHernández-Rodríguez, Carmen Sara, Óscar Marín, Fernando Calatayud, María José Mahiques, Ana Mompó, Inmaculada Segura, Enrique Simó, and Joel González-Cabrera. 2021. "Large-Scale Monitoring of Resistance to Coumaphos, Amitraz, and Pyrethroids in Varroa destructor" Insects 12, no. 1: 27. https://doi.org/10.3390/insects12010027
APA StyleHernández-Rodríguez, C. S., Marín, Ó., Calatayud, F., Mahiques, M. J., Mompó, A., Segura, I., Simó, E., & González-Cabrera, J. (2021). Large-Scale Monitoring of Resistance to Coumaphos, Amitraz, and Pyrethroids in Varroa destructor. Insects, 12(1), 27. https://doi.org/10.3390/insects12010027