Kairomone and Camera Trapping New Zealand Flower Thrips, Thrips obscuratus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
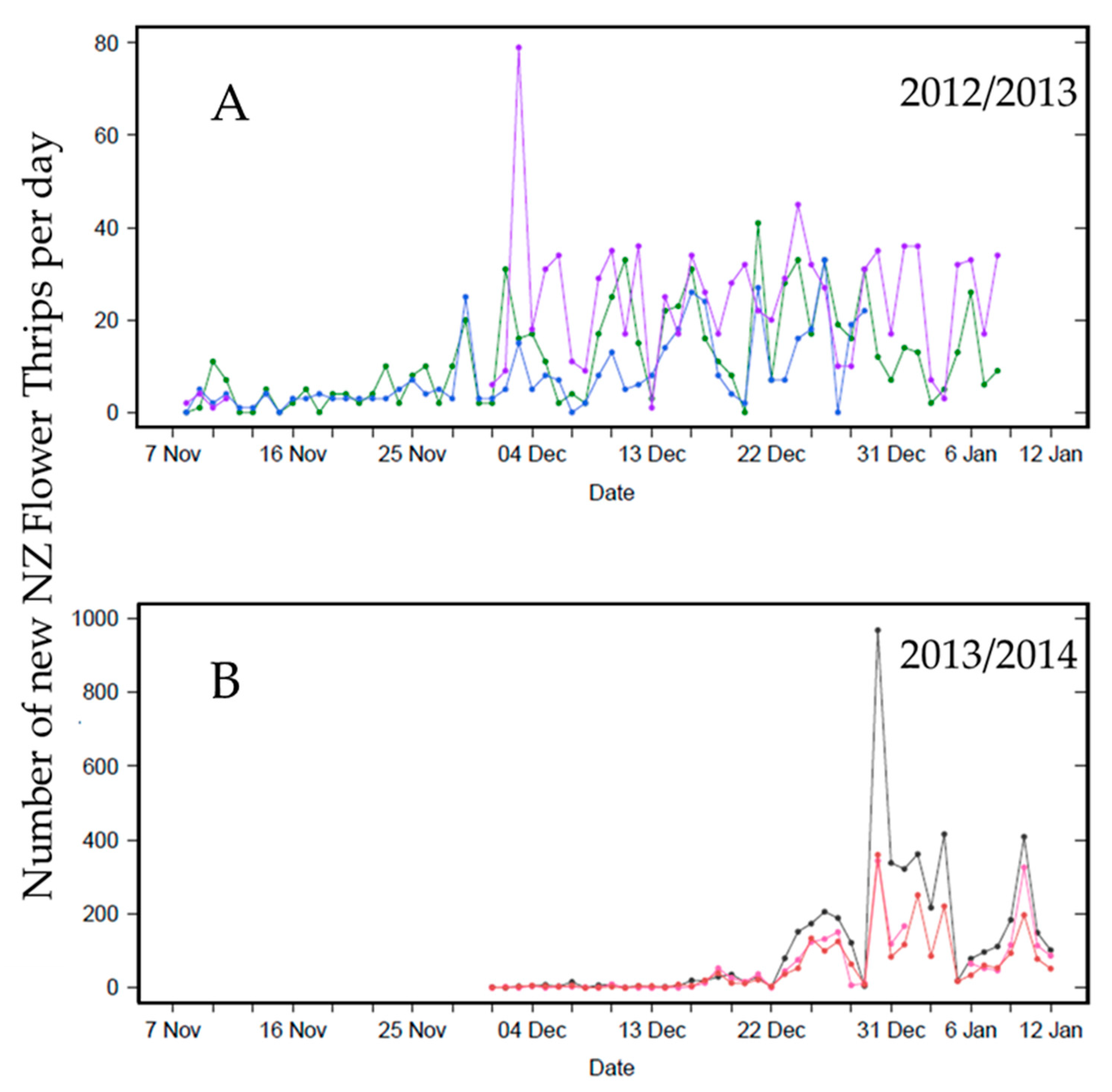
2.1. Daily Thrips Phenology from Self-Reporting Camera Traps
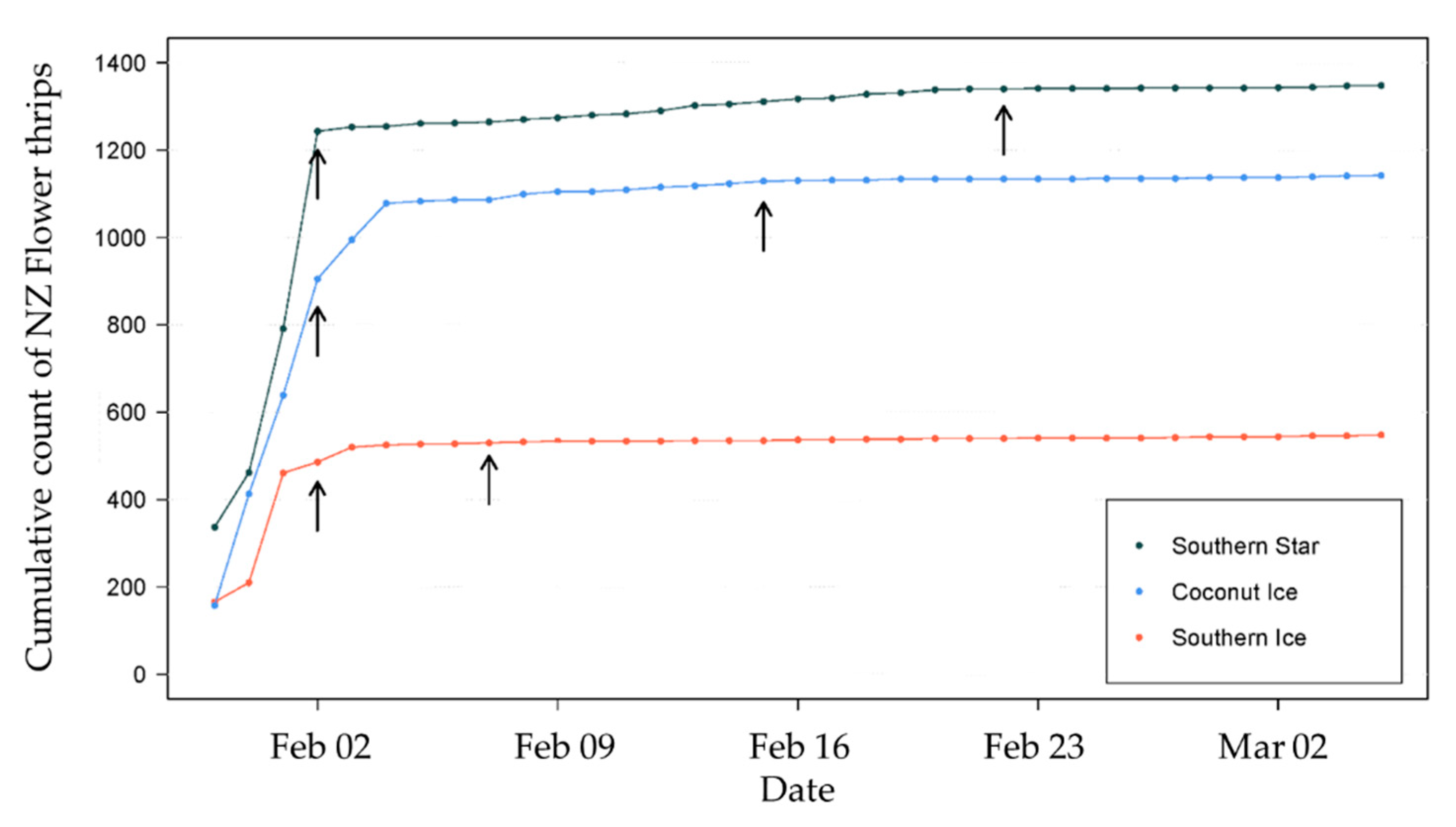
2.2. Effect of Insecticides on Catch
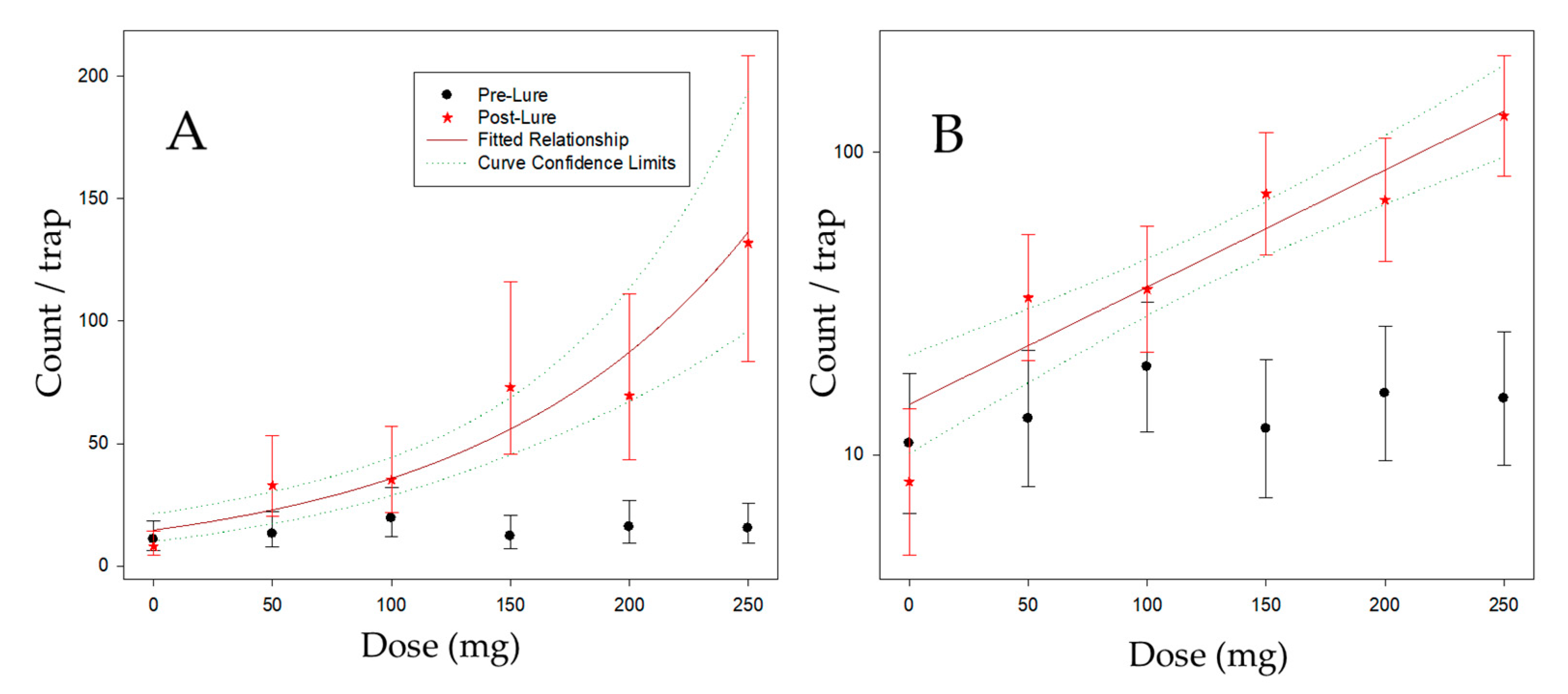
2.3. Dose Response of Thrips to Different 6-PAP Kairomone Loadings in Trees
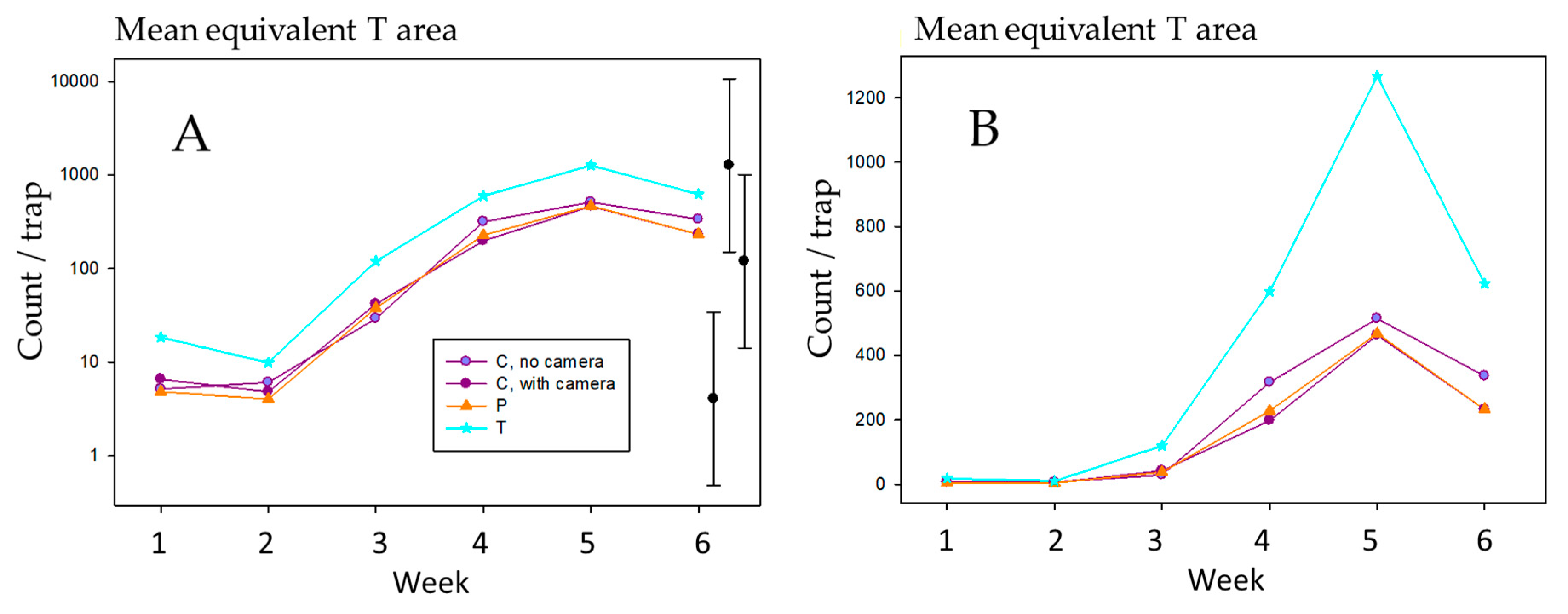
2.4. Effect of Trap Density in One Tree on Catch
3. Results
3.1. Daily Thrips Phenology from Self-Reporting Camera Traps
3.2. Effect of Insecticide Applications on Thrips Catch
3.3. Dose Response of Thrips to Different Kairomone Loadings in Trees
3.4. Effect of Trap Density on Thrips Catch
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wearing, C.H. Evaluating the IPM implementation process. Annu. Rev. Entomol. 1988, 33, 17–38. [Google Scholar] [CrossRef]
- Rothschild, G.H.L.; Minks, A.K. Some factors influencing the performance of pheromone traps for oriental fruit moth in Australia. Entomol. Exp. Appl. 1977, 22, 171–182. [Google Scholar] [CrossRef]
- Acebes-Doria, A.L.; Agnello, A.M.; Alston, D.G.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Blaauw, B.R.; Buntin, G.D.; Burkness, E.C.; et al. Season-long monitoring of the brown marmorated stink bug (Hemiptera: Pentatomidae) throughout the United States using commercially available traps and lures. J. Econ. Entomol. 2019, 113, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Cutler, T.L.; Swann, D.E. Using remote photography in wildlife ecology: A review. Wildl. Soc. Bull. 1999, 27, 571–581. [Google Scholar]
- Lortie, C.; Budden, A.; Reid, A. From birds to bees: Applying video observation techniques to invertebrate pollinators. J. Pollinat. Ecol. 2012, 6, 125–128. [Google Scholar] [CrossRef]
- Reynolds, D.R.; Riley, J. Remote-sensing, telemetric and computer-based technologies for investigating insect movement: A survey of existing and potential techniques. Comput. Electron. Agric. 2002, 35, 271–307. [Google Scholar] [CrossRef]
- Selby, R.D.; Gage, S.H.; Whalon, M.E. Precise and low-cost monitoring of Plum curculio (Coleoptera: Curculionidae) pest activity in pyramid traps with cameras. Environ. Entomol. 2014, 43, 421–431. [Google Scholar] [CrossRef]
- Yao, Q.; Lv, J.; Liu, Q.J.; Diao, G.Q.; Yang, B.J.; Chen, H.M.; Tang, J. An insect imaging system to automate rice light-trap pest identification. J. Integr. Agric. 2012, 11, 978–985. [Google Scholar] [CrossRef]
- Guarnieri, A.; Maini, S.; Molari, G.; Rondelli, V. Automatic trap for moth detection in integrated pest management. Bull. Insect 2011, 64, 247–251. [Google Scholar]
- Shimoda, N.; Kataoka, T.; Okamoto, H.; Terawaki, M.; Hata, S.I. Automatic pest counting system using image processing technique. J. Jap. Soc. Agric. Mach. 2006, 68, 59–64. [Google Scholar]
- Sipos, K. The possibility of automation of sex pheromone trapping: Tested on Resseliella theobaldi (barnes) (Dip., Cecidomyiidae). Afr. J. Agric. Res. 2012, 7, 1410–1413. [Google Scholar] [CrossRef]
- El-Sayed, A.M. The Pherobase: Database of Pheromones and Semiochemicals. Available online: www.pherobase.com (accessed on 30 July 2020).
- McLaren, G.; Alspach, P. The incidence of New Zealand flower thrips in stonefruit orchards between flowering and harvest. N. Z. Plant. Prot. 2006, 59, 63–68. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Mitchell, V.J.; Suckling, D.M.; El-Sayed, A.M. 6-Pentyl-2H-pyran-2-one: A potent peach-derived kairomone for New Zealand flower thrips, Thrips obscuratus. J. Chem. Ecol. 2014, 40, 50–55. [Google Scholar] [CrossRef]
- El-Sayed, A.M.; Suckling, D.M.; Wearing, C.H.; Byers, J.A. Potential of mass trapping for long-term pest management and eradication of invasive species. J. Econ. Entomol. 2006, 99, 1550–1564. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J. A new Isoneurothrips from New Zealand (Thysanoptera, Thripidae). Proc. Entomol. Soc. Wash. 1941, 43, 63–64. [Google Scholar]
- Teulon, D.A.J.; Penman, D.R. Host records for the New Zealand flower thrips (Thrips obscuratus (Crawford) Thysanoptera: Thripidae). N. Z. Entomol. 1990, 13, 46–51. [Google Scholar] [CrossRef]
- McLaren, G.; Fraser, J. Alternative strategies to control New Zealand flower thrips on nectarines. N. Z. Plant. Prot. 2001, 54, 10–14. [Google Scholar] [CrossRef]
- Chhagan, A.; Jamieson, L.E.; Griffin, M.J.; Page-Weir, N.E.M.; Poulton, J.; Zulhendri, F.; Feng, R.; Connolly, P.G.; Davis, V.A.; Olsson, S.; et al. Postharvest management of New Zealand flower thrips (Thrips obscuratus) on apricots using ethyl formate or pyrethrum-based treatments. N. Z. Plant Prot. 2013, 66, 63–74. [Google Scholar]
- Marroni, V.; Teulon, D.A.J.; Scott, R.R.; Jaspers, M.V. Abundance of Thrips obscuratus (Crawford) (Thysanoptera: Thripidae) during wine grape flowering in different regions in New Zealand. N. Z. Entomol. 2012, 35, 10–13. [Google Scholar] [CrossRef]
- McLaren, G.; Fraser, J. Development of thresholds for insecticidal control of New Zealand flower thrips on nectarines in spring. N. Z. Plant. Prot. 2000, 53, 194–199. [Google Scholar] [CrossRef]
- Allen, W.J.; Mitchell, V.J.; Colhoun, K.; Attfield, B.A.; Stanbury, M.E.; Suckling, D.M.; El-Sayed, A.M. Development of an efficient trapping system for New Zealand flower thrips, Thrips obscuratus. Pest. Manag. Sci. 2015, 71, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Penman, D.R.; Osborne, G.O.; Worner, S.P.; Chapman, R.B.; McLaren, G.F. Ethyl nicotinate: A chemical attractant for Thrips obscuratus (Thysanoptera: Thripidae) in stonefruit in New Zealand. J. Chem. Ecol. 1982, 8, 1299–1303. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Stringer, L.D.; Kean, J.M.; Lo, P.L.; Bell, V.; Walker, J.T.S.; Twidle, A.M.; Jiménez-Pérez, A.; El-Sayed, A.M. Spatial analysis of mass trapping: How close is close enough? Pest Manage. Sci. 2015, 71, 1452–1461. [Google Scholar] [CrossRef]
- Clare, G.; Suckling, D.M.; Bradley, S.J.; Walker, J.T.S.; Shaw, P.W.; Daly, J.M.; McLaren, G.F.; Wearing, C.H. Pheromone trap colour determines catch of non-target insects. N. Z. Plant Prot. 2000, 53, 216–220. [Google Scholar]
- Rassati, D.; Faccoli, M.; Chinellato, F.; Hardwick, S.; Suckling, D.M.; Battisti, A. Web-based automatic traps for early detection of alien wood-boring beetles. Entomol. Exp. Appl. 2016, 160, 91–95. [Google Scholar] [CrossRef]
- Lennon, Z.; Stanbury, M.; Suckling, D.; El-Sayed, A. Making data count. N. Z. Plant. Prot. 2015, 68, 450. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman & Hall: London, UK, 1989; p. 511. [Google Scholar]
- Lee, Y.; Nelder, J.A.; Pawitan, Y. Generalized Linear Models with Random Effects: Unified Analysis via H-likelihood; Chapman & Hall/CRC Press: London, UK, 2006; Volume 106, p. 416. [Google Scholar]
- GenStat Committee. The Guide to the GenStat® Command Language (Release 17); VSN International: Hemel Hempsted, UK, 2014. [Google Scholar]
- El-Sayed, A.M.; Mitchell, V.J.; McLaren, G.F.; Manning, L.A.; Bunn, B.; Suckling, D.M.; El-Sayed, A.M. Attraction of New Zealand flower thrips, Thrips obscuratus, to cis-jasmone, a volatile identified from japanese honeysuckle flowers. J. Chem. Ecol. 2009, 35, 656–663. [Google Scholar] [CrossRef]
- Davidson, J.; Andrewartha, H.G. Annual trends in a natural population of Thrips imaginis (thysanoptera). J. Anim. Ecol. 1948, 17, 193–199. [Google Scholar] [CrossRef]
- Martin, N.A.; Workman, P.J.; Hedderley, D.; Fagan, L.L. Monitoring onion (Allium cepa) crops for onion thrips (Thrips tabaci) (Thysanoptera: Thripidae): Testing a commercial protocol. N. Z. J. Crop. Hortic. Sci. 2008, 36, 145–152. [Google Scholar] [CrossRef]
- Suckling, D.M.; McLaren, G.F.; Manning, L.-A.; Mitchell, V.J.; Attfield, B.; Colhoun, K.; El-Sayed, A.M. Development of single-dispenser pheromone suppression of Epiphyas postvittana, Planotortrix octo and Ctenopseustis obliquana in New Zealand stone fruit orchards. Pest Manag. Sci. 2012, 68, 928–934. [Google Scholar] [CrossRef]
- Walker, J.T.; Suckling, D.M.; Wearing, C.H. Past, present, and future of integrated control of apple pests: The New Zealand experience. Annu. Rev. Entomol. 2017, 62, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; El-Sayed, A.M.; Walker, J.T. Regulatory innovation, mating disruption and 4-playtm in New Zealand. J. Chem. Ecol. 2016, 42, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Stringer, L.D.; Stephens, A.E.A.; Woods, B.; Williams, D.G.; Baker, G.; El-Sayed, A.M. From integrated pest management to integrated pest eradication: Technologies and future needs. Pest Manag. Sci. 2014, 70, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Broughton, S.; Cousins, D.A.; Rahman, T. Evaluation of semiochemicals for their potential application in mass trapping of Frankliniella occidentalis (Pergande) in roses. Crop Prot. 2015, 67, 130–135. [Google Scholar] [CrossRef]
- Abdullah, Z.S.; Greenfield, B.P.J.; Ficken, K.J.; Taylor, J.; Wood, M.; Butt, T. A new attractant for monitoring western flower thrips, Frankliniella occidentalis in protected crops. SpringerPlus 2015, 4, 89. [Google Scholar] [CrossRef]
- Natwick, E.T.; Byers, J.A.; Chu, C.C.; Lopez, M.; Henneberry, T.J. Early detection and mass trapping of Frankliniella occidentalis1, and Thrips tabaci (Lin) vegetable Crops. Southwest. Entomol. 2007, 32, 229–238. [Google Scholar] [CrossRef]
- Sampson, C.; Kirk, W.D.J. Can mass trapping reduce thrips damage and is it economically viable? Management of the western flower thrips in strawberry. PLoS ONE 2013, 8, 80787. [Google Scholar] [CrossRef]
- Teulon, D.A.J.; Davidson, M.M.; Perry, N.B.; Nielsen, M.C.; Castañé, C.; Bosch, D.; Riudavets, J.; van Tol, R.W.H.M.; de Kogel, W.J. Methyl isonicotinate—A non-pheromone thrips semiochemical—And its potential for pest management. Int. J. Trop. Insect Sci. 2017, 37, 50–56. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suckling, D.M.; Stanbury, M.E.; Lennon, O.; Colhoun, K.M.; Chinellato, F.; El-Sayed, A.M. Kairomone and Camera Trapping New Zealand Flower Thrips, Thrips obscuratus. Insects 2020, 11, 622. https://doi.org/10.3390/insects11090622
Suckling DM, Stanbury ME, Lennon O, Colhoun KM, Chinellato F, El-Sayed AM. Kairomone and Camera Trapping New Zealand Flower Thrips, Thrips obscuratus. Insects. 2020; 11(9):622. https://doi.org/10.3390/insects11090622
Chicago/Turabian StyleSuckling, David Maxwell, Mailee E. Stanbury, Ox Lennon, Kate M. Colhoun, Fabio Chinellato, and Ashraf M. El-Sayed. 2020. "Kairomone and Camera Trapping New Zealand Flower Thrips, Thrips obscuratus" Insects 11, no. 9: 622. https://doi.org/10.3390/insects11090622
APA StyleSuckling, D. M., Stanbury, M. E., Lennon, O., Colhoun, K. M., Chinellato, F., & El-Sayed, A. M. (2020). Kairomone and Camera Trapping New Zealand Flower Thrips, Thrips obscuratus. Insects, 11(9), 622. https://doi.org/10.3390/insects11090622