Resilin Distribution and Sexual Dimorphism in the Midge Antenna and Their Influence on Frequency Sensitivity

 , ,
, ,  and
and 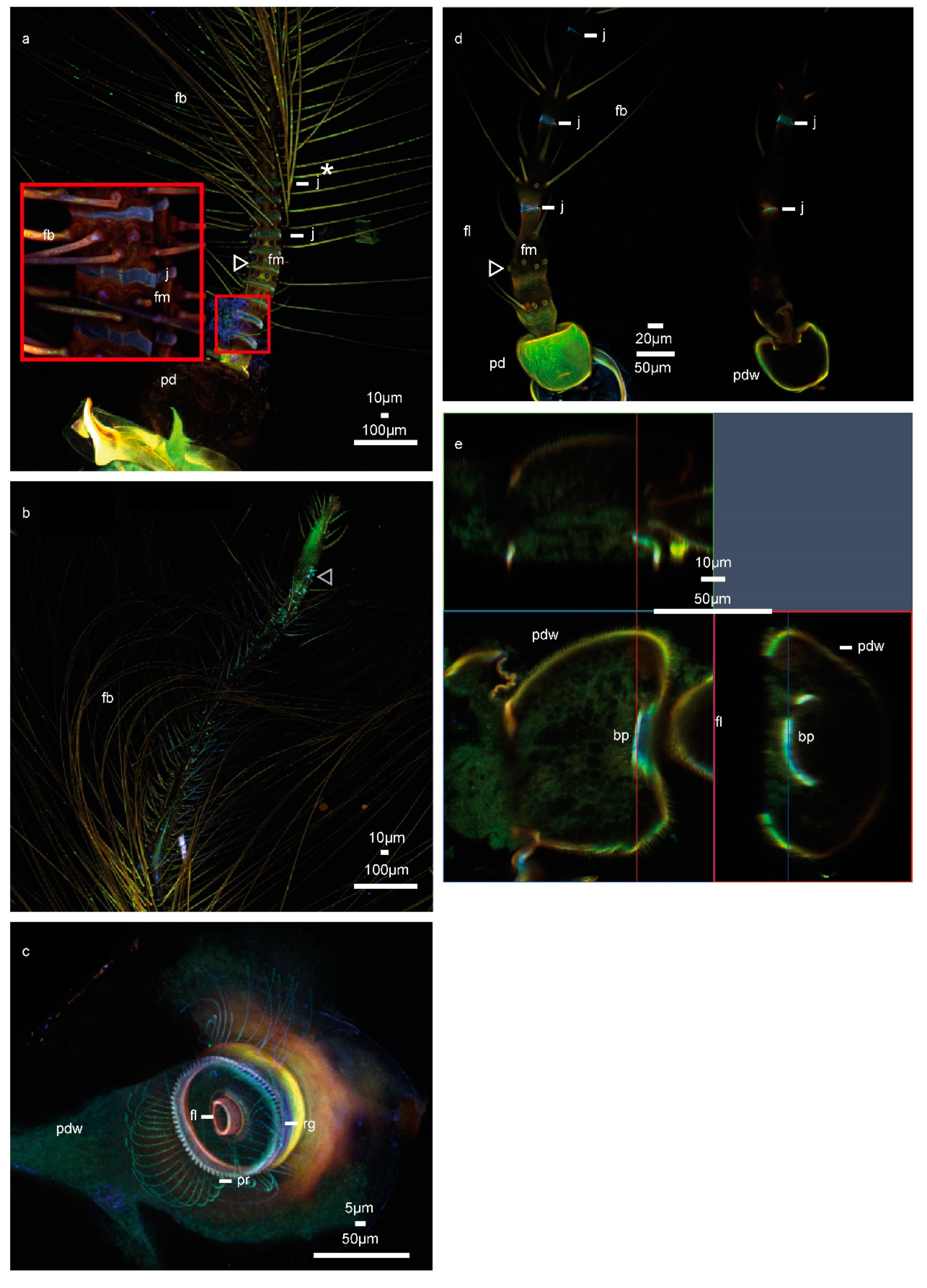{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Specimen Preparation
2.2. Confocal Laser Scanning Microscopy (CLSM)
2.3. Finite Element Modelling (FEM)
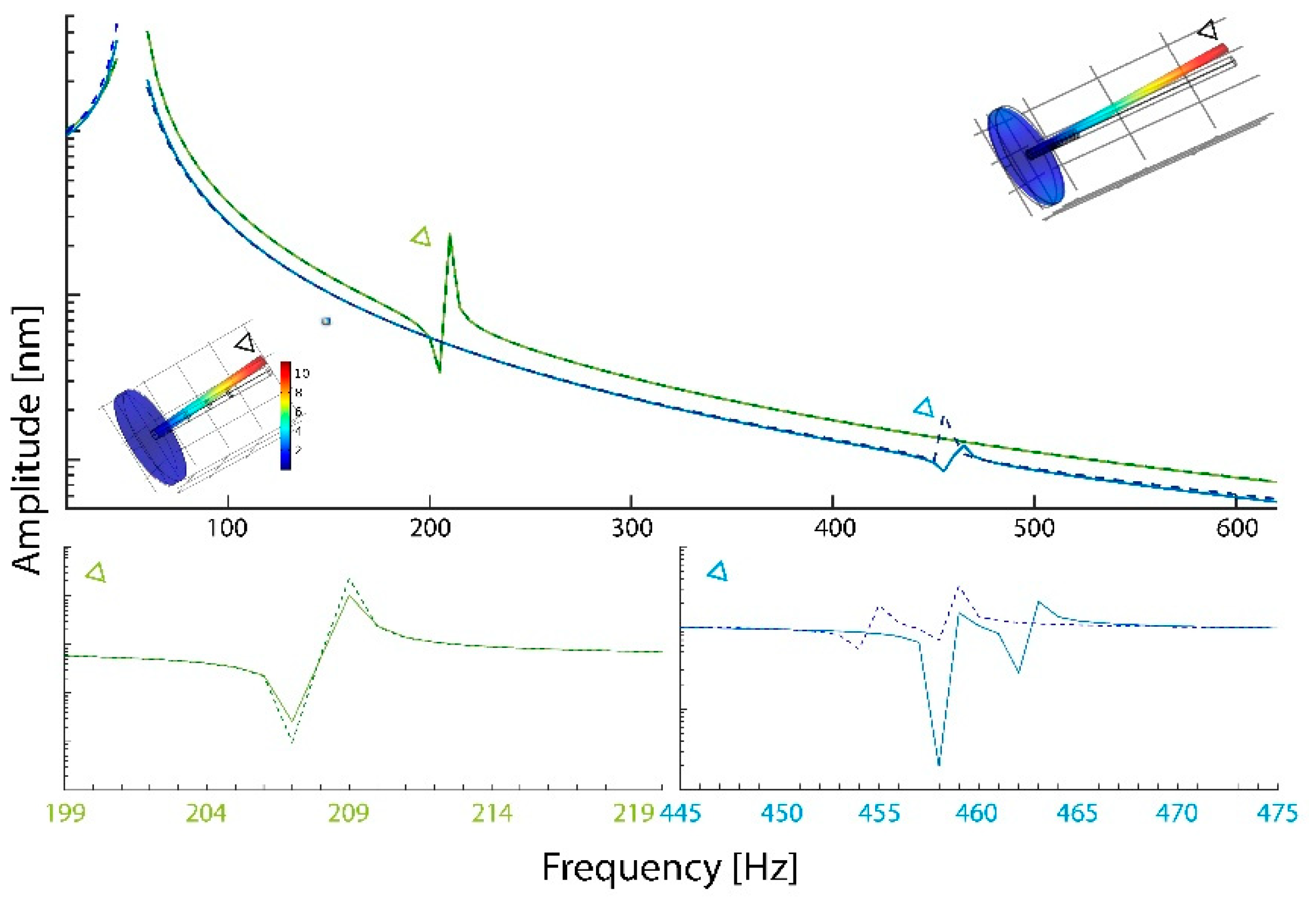
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The Chironomidae: Biology and Ecology of Non-Biting Midges, 1st ed.; Armitage, P.D., Pinder, L.C., Cranston, P.S., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Dévai, G. Ecological background and importance of the change of chironomid fauna (Diptera: Chironomidae) in shallow Lake Balaton. In Developments in Hydrobiology; Dumont, H.J., Ed.; Springer: Heidelberg, Germany, 1990; Volume 53, pp. 189–198. [Google Scholar]
- Berg, M.B.; Hellenthal, R.A. The role of Chironomidae in energy flow of a lotic ecosystem. Neth. J. Aquat. Ecol. 1992, 26, 471–476. [Google Scholar] [CrossRef]
- Huryn, A.D.; Wallace, J.B. A method for obtaining in situ growth rates of larval Chironomidae (Diptera) and its application to studies of secondary production 1. Limnol. Oceanogr. 1986, 31, 216–221. [Google Scholar] [CrossRef]
- Downes, J.A. The swarming and mating flight of diptera. Annu. Rev. Entomol. 1969, 14, 271–298. [Google Scholar] [CrossRef]
- Caspary, V.G.; Downe, A.E.R. Swarming and mating of Chironomus riparius (Diptera: Chironomidae). Can. Entomol. 1971, 103, 444–448. [Google Scholar] [CrossRef]
- Belton, P. An analysis of direction finding in male mosquitoes. In Experimental Analysis of Insect Behavior; Springer: Berlin/Heidelberg, Germany, 1974; pp. 139–148. [Google Scholar]
- Göpfert, M.C.; Briegel, H.; Robert, D. Mosquito hearing: Sound-induced antennal vibrations in male and female Aedes aegypti. J. Exp. Biol. 1990, 202, 2727–2738. [Google Scholar]
- Cator, L.J.; Arthur, B.J.; Harrington, L.C.; Hoy, R.R. Harmonic convergence in the love songs of the dengue vector mosquito. Science 2009, 323, 1077–1079. [Google Scholar] [CrossRef]
- Kamikouchi, A.; Inagaki, H.K.; Effertz, T.; Hendrich, O.; Fiala, A.; Göpfert, M.C.; Ito, K. The neural basis of Drosophila gravity-sensing and hearing. Nature 2009, 458, 165–171. [Google Scholar] [CrossRef]
- Matsuo, E.; Kamikouchi, A. Neuronal encoding of sound, gravity, and wind in the fruit fly. J. Comp. Physiol. A 2013, 199, 253–262. [Google Scholar] [CrossRef]
- Johnston, C. Original communications: Auditory apparatus of the Culex mosquito. Q. J. Microsc. Sci. 1855, 1, 97–102. [Google Scholar]
- Boo, K.S.; Richards, A.G. Fine structure of the scolopidia in the Johnston’s organ of male Aedes aegypti (L.) (Diptera: Culicidae). Int. J. Insect Morphol. Embryol. 1975, 4, 549–566. [Google Scholar] [CrossRef]
- Hart, M.; Belton, P.; Kuhn, R. The Risler Manuscript. Eur. Mosq. Bull. 2011, 29, 103–113. [Google Scholar]
- Sane, S.P.; McHenry, M.J. The biomechanics of sensory organs. Integr. Comp. Biol. 2009, 49, i8–i23. [Google Scholar] [CrossRef]
- Saltin, B.D.; Matsumura, Y.; Reid, A.; Windmill, J.F.; Gorb, S.N.; Jackson, J.C. Material stiffness variation in mosquito antennae. J. R. Soc. Interface 2009, 16, 20190049. [Google Scholar] [CrossRef] [PubMed]
- Michels, J.; Gorb, S.N. Detailed three-dimensional visualization of resilin in the exoskeleton of arthropods using confocal laser scanning microscopy. J. Microsc. 2012, 245, 1–16. [Google Scholar] [CrossRef]
- Peisker, H.; Michels, J.; Gorb, S.N. Evidence for a material gradient in the adhesive tarsal setae of the ladybird beetle Coccinella septempunctata. Nat. Commun. 2013, 4, 1661. [Google Scholar] [CrossRef]
- Willkommen, J.; Michels, J.; Gorb, S.N. Functional morphology of the male caudal appendages of the Damselfly Ischnura elegans (Zygoptera: Coenagrionidae). Arthropod Struct. Dev. 2015, 44, 289–300. [Google Scholar] [CrossRef]
- Filippov, A.E.; Matsumura, Y.; Kovalev, A.E.; Gorb, S.N. Stiffness gradient of the beetle penis facilitates propulsion in the spiraled female spermathecal duct. Sci. Rep. 2016, 6, 27608. [Google Scholar] [CrossRef]
- Michels, J.; Appel, E.; Gorb, S.N. Functional diversity of resilin in Arthropoda. Beilstein J. Nanotechnol. 2016, 7, 1241–1259. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, M.; Büscher, T.H.; Gorb, S.N.; Rajabi, H. How does a slender tibia resist buckling? Effect of material, structural and geometric characteristics on buckling behaviour of the hindleg tibia in stick insect postembryonic development. J. Exp. Biol. 2018, 221, 1–4. [Google Scholar] [CrossRef]
- Bergmann, P.; Richter, S.; Glöckner, N.; Betz, O. Morphology of hindwing veins in the shield bug Graphosoma italicum (Heteroptera: Pentatomidae). Arthropod Struct. Dev. 2018, 47, 375–390. [Google Scholar] [CrossRef]
- Eshghi, S.H.; Jafarpour, M.; Darvizeh, A.; Gorb, S.N.; Rajabi, H. A simple, high-resolution, non-destructive method for determining the spatial gradient of the elastic modulus of insect cuticle. J. R. Soc. Interface 2018, 15, 20180312. [Google Scholar] [CrossRef] [PubMed]
- Neff, D.; Frazier, S.F.; Quimby, L.; Wang, R.-T.; Zill, S. Identification of resilin in the leg of cockroach, Periplaneta americana: Confirmation by a simple method using pH dependence of UV fluorescence. Arthropod Struct. Dev. 2000, 29, 75–83. [Google Scholar] [CrossRef]
- Matsumura, Y.; Kovalev, A.E.; Gorb, S.N. Penetration mechanics of a beetle intromittent organ with bending stiffness gradient and a soft tip. Sci. Adv. 2017, 3, 5469–5477. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.F.; Wegst, U.G. Design and mechanical properties of insect cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef]
- Wegst, U.G.K.; Ashby, M.F. The mechanical efficiency of natural materials. Philos. Mag. 2004, 84, 2167–2181. [Google Scholar] [CrossRef]
- Kovalev, A.; Filippov, A.; Gorb, S.N. Slow viscoelastic response of resilin. J. Comp. Physiol. A 2018, 204, 409–417. [Google Scholar] [CrossRef]
- Rajabi, H.; Shafiei, A.; Darvizeh, A.; Gorb, S.N.; Duerr, V.; Dirks, J.-H. Both stiff and compliant: Morphological and biomechanical adaptations of stick insect antennae for tactile exploration. J. R. Soc. Interface 2018, 15, 20180246. [Google Scholar] [CrossRef]
- Avitabile, D.; Homer, M.; Champneys, A.R.; Jackson, J.C.; Robert, D. Mathematical modelling of the active hearing process in mosquitoes. J. R. Soc. Interface 2010, 7, 105–122. [Google Scholar] [CrossRef]
- Darwin, C. The Origin of Species; The Temple Press: Letchworth, UK, 1928; reprinted 1934. [Google Scholar]
- West-Eberhard, M.J. Sexual selection, competitive communication and species specific signals in insects. In Insect Communication, Proceedings of the 12th Symposium of the Royal Entomological Society of London, London, UK, 7–9 September 1983; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Laland, K.; Uller, T.; Feldman, M.; Sterelny, K.; Müller, G.B.; Moczek, A.; Jablonka, E.; Odling-Smee, J.; Wray, G.A.; Hoekstra, H.E.; et al. Does evolutionary theory need a rethink? Nature News 2014, 514, 161. [Google Scholar] [CrossRef]
- Gerhardt, R.R.; Hribar, L.J. Flies (Diptera). In Medical and Veterinary Entomology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 171–190. [Google Scholar]
- Loudon, C. Flexural stiffness of insect antennae. Am. Entomol. 2005, 51, 48–49. [Google Scholar] [CrossRef]
- Dickinson, M.H.; Lehmann, F.-O.; Sanjay, P.; Sane, S.P. Wing rotation and the aerodynamic basis of insect flight. Science 1999, 284, 1954–1960. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Bendy bugs inspire roboticists. Science 2016, 351, 647. [Google Scholar] [CrossRef] [PubMed]
- White, Z.W.; Vernerey, F.J. Armours for soft bodies: How far can bioinspiration take us? Bioinspiration Biomim. 2018, 13, 041004. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Ueno, N.; Tsuji, T. Active Antenna-basic considerations on the working principle. In Proceedings of the IEEE/RSJ International Conference on Intelligent Robots and Systems (IROS’94), Munich, Germany, 12–16 September 1994; Volume 3, pp. 1744–1750. [Google Scholar] [CrossRef]
- Cowan, N.J.; Ma, E.J.; Cutkosky, M.; Full, R.J. A biologically inspired passive antenna for steering control of a running robot. In Robotics Research. The Eleventh International Symposium; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Barth, F.G.; Németh, S.S.; Friedrich, O.C. Arthropod touch reception: Structure and mechanics of the basal part of a spider tactile hair. J. Comp. Physiol. A 2004, 190, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Schaber, C.F.; Gorb, S.N.; Barth, F.G. Force transformation in spider strain sensors: White light interferometry. J. R. Soc. Interface 2012, 9, 1254–1264. [Google Scholar] [CrossRef] [PubMed]
- Schaber, C.F.; Barth, F.G. Spider joint hair sensilla: Adaptation to proprioreceptive stimulation. J. Comp. Physiol. A 2015, 201, 235–248. [Google Scholar] [CrossRef]
- Sutton, G.P.; Clarke, D.; Morley, E.L.; Robert, D. Mechanosensory hairs in bumblebees (Bombus terrestris) detect weak electric fields. Proc. Natl. Acad. Sci. USA 2016, 113, 7261–7265. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saltin, B.D.; Matsumura, Y.; Reid, A.; Windmill, J.F.; Gorb, S.N.; Jackson, J.C. Resilin Distribution and Sexual Dimorphism in the Midge Antenna and Their Influence on Frequency Sensitivity. Insects 2020, 11, 520. https://doi.org/10.3390/insects11080520
Saltin BD, Matsumura Y, Reid A, Windmill JF, Gorb SN, Jackson JC. Resilin Distribution and Sexual Dimorphism in the Midge Antenna and Their Influence on Frequency Sensitivity. Insects. 2020; 11(8):520. https://doi.org/10.3390/insects11080520
Chicago/Turabian StyleSaltin, Brian D., Yoko Matsumura, Andrew Reid, James F. Windmill, Stanislav N. Gorb, and Joseph C. Jackson. 2020. "Resilin Distribution and Sexual Dimorphism in the Midge Antenna and Their Influence on Frequency Sensitivity" Insects 11, no. 8: 520. https://doi.org/10.3390/insects11080520
APA StyleSaltin, B. D., Matsumura, Y., Reid, A., Windmill, J. F., Gorb, S. N., & Jackson, J. C. (2020). Resilin Distribution and Sexual Dimorphism in the Midge Antenna and Their Influence on Frequency Sensitivity. Insects, 11(8), 520. https://doi.org/10.3390/insects11080520