Ultramorphological Characteristics of Falsogastrallus sauteri Pic (Coleoptera: Ptinidae) and a New Species of Cephalonomia Westwood (Hymenoptera: Bethylidae): A Book-Boring Beetle and Its Natural Enemy in Taiwan



Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collecting
2.2. Specimen Examination and Deposition
2.3. Terminology
- Total length (TL): total length of outstretched individual, from the clypeus to the abdominal apex.
- Total width (TW): maximum width of outstretched individual.
- Head length (HL): maximum length of head excluding mandibles in full-face view.
- Head width (HW): maximum width of head including eyes in full-face view.
- Width of frons (WF): minimum width between eyes in full-face view.
- Eye length (EL): maximum length or height of eye in lateral view.
- Diameter of anterior ocellus (DAO): maximum diameter of anterior ocellus.
- Width of ocellar triangle (WOT): distance across and including posterior ocelli.
- Posterior ocellar-line (POL): shortest distance between posterior ocelli.
- Antero-posterior ocellar-line (AOL): shortest distance between anterior ocellus and posterior ocellus.
- Ocello-ocular line (OOL): shortest distance from a posterior ocellus to nearest eye margin.
- Length of mesosoma (LM): maximum diagonal length of mesosoma excluding the pronotal collar, in lateral view.
- Length of propodeal disc (LPD): measured along the midline excluding posterior declivity, in dorsal view.
- Width of propodeal disc (WPD): maximum width of the disc excluding the anterior potion from the level of propodeal spiracles, in dorsal view.
- Forewing length (FWL): maximum length of forewing.
- Total length (TL): total length of outstretched individual, from the mandibular apex to the metasomal apex.
3. Results
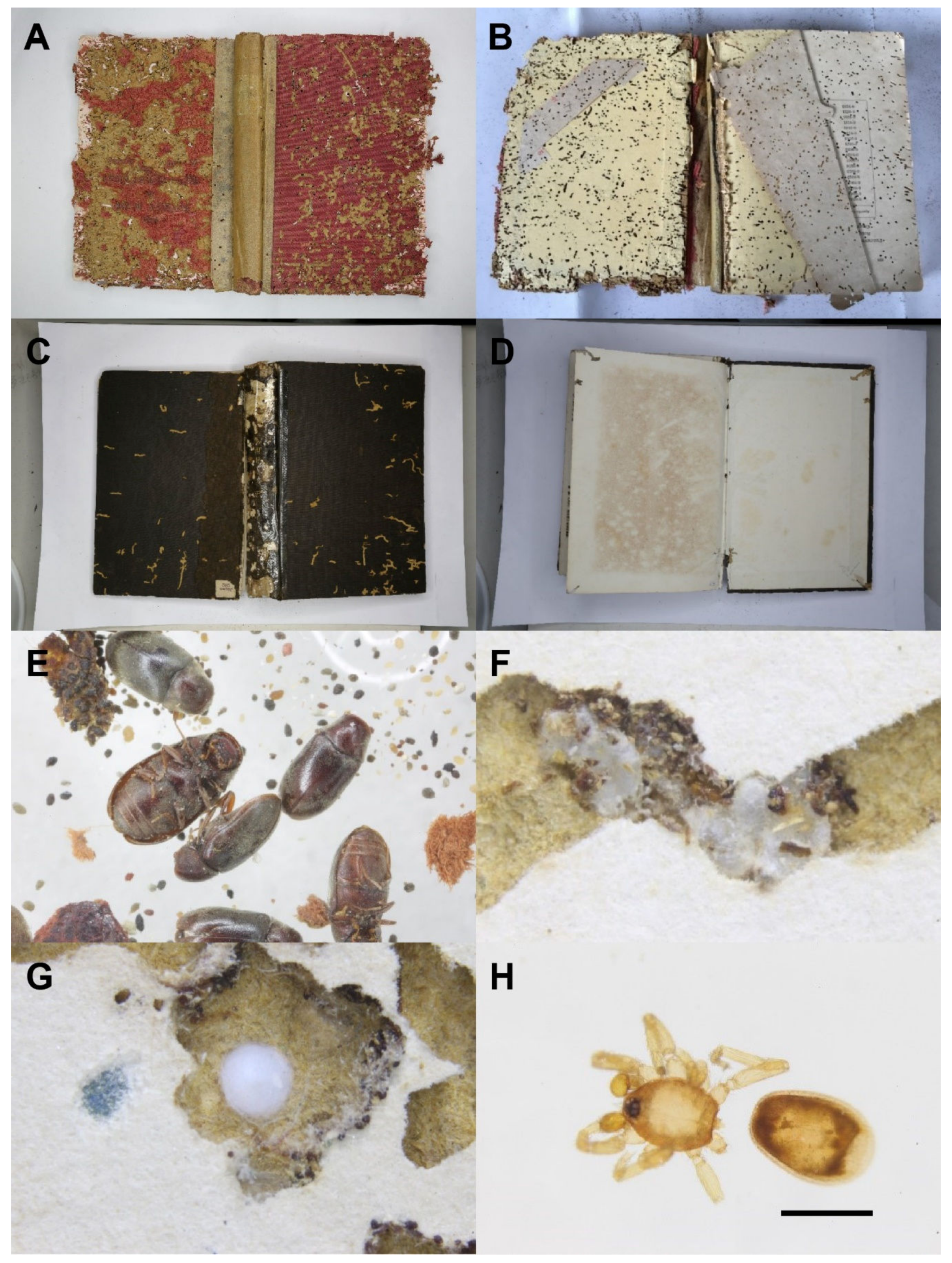
3.1. Examination of Books Damaged by Falsogastrallus sauteri and the Frass from the Books
3.2. Taxonomy
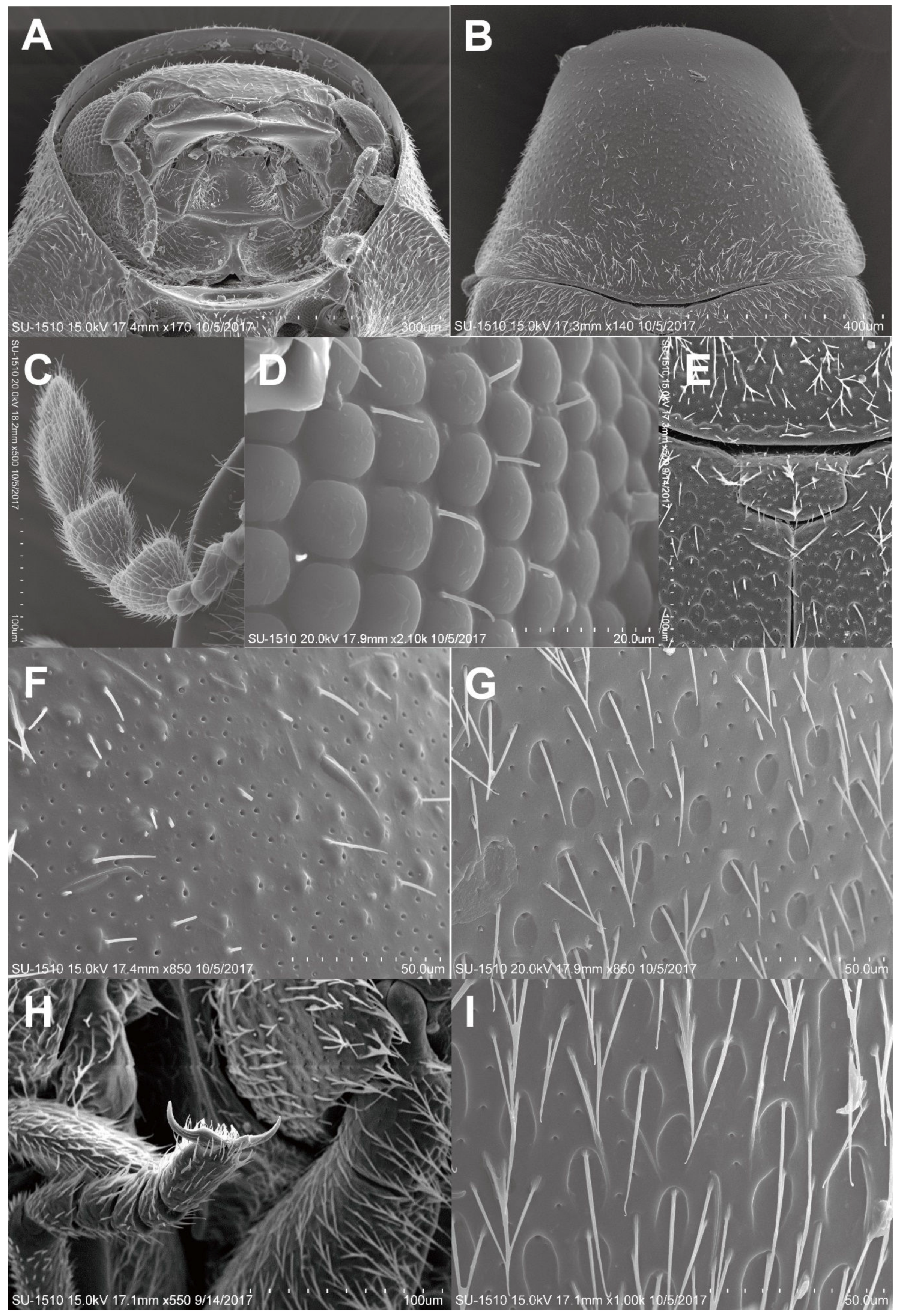
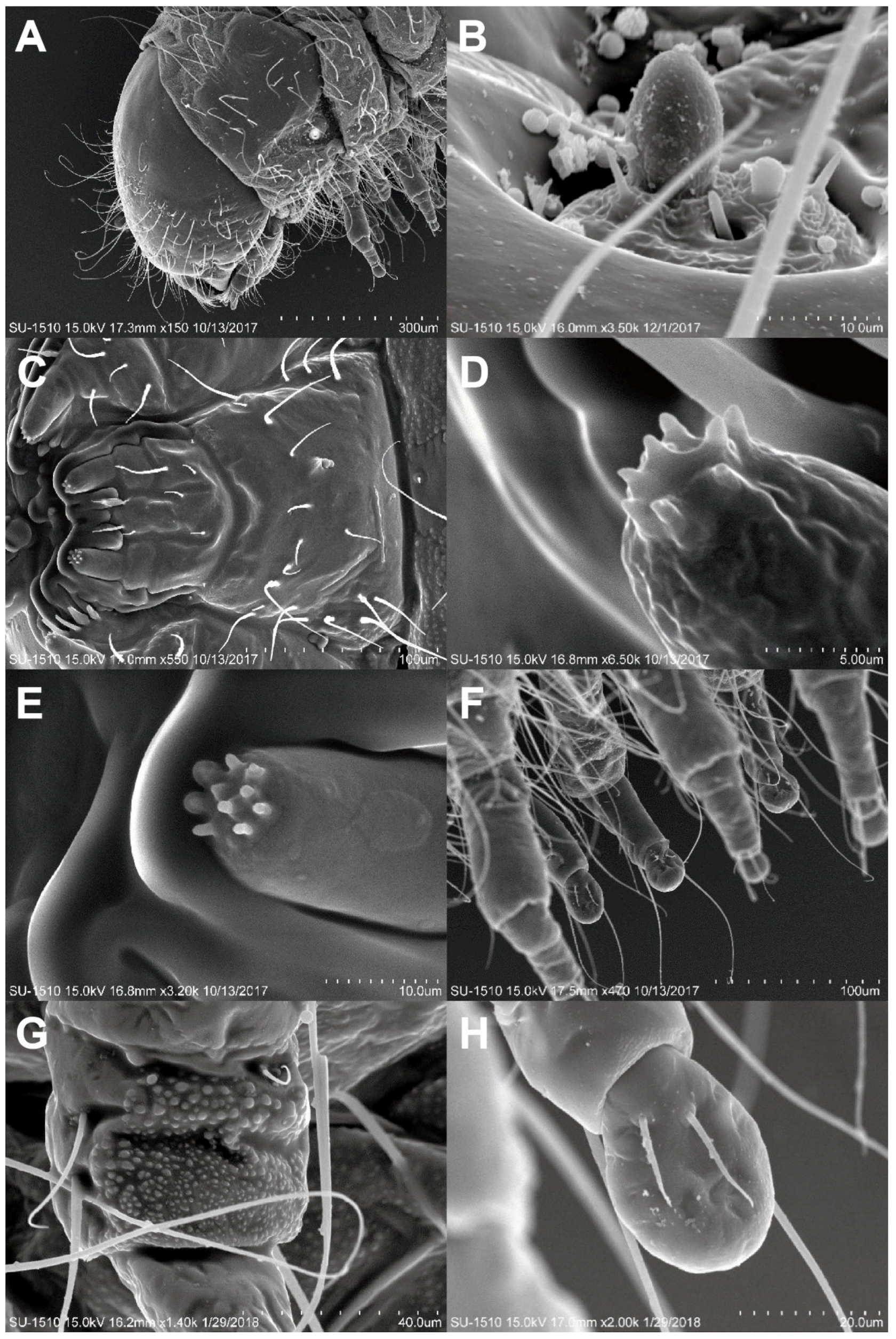
3.2.1. Falsogastrallus sauteri Pic, 1914
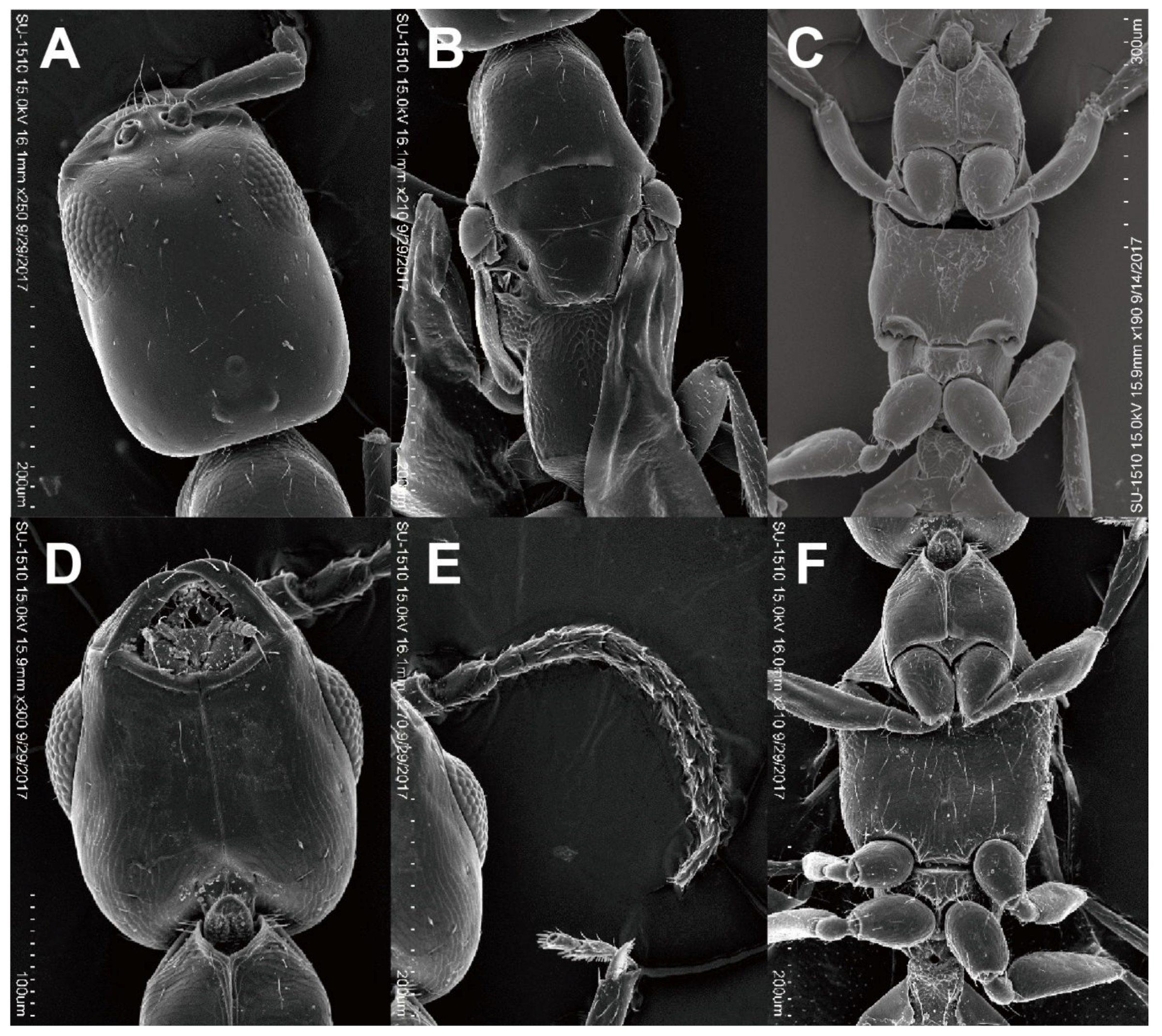
3.2.2. Cephalonomia formosiensis Terayama & Ho sp. nov.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pinniger, D. Managing Pests in Paper-Based Collections; Preservation Advisory Centre, British Library: London, UK, 2012; p. 15. [Google Scholar]
- Pic, M. Anobiidae. Coleoptm. Cat. 1912, 10, 1–92. [Google Scholar]
- Borowski, J.; Zahradníc, P. Ptinidae. In Catalogue of Palaearctic Coleoptera; Lobl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; pp. 328–362. [Google Scholar]
- Bell, K.; Philips, T. Molecular systematics and evolution of the Ptinidae (Coleoptera: Bostrichoidea) and related families. Zool. J. Linn. Soc. 2012, 165, 88–108. [Google Scholar] [CrossRef][Green Version]
- Komine, Y.; Hayashi, M.; Kigawa, R.; Harada, M.; Miura, S.; Kawanobe, W.; Ishizaki, T. Species and characteristics of wood-boring anobiids caught by sticky insect traps in historic buildings in Nikko world heritage site. Sci. Conserv. 2011, 50, 133–140. (In Japanese) [Google Scholar]
- Manachini, B. Alien insect impact on cultural heritage and landscape: An underestimated problem. Conserv. Sci. Cult. Herit. 2015, 15, 61. [Google Scholar]
- Moşneagu, M. The preservation of cultural heritage damaged by anobiids (Insecta, Coleoptera, Anobiidae). Ann. Ser. Biol. Sci. Acad. Rom. Sci. 2012, 1, 32–65. [Google Scholar]
- Querner, P. Insect Pests and Integrated Pest Management in Museums, Libraries and Historic Buildings. Insects 2015, 6, 595–607. [Google Scholar] [CrossRef]
- Trematerra, P.; Pinniger, D. Museum Pests—Cultural Heritage Pests. In Recent Advances in Stored Product Protection; Springer: Berlin/Heidelberg, Germany, 2018; pp. 229–260. [Google Scholar]
- Silva, C.R.; Anjos, N.D.; Zanuncio, J.C.; Serrão, J.E. Damage to books caused by Tricorynus herbarius (Gorham) (Coleoptera: Anobiidae). Coleopt. Bull. 2013, 67, 175–178. [Google Scholar] [CrossRef]
- Watson, J.R. A tropical book worm in Florida. Neogastrallus librinocens Fisher, in Florida (Anobiidae, Col.). Fla. Entomol. 1943, 26, 61–63. [Google Scholar] [CrossRef]
- White, R.E. The Mexican book beetle, Catorama herbarium, established in the United States (Coleoptera: Anobiidae). Ann. Ent. Soc. Am. 1963, 56, 280–285. [Google Scholar] [CrossRef]
- Hong-Wang, H.H. Study on Pest Control of Chinese Rare Books in Taiwan; Hanmei tushu Ltd.: Taipei, Taiwan, 1991; 173p. [Google Scholar]
- Sakai, M. Mimogastrallus hideoi, a new genus and a new species of the subfamily Anobiinae, with notes on the related genus Falsogastrallus (Coleoptera, Anobiidae). Jpn. J. Syst. Entomol. 2003, 9, 333–338. [Google Scholar]
- Yen, S.F. Pests control and prevention in libraries. Inf. Manag. Buddh. Libr. 2006, 43, 40–49. [Google Scholar]
- Zahradník, P. Contribution to knowledge of the tribe Gastrallini (Coleoptera: Bostrichoidea: Ptinidae)—IV. Review of the genus Falsogastrallus, with description of new species. Stud. Rep. Taxon. Ser. 2010, 6, 271–275. [Google Scholar]
- Xiong, X.Z.; Feng, H.F.; Xiong, H.Z.; Sun, K. Studies on the morphology of Falsogastrallus sauteri Pic. J. Southwest Agric. Univ. 1990, 12, 508–512. (In Chinese) [Google Scholar]
- Azevedo, C.O.; Alencar, I.D.C.C.; Ramos, M.S.; Barbosa, D.N.; Colombo, W.D.; Vargas, J.M.R.; Lim, J. Global guide of the flat wasps (Hymenoptera, Bethylidae). Zootaxa 2018, 4489, 1–294. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E. A synopsis of the American Bethylidae (Hymenoptera, Aculeata). Bull. Mus. Comp. Zool. 1964, 132, 1–222. [Google Scholar]
- Gordh, G.; Móczár, L. A catalog of the world Bethylidae (Hymenoptera, Aculeata). Mem. Am. Ent. Inst. 1990, 46, 1–364. [Google Scholar]
- Lim, J.; Lee, S. Pararhabdepyris gorbatobskii (Hymenoptera: Bethylidae), new to Korea and the first host record of Allobethylus Kieffer. J. Asia Pac. Biodivers. 2014, 7, 457–461. [Google Scholar] [CrossRef]
- Terayama, M. Additions to knowledge of the bethylid wasp fauna of Japan (Hymenoptera; Bethylidae): Part 1. J. Hymenopterol. Soc. Jpn. 2014, 25, 1–10. [Google Scholar]
- Terayama, M. Check list of Bethylidae of the Oriental and the southeastern part of Palaearctic regions (Insecta: Hymenoptera). Bull. Toho Gakuen. 1993, 8, 1–32. [Google Scholar]
- Terayama, M. Bethylidae (Hymenoptera). Insects Jpn. 2006, 1, 1–319. [Google Scholar]
- Xu, Z.; Weng, L.; He, J. A new species of Cephalonomia Westwood (Hymenoptera: Bethylidae) parasitizing Rhizopertha dominica (F.) (Coleoptera: Bostrychidae). Proc. Ent. Soc. Washi. 2008, 110, 477–480. [Google Scholar] [CrossRef]
- Terayama, M. Descriptions of new taxa and distribution records of the family Bethylidae (Insecta, Hymenoptera). III. Subfamily Epyrinae. Bull. Kanto Gakuen Univ. 2005, 13, 51–112. [Google Scholar]
- Richards, O.W. The British Bethylidae (s.l.) (Hymenoptera). Trans. R. Ent. Soc. Lond. 1939, 89, 299–305. [Google Scholar] [CrossRef]
- Emden, F.V. Zur Kenntnis der morphologie und Oekologie der Brotkäfer-Parasiten Cephalonomia quadridentata Duchoussoy. Z. Morph. Oekol. Yiere. 1931, 23, 425–574. [Google Scholar] [CrossRef]
- Kearns, C.M. Method of wing inheritance in Cephalonomia gallicola Ashmead (Bethylidae: Hymenoptera). Ann. Ent. Soc. Am. 1934, 27, 533–541. [Google Scholar] [CrossRef]
- Moraal, L.; de Rond, J. Nieuwe vindplaatsen van de zwamplakopwesp Caphalonomia formiciformis (Hymenoptera: Bethylidae). Ent. Berichten. 2007, 4, 150–151. [Google Scholar]
- Evans, H.E. A new species of Cephalonomia exhibiting an unusually complex polymorphism (Hymenoptera, Bethylidae). Psyche 1963, 70, 151–163. [Google Scholar] [CrossRef][Green Version]
- Evans, H.E. The Bethylidae of America north of Mexico. Mem. Am. Ent. Inst. 1978, 27, 1–332. [Google Scholar]
- Philips, T.K.; Bell, K.L. 6.4. Ptinidae Latreille. In Handbuch der Zoologie/Handbook of Zoology; Beutel, R.G., Leschen, R.A.B., Eds.; Insecta: Coleoptera, Beetles; Walter de Gruyter: Berlin, Germany, 2010; Volume 2, pp. 217–225. [Google Scholar]
- Pic, M. Ptinidae et Anobiidae (Col.). Suppl. Ent. 1914, 3, 8–11. [Google Scholar]
- Xiong, X.Z. Laboratory rearing method and life history of Falsogastrallus sauteri Pic (Coleoptera: Anobiidae). Coleopt. Bull. 1992, 46, 321–324. [Google Scholar]
- Xiong, H.Z.; Sun, K.; Feng, H.F.; Xiong, X.Z. Study on occurrence of Falsogastrallus sauteri Pic. Arch. Sci. Study 1991, 5, 73–78. (In Chinese) [Google Scholar]
- Abraham, Y.J.; Moore, D.; Godwin, G. Rearing and aspects of biology of Cephalonomia stephanoderis and Prorops nasuta (Hymenoptera: Bethylidae) parasitoids of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entomol. Res. 1990, 80, 121–128. [Google Scholar] [CrossRef]
- Howard, R.W. Comparative analysis of cuticular hydrocarbons from ectoparasitoids Cephalonomia waterstoni and Laelius utilis (Hymenoptera: Bethylidae) and their respective hosts, Cryptolestes ferrugineus (Coleoptera: Cucujidae) and Trogoderma variable (Coleoptera: Dermestidae). Ann. Entomol. Soc. Am. 1992, 80, 317–325. [Google Scholar]
- Howard, R.W.; Pérez-Lachaud, G. Cuticular hydrocarbons of the ectoparasitic wasp Cephalonomia hyalinipennis (Hymenoptera: Bethylidae) and its alternative host, the stored product pest Caulophilus oryzae (Coleoptera: Curculionidae). Arch. Insect Biochem. 2002, 50, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Oh, M.; Lee, J.; Lee, S. Cephalonomia gallicola (Hymenoptera: Bethylidae), New to Korea, an ectoparasitoid of the cigarette beetle, Lasioderma serricorne (Coleoptera: Anobiidae). J. Asia Pac. Entomol. 2007, 10, 335–338. [Google Scholar] [CrossRef]
- Muesebeck, C.F.W.; Walkley, L.M. Family Bethylidae. In Hymenoptera of America North of Mexico. Synoptic Catalog; Muesebeck, C.F.W., Krombein, K.V., Townes, H.K., Eds.; U.S. Department of Agriculture: Washington, DC, USA, 1951; Volume 2, pp. 726–734. [Google Scholar]
- Oda, T.; Mori, A.; Fujita, K.; Monkada, L.; Tachikawa, T.; Tanaka, S. A case of the sting of a parasitic wasp, Sclerodermus sp. (Hymenoptera: Bethylidae). Trop. Med. 1981, 23, 213–216. [Google Scholar]
- Pérez-Lachaud, G.; Hardy, I.C.W. Alternative hosts for bethylid parasitoids of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Biol. Control 2001, 22, 265–277. [Google Scholar] [CrossRef]
- Rilett, R.O. The biology of Cephalonomia waterstoni Gahan. Can. J. Res. 1949, 27, 93–111. [Google Scholar] [CrossRef]
- Zeiri, A.; Braham, M. First record of Cephalonomia hypobori on Scolytus amygdali in Tunisia. Tunis. J. Plant Prot. 2011, 6, 43–47. [Google Scholar]
- Kimberly, A.E.; Emley, A.L. A Study of the Deterioration of Book Papers in Libraries; U.S. Bur. Standards Misc. Pub. 140: Washington, DC, USA, 1933; pp. 1–7. [Google Scholar]
- Harris, A.C. The effects on a human of the sting of Goniozus antipodum (Hymenoptera: Bethylidae). N. Z. Entomol. 1996, 19, 49. [Google Scholar] [CrossRef]
- Papini, P.A. A case of stings in humans caused by Sclerodermus sp. in Italy. J. Venom Anim. Toxins Incl. Trop. Dis. 2014, 20, 11. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Publishing Country Year | UK | Japan | USA | Germany | The Netherlands | Australia | India | Taiwan | Sum |
|---|---|---|---|---|---|---|---|---|---|
| 1862 | 1 | 1 | |||||||
| 1891 | 1 | 1 | |||||||
| 1892 | 1 | 1 | |||||||
| 1893 | 1 | 1 | |||||||
| 1897 | 3 | 3 | |||||||
| 1900 | 1 | 1 | |||||||
| 1901 | 1 | 1 | 1 | 3 | |||||
| 1902 | 2 | 1 | 3 | ||||||
| 1904 | 1 | 1 | |||||||
| 1905 | 1 | 1 | |||||||
| 1906 | 1 | 1 | 2 | ||||||
| 1907 | 1 | 1 | |||||||
| 1908 | 1 | 1 | |||||||
| 1909 | 1 | 1 | 2 | ||||||
| 1910 | 1 | 1 | |||||||
| 1911 | 1 | 1 | 2 | ||||||
| 1913 | 3 | 1 | 4 | ||||||
| 1914 | 1 | 1 | |||||||
| 1916 | 1 | 1 | 1 | 3 | |||||
| 1919 | 1 | 1 | |||||||
| 1920 | 1 | 1 | |||||||
| 1922 | 2 | 2 | |||||||
| 1923 | 1 | 1 | |||||||
| 1926 | 1 | 1 | |||||||
| 1931 | 1 | 1 | |||||||
| 1934 | 1 | 2 | 3 | ||||||
| 1943 | 2 | 2 | |||||||
| 1974 | 1 | 1 | |||||||
| Sum | 19 | 10 | 6 | 4 | 4 | 1 | 1 | 1 | 46 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, Y.-H.; Hsiao, Y.; Terayama, M.; Chan, M.-L. Ultramorphological Characteristics of Falsogastrallus sauteri Pic (Coleoptera: Ptinidae) and a New Species of Cephalonomia Westwood (Hymenoptera: Bethylidae): A Book-Boring Beetle and Its Natural Enemy in Taiwan. Insects 2020, 11, 223. https://doi.org/10.3390/insects11040223
Ho Y-H, Hsiao Y, Terayama M, Chan M-L. Ultramorphological Characteristics of Falsogastrallus sauteri Pic (Coleoptera: Ptinidae) and a New Species of Cephalonomia Westwood (Hymenoptera: Bethylidae): A Book-Boring Beetle and Its Natural Enemy in Taiwan. Insects. 2020; 11(4):223. https://doi.org/10.3390/insects11040223
Chicago/Turabian StyleHo, Yu-Hsiang, Yun Hsiao, Mamoru Terayama, and Mei-Ling Chan. 2020. "Ultramorphological Characteristics of Falsogastrallus sauteri Pic (Coleoptera: Ptinidae) and a New Species of Cephalonomia Westwood (Hymenoptera: Bethylidae): A Book-Boring Beetle and Its Natural Enemy in Taiwan" Insects 11, no. 4: 223. https://doi.org/10.3390/insects11040223
APA StyleHo, Y.-H., Hsiao, Y., Terayama, M., & Chan, M.-L. (2020). Ultramorphological Characteristics of Falsogastrallus sauteri Pic (Coleoptera: Ptinidae) and a New Species of Cephalonomia Westwood (Hymenoptera: Bethylidae): A Book-Boring Beetle and Its Natural Enemy in Taiwan. Insects, 11(4), 223. https://doi.org/10.3390/insects11040223