Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Insects
2.3. Generation of pUAST.CYP392A16
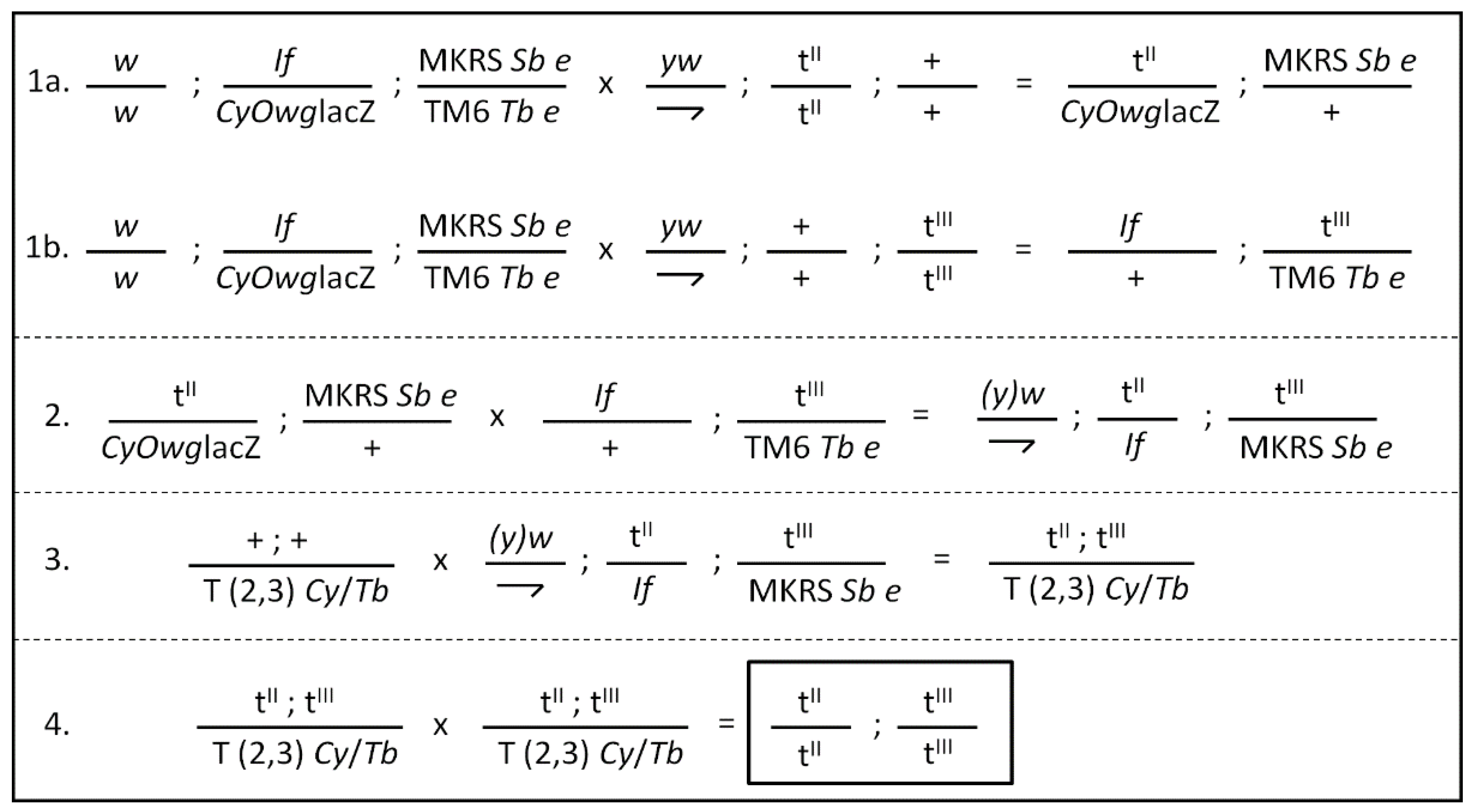
2.4. Construction of the Transgenic Fly Strains
2.5. Expression of CYP392A16 and/or TuCPR in D. Melanogaster
2.6. Extraction of RNA, cDNA Synthesis and Reverse Transcription PCR
2.7. Toxicity Bioassays
3. Results
3.1. Generation of Transgenic Lines Bearing UAS-CYP392A16 and UAS-TuCPR
3.2. GAL4/UAS Transgenic Co-Expression of CYP392A16 and TuCPR Confers Resistance to Abamectin
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Migeon, A.; Nouguier, E.; Dorkeld, F. Spider mites web: A comprehensive database for the Tetranychidae. In Trends in Acarology; Sabelis, M., Bruin, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 557–560. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Dermauw, W.; Grbić, M.; Tirry, L.; Feyereisen, R. Spider mite control and resistance management: Does a genome help? Pest Manag. Sci. 2013, 69, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Tirry, L.; Yamamoto, A.; Nauen, R.; Dermauw, W. The economic importance of acaricides in the control of phytophagous mites and an update on recent acaricide mode of action research. Pestic. Biochem. Physiol. 2015, 121, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Tirry, L. Mechanisms of acaricide resistance in the two-spotted spider mite Tetranychus urticae. In Biorational Control of Arthropod Pests; Ishaaya, I., Horowitz, A.R., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 347–393. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Dermauw, W. The molecular evolution of xenobiotic metabolism and resistance in chelicerate mites. Annu. Rev. Entomol. 2016, 61, 475–498. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vanholme, B.; Van Pottelberge, S.; Van Nieuwenhuyse, P.; Nauen, R.; Tirry, L.; Denholm, I. Mitochondrial heteroplasmy and the evolution of insecticide resistance: Non-Mendelian inheritance in action. Proc. Nat. Acad. Sci. USA 2008, 105, 5980–5985. [Google Scholar] [CrossRef] [PubMed]
- Tsagkarakou, A.; Van Leeuwen, T.; Khajehali, J.; Ilias, A.; Grispou, M.; Williamson, M.S.; Tirry, L.; Vontas, J. Identifications of pyrethroid resistance associated mutations in the para sodium channel of the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). Insect Mol. Biol. 2009, 18, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Khajehali, J.; Van Leeuwen, T.; Grispoy, M.; Morou, E.; Alout, H.; Weill, M.; Tirry, L.; Vontas, J.; Tsagkarakou, A. Acetylcholinesterase point mutations in European strains of Tetranychus urticae (Acari: Tetranychidae) resistant to organophosphates. Pest Manag. Sci. 2010, 66, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.H.; Yoon, K.S.; Clark, J.M.; Lee, S.H. A point mutation in a glutamate-gated chloride channel confers abamectin resistance in the two-spotted spider mite, Tetranychus urticae Koch. Insect Mol. Biol. 2010, 19, 583–591. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Demaeght, P.; Osborne, E.J.; Dermauw, W.; Gohlke, S.; Nauen, R.; Grbić, M.; Tirry, L.; Merzendorfer, H.; Clark, R.M. Population bulk segregant mapping uncovers resistance mutations and the mode of action of a chitin synthesis inhibitor in arthropods. Proc. Nat. Acad. Sci. USA 2012, 109, 4407–4412. [Google Scholar] [CrossRef]
- Dermauw, W.; Ilias, A.; Riga, M.; Tsagkarakou, A.; Grbić, M.; Tirry, L.; Van Leeuwen, T.; Vontas, J. The cys-loop ligand-gated ion channel gene family of Tetranychus urticae: Implications for acaricide toxicology and a novel mutation associated with abamectin resistance. Insect Biochem. Mol. Biol. 2012, 42, 455–465. [Google Scholar] [CrossRef]
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Bajda, S.; Dermauw, W.; Panteleri, R.; Sugimoto, N.; Douris, V.; Tirry, L.; Osakabe, M.; Vontas, J.; Van Leeuwen, T. A mutation in the PSST homologue of complex I (NADH: Ubiquinone oxidoreductase) from Tetranychus urticae is associated with resistance to METI acaricides. Insect Biochem. Mol. Biol. 2017, 80, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Riga, M.; Bajda, S.; Themistokleous, C.; Papadaki, S.; Palzewicz, M.; Dermauw, W.; Vontas, J.; Van Leeuwen, T. The relative contribution of target-site mutations in complex acaricide resistant phenotypes as assessed by marker assisted backcrossing in Tetranychus urticae. Sci. Rep. 2017, 7, 9202. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Adenasaya, A.W.; Morales, M.A.; Walsh, D.B.; Lavine, L.C.; Lavine, M.D.; Zhu, F. Multiple acaricide resistance and underlying mechanisms in Tetranychus urticae on hops. J. Pest Sci. 2019, 92, 543–555. [Google Scholar] [CrossRef]
- Fotoukkiaii, S.M.; Tan, Z.; Xue, W.; Wybouw, N.; Van Leeuwen, T. Identification and characterization of new mutations in mitochondrial cytochrome b that confer resistance to bifenazate and acequinocyl in the spider mite Tetranychus urticae. Pest Manag. Sci. 2020, 76, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Snoeck, S.; Njiru, C.; Inak, E.; Dermauw, W.; Van Leeuwen, T. Geographical distribution and molecular insights into abamectin and milbemectin cross-resistance in European field populations of Tetranychus urticae. Pest Manag. Sci. 2020, 76, 2569–2581. [Google Scholar] [CrossRef]
- Sugimoto, N.; Takahashi, A.; Ihara, R.; Itoh, Y.; Jouraku, A.; Van Leeuwen, T.; Osakabe, M. QTL mapping using microsatellite linkage reveals target-site mutations associated with high levels of resistance against three mitochondrial complex II inhibitors in Tetranychus urticae. Insect Biochem. Mol. Biol. 2020, 123, 103410. [Google Scholar] [CrossRef]
- Papapostolou, K.M.; Riga, M.; Charamis, J.; Skoufa, E.; Souchlas, V.; Ilias, A.; Dermauw, D.; Ioannidis, P.; Van Leeuwen, T.; Vontas, J. Identification and characterization of striking multiple-insecticide resistance in a Tetranychus urticae field population from Greece. Pest Manag. Sci. 2020, in press. [Google Scholar] [CrossRef]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbić, M.; Clark, R.M.; Feyereisen, R.; Van Leeuwen, T. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Nat. Acad. Sci. USA 2013, 110, E113–E122. [Google Scholar] [CrossRef]
- Demaeght, P.; Dermauw, W.; Tsakireli, D.; Khajehali, J.; Nauen, R.; Tirry, L.; Vontas, J.; Lümmen, P.; Van Leeuwen, T. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae: CYP392E10 metabolizes spirodiclofen, but not its corresponding enol. Insect Biochem. Mol. Biol. 2013, 43, 544–554. [Google Scholar] [CrossRef]
- Riga, M.; Tsakireli, D.; Ilias, A.; Morou, E.; Myridakis, A.; Stephanou, E.G.; Nauen, R.; Dermauw, W.; Van Leeuwen, T.; Paine, M.; et al. Abamectin is metabolized by CYP392A16, a cytochrome P450 associated with high levels of acaricide resistance in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 46, 43–53. [Google Scholar] [CrossRef]
- Riga, M.; Myridakis, A.; Tsakireli, D.; Morou, E.; Stephanou, E.G.; Nauen, R.; Van Leeuwen, T.; Douris, V.; Vontas, J. Functional characterization of the Tetranychus urticae CYP392A11, a cytochrome P450 that hydroxylates the METI acaricides cyenopyrafen and fenpyroximate. Insect Biochem. Mol. Biol. 2015, 65, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Fotoukkiaii, S.M.; Mermans, C.; Wybouw, N.; Van Leeuwen, T. Resistance risk assessment of the novel complex II inhibitor pyflubumide in the polyphagous pest Tetranychus urticae. J. Pest Sci. 2020, 93, 1085–1096. [Google Scholar] [CrossRef]
- Pavlidi, N.; Mousaalreza, K.; Myridakis, A.; Dermauw, W.; Wybouw, N.; Tsakireli, D.; Stephanou, E.G.; Labrou, N.E.; Vontas, J.; Van Leeuwen, T. A glutathione-S-transferase (TuGSTd05) associated with acaricide resistance in Tetranychus urticae directly metabolizes the complex II inhibitor cyflumetofen. Insect Biochem. Mol. Biol. 2017, 80, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Koo, H.N.; Kim, S.I.; Park, B.; Kim, H.; Kim, G.H. Target-site mutations and glutathione S-transferases are associated with acequinocyl and pyridaben resistance in the two-spotted spider mite Tetranychus urticae (Acari: Tetranychidae). Insects 2020, 11, 511. [Google Scholar] [CrossRef]
- Wei, P.; Demaeght, P.; De Schutter, K.; Grigoraki, L.; Labropoulou, V.; Riga, M.; Vontas, J.; Nauen, R.; Dermauw, W.; Van Leeuwen, T. Overexpression of an alternative allele of carboxyl/choline esterase 4 (CCE04) of Tetranychus urticae is associated with high levels of resistance to the keto-enol acaricide spirodiclofen. Pest Manag. Sci. 2020, 76, 1142–1153. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T. The ABC gene family in arthropods: Comparative genomics and role in insecticide transport and resistance. Insect Biochem. Mol. Biol. 2014, 45, 89–110. [Google Scholar] [CrossRef]
- Snoeck, S.; Pavlidi, N.; Pipini, D.; Vontas, J.; Dermauw, W.; Van Leeuwen, T. Substrate specificity and promiscuity of horizontally transferred UDP-glucosyltransferases in the generalist herbivore Tetranychus urticae. Insect Biochem. Mol. Biol. 2019, 109, 116–127. [Google Scholar] [CrossRef]
- Clark, J.M.; Scott, J.G.; Campos, F.; Bloomquist, J.R. Resistance to avermectins: Extent, mechanisms, and management implications. Annu. Rev. Entomol. 1994, 40, 1–30. [Google Scholar] [CrossRef]
- Wolstenholme, A.J. Glutamate-gated chloride channels. J. Biol. Chem. 2012, 287, 40232–40238. [Google Scholar] [CrossRef]
- Bajda, S.; Riga, M.; Wybouw, N.; Papadaki, S.; Ouranou, E.; Fotoukkiaii, S.M.; Vontas, J.; Van Leeuwen, T. Fitness costs of key point mutations that underlie acaricide target-site resistance in the two-spotted spider mite Tetranychus urticae. Evol. Appl. 2018, 11, 1540–1553. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y.; Zhang, Y.; Wu, Q.; Guo, Z.; Xie, W.; Zhou, X.; Wang, S. Transcriptome profiling and functional analysis suggest that the constitutive overexpression of four cytochrome P450s confers resistance to abamectin in Tetranychus urticae from China. Pest Manag. Sci. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP genes and P450 Enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Academic Press: London, UK, 2012; pp. 236–316. [Google Scholar]
- Grbić, M.; Van Leeuwen, T.; Clark, R.M.; Rombauts, S.; Rouzé, P.; Grbić, V.; Osborne, E.J.; Dermauw, W.; Ngoc, P.C.T.; Ortego, F.; et al. The genome of Tetranychus urticae reveals herbivorous pest adaptations. Nature 2011, 479, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, S.; Kurlovs, A.H.; Bajda, S.; Feyereisen, R.; Greenhalgh, R.; Villacis-Perez, E.; Kosterlitz, O.; Dermauw, W.; Clark, R.M.; Van Leeuwen, T. High-resolution QTL mapping in Tetranychus urticae reveals acaricide-specific responses and common target-site resistance after selection by different METI-I acaricides. Insect Biochem. Mol. Biol. 2019, 110, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Wybouw, N.; Kosterlitz, O.; Kurlovs, A.H.; Bajda, S.; Greenhalgh, R.; Snoeck, S.; Bui, H.; Bryon, A.; Dermauw, W.; Van Leeuwen, T.; et al. Long-term population studies uncover the genome structure and genetic basis of xenobiotic and host plant adaptation in the herbivore Tetranychus urticae. Genetics 2019, 211, 1409–1427. [Google Scholar] [CrossRef]
- Adesanya, A.W.; Cardenas, A.; Lavine, M.D.; Walsh, D.B.; Lavine, L.C.; Zhu, F. RNA interference of NADPH-cytochrome P450 reductase increases susceptibilities to multiple acaricides in Tetranychus urticae. Pestic. Biochem. Physiol. 2020, 165, 104550. [Google Scholar] [CrossRef]
- Homem, R.A.; Davies, T.G.E. An overview of functional genomic tools in deciphering insect resistance. Curr. Opin. Insect Sci. 2018, 27, 103–110. [Google Scholar] [CrossRef]
- Perry, T.; Batterham, P. Harnessing model organisms to study insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 61–67. [Google Scholar] [CrossRef]
- Scott, J.G.; Buchon, N. Drosophila melanogaster as a powerful tool for studying insect toxicology. Pestic. Biochem. Physiol. 2019, 161, 95–103. [Google Scholar] [CrossRef]
- Douris, V.; Denecke, S.; Van Leeuwen, T.; Nauen, R.; Bass, C.; Vontas, J. Using CRISPR/Cas9 genome modification to understand the genetic basis of insecticide resistance: Drosophila and beyond. Pestic. Biochem. Physiol. 2020, 167, 104595. [Google Scholar] [CrossRef]
- McLeman, A.; Troczka, B.J.; Homem, R.A.; Duarte, A.; Zimmer, C.; Garrood, W.T.; Pym, A.; Beadle, K.; Reid, R.J.; Douris, V.; et al. Fly-Tox: A panel of transgenic flies expressing pest and pollinator cytochrome P450s. Pestic. Biochem. Physiol. 2020, 169, 104674. [Google Scholar] [CrossRef]
- Samantsidis, G.R.; Panteleri, R.; Denecke, S.; Kounadi, S.; Christou, I.; Nauen, R.; Douris, V.; Vontas, J. ‘What I cannot create, I do not understand’: Functionally validated synergism of metabolic and target site insecticide resistance. Proc. R. Soc. B 2020, 287, 20200838. [Google Scholar] [CrossRef] [PubMed]
- Daborn, P.J.; Lumb, C.; Boey, A.; Wong, W.; ffrench-Constant, R.H.; Batterham, P. Evaluating the insecticide resistance potential of eight Drosophila melanogaster cytochrome P450 genes by transgenic over-expression. Insect Biochem. Mol. Biol. 2007, 37, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Bloomington Drosophila Stock Center, Indiana University Bloomington. Available online: https://bdsc.indiana.edu/information/recipes/uclfood.html (accessed on 22 November 2020).
- Pavlidi, N.; Monastirioti, M.; Daborn, P.; Livadaras, I.; Van Leeuwen, T.; Vontas, J. Transgenic expression of the Aedes aegypti CYP9J28 confers pyrethroid resistance in Drosophila melanogaster. Pestic. Biochem. Physiol. 2012, 104, 132–135. [Google Scholar] [CrossRef]
- Chung, H.; Bogwitz, M.R.; McCart, C.; Andrianopoulos, A.; ffrench-Constant, R.H.; Batterham, P.; Daborn, P.J. Cis-regulatory elements in the Accord retrotransposon result in tissue-specific expression of the Drosophila melanogaster insecticide resistance gene Cyp6g1. Genetics 2007, 175, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.L.; Russel, R.M.; Preisler, H.K.; Savin, N.E. Bioassays in Arthropods, 2nd ed.; CRC Press, Taylor and Francis Group LLC: Boca Raton, FL, USA, 2007. [Google Scholar]
- Karunker, I.; Morou, E.; Nikou, D.; Nauen, R.; Sertchook, R.; Stevenson, B.J.; Paine, M.J.I.; Morin, S.; Vontas, J. Structural model and functional characterization of the Bemisia tabaci CYP6CM1vQ, a cytochrome P450 associated with high levels of imidacloprid resistance. Insect Biochem. Mol. Biol. 2009, 39, 697–706. [Google Scholar] [CrossRef]
- Zimmer, C.T.; Garrood, W.T.; Singh, K.S.; Randall, E.; Lueke, B.; Gutbrod, O.; Matthiesen, S.; Kohler, M.; Nauen, R.; Davies, T.G.E. Neofunctionalization of duplicated P450 genes drives the evolution of insecticide resistance in the brown planthopper. Curr. Biol. 2018, 28, 268–274. [Google Scholar] [CrossRef]
- Manjon, C.; Troczka, B.J.; Zaworra, M.; Beadle, K.; Randall, E.; Hertlein, G.; Singh, K.S.; Zimmer, C.T.; Homem, R.A.; Lueke, B.; et al. Unravelling the molecular determinants of bee sensitivity to neonicotinoid insecticides. Curr. Biol. 2018, 28, 1137–1143. [Google Scholar] [CrossRef]
- Troczka, B.J.; Homem, R.A.; Reid, R.; Beadle, K.; Kohler, M.; Zaworra, M.; Field, L.M.; Williamson, M.S.; Nauen, R.; Bass, C.; et al. Identification and functional characterisation of a novel N-cyanoamidine neonicotinoid metabolising cytochrome P450, CYP9Q6, from the buff-tailed bumblebee Bombus terrestris. Insect Biochem. Mol. Biol. 2019, 111, 103171. [Google Scholar] [CrossRef]
- Daborn, P.J.; Lumb, C.; Harrop, T.W.R.; Blasetti, A.; Pasricha, S.; Morin, S.; Michell, S.N.; Donnelly, M.J.; Muller, P.; Batterham, P. Using Drosophila melanogaster to validate metabolism-based insecticide resistance from insect pests. Insect Biochem. Mol. Biol. 2012, 42, 918–924. [Google Scholar] [CrossRef]
- Riveron, J.M.; Irving, H.; Ndula, M.; Barnes, K.G.; Ibrahim, S.; Cuamba, N.; Paine, M.J.I.; Wondji, C. Directionally selected P450 allelles are driving the spread of pyrethroid resistance in the major malaria vector Anopheles funestus. Proc. Natl. Acad. Sci. USA 2013, 110, 252–257. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Riveron, J.M.; Bibby, J.; Irving, H.; Yunta, C.; Paine, M.J.I.; Wondji, C.S. Allelic variation of cytochrome P450s drives resistance to bednet insecticides in a major malaria vector. PLoS Genet. 2015, 11, e1005618. [Google Scholar] [CrossRef] [PubMed]
- Ishak, I.H.; Riveron, J.M.; Ibrahim, S.S.; Stott, R.; Longbottom, J.; Irving, H.; Wondji, C.S. The cytochrome P450 gene CYP6P12 confers pyrethroid resistance in kdr-free Malaysian populations of the dengue vector Aedes albopictus. Sci. Rep. 2016, 6, 24707. [Google Scholar] [CrossRef] [PubMed]
- Tsakireli, D.; Riga, M.; Kounadi, S.; Douris, V.; Vontas, J. Functional characterization of CYP6A51, a cytochrome P450 associated with pyrethroid resistance in the Mediterranean fruit fly Ceratitis capitata. Pestic. Biochem. Physiol. 2019, 157, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Vassiliou, V.A.; Kitsis, P. Acaricide resistance in Tetranychus urticae (Acari: Tetranychidae) populations from Cyprus. J. Econ. Entomol. 2013, 106, 1848–1854. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, C.B.S.; Andrade, F.H.N.; Rodrigues, A.R.S.; Siqueira, H.A.A. Resistance in field populations of Tetranychus urticae to acaricides and characterization of the inheritance of abamectin resistance. Crop Prot. 2015, 67, 77–83. [Google Scholar] [CrossRef]
- Çağatay, N.S.; Menault, P.; Riga, M.; Vontas, J.; Ay, R. Identification and characterization of abamectin resistance in Tetranychus urticae Koch populations from greenhouses in Turkey. Crop Prot. 2018, 112, 112–117. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Primer Name | Sequence (5′–3′) | Product Size (bp) | Reference |
|---|---|---|---|
| CYP392A16_F | AAATACCGAGGTCGGACGTA | 117 | [20] |
| CYP392A16_R | AAGCACTTTTTCAATCTGGTCAC | ||
| RPL11_Dm_F | CGATCCCTCCATCGGTATCT | 120 | [45] |
| RPL11_Dm_R | AACCACTTCATGGCATCCTC | ||
| pUASTF | TATGTCACACCACAGAAGTAAG | n/a | [47] |
| pUASTR | CAAGTAAATCAACTGCAACTACTG |
| Cross | Transgene Dosage | LC50 (95% FL) (mg/L) | Slope | χ2 (df) | RR 1 (95% FL) | |||
|---|---|---|---|---|---|---|---|---|
| Female | Male | Gal4 | CPR | A16 | ||||
| yw | HR-GAL4 | 1 | - | - | 45.5 (33.3–56.9) | 1.9 ± 0.35 | 12.1 (16) | |
| UAS-CYP392A16.71 | HR-GAL4 | 1 | - | 1 | 53.4 (38.7–76.2) | 1.316 ± 0.25 | 17.8 (16) | 1.17 (0.81–1.71) |
| UAS-TuCPR92; UAS-CYP392A16.71 | HR-GAL4 | 1 | 1 | 1 | 85.2 (78.6–92.7) | 7.1 ± 1.6 | 10.9 (16) | 1.88 (1.44–2.44) |
| UAS-TuCPR32; UAS-CYP392A16.71 | HR-GAL4 | 1 | 1 | 1 | 101.8 (88.6–127.6) | 3.7 ± 0.62 | 10.2 (15) | 2.24 (1.65–3.04) |
| HR-GAL4; UAS-CYP392A16.71 | HRGAL4; UAS-CYP392A16.71 | 2 | - | 2 | 45.07 (37.85–51.91) | 3.41 ± 0.43 | 7.98 (10) | 0.99 (0.74–1.33) |
| UAS-TuCPR32; UAS-CYP392A16.71 | HR-GAL4; UAS-CYP392A16.71 | 1 | 1 | 2 | 77.01 (65.99–89.73) | 4.37 ± 0.47 | 14.4 (10) | 1.69 (1.29–2.23) |
| UAS-CYP392A16.9 | HR-GAL4 | 1 | - | 1 | 28.3 (10.3–42) | 2.1 ± 0.4 | 40.3 (15) | 0.62 (0.42–0.93) |
| UAS-TuCPR92; UAS-CYP392A16.9 | HR-GAL4 | 1 | 1 | 1 | 54.4 (40.1–64.3) | 4.9 ± 0.8 | 30.3 (16) | 1.19 (0.89–1.59) |
| UAS-TuCPR32; UAS-CYP392A16.9 | HR-GAL4 | 1 | 1 | 1 | 82.7 (61.7–117.5) | 3.1 ± 0.4 | 49.6 (13) | 1.82 (1.37–2.42) |
| HR-GAL4; UAS-CYP392A16.9 | HR-GAL4; UAS-CYP392A16.9 | 2 | - | 2 | 31.06 (18.82–40.36) | 3.2 ± 0.5 | 14.88 (9) | 0.68 (0.79–0.96) |
| UAS-TuCPR32; UAS-CYP392A16.9 | HR-GAL4; UAS-CYP392A16.9 | 1 | 1 | 2 | 105.29 (79.91–135.06) | 4.6 ± 0.6 | 17.4 (8) | 2.32 (1.74–3.08) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riga, M.; Ilias, A.; Vontas, J.; Douris, V. Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects 2020, 11, 829. https://doi.org/10.3390/insects11120829
Riga M, Ilias A, Vontas J, Douris V. Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects. 2020; 11(12):829. https://doi.org/10.3390/insects11120829
Chicago/Turabian StyleRiga, Maria, Aris Ilias, John Vontas, and Vassilis Douris. 2020. "Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila" Insects 11, no. 12: 829. https://doi.org/10.3390/insects11120829
APA StyleRiga, M., Ilias, A., Vontas, J., & Douris, V. (2020). Co-Expression of a Homologous Cytochrome P450 Reductase Is Required for In Vivo Validation of the Tetranychus urticae CYP392A16-Based Abamectin Resistance in Drosophila. Insects, 11(12), 829. https://doi.org/10.3390/insects11120829