Search for Alternative Control Strategies of Drosophila suzukii (Diptera: Drosophilidae): Laboratory Assays Using Volatile Natural Plant Compounds



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Chemical Substances
2.3. Bioassays
2.3.1. Contact Toxicity
2.3.2. Capillary Feeding
2.3.3. Oviposition Assay
2.4. Chemical Analyses
2.5. Statistical Analyses
3. Results
3.1. Bioassays
3.1.1. Contact Toxicity
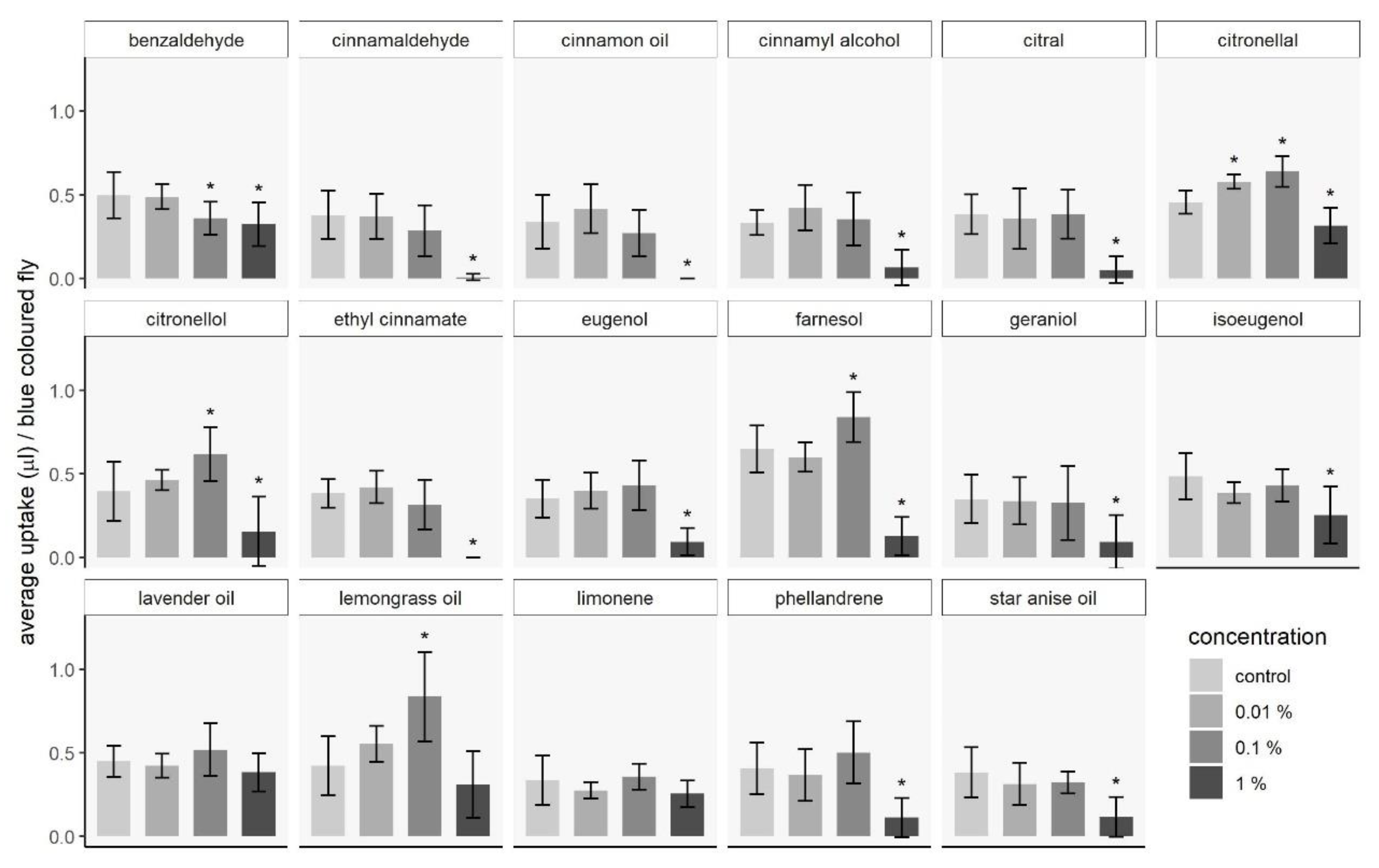
3.1.2. Capillary Feeding Test
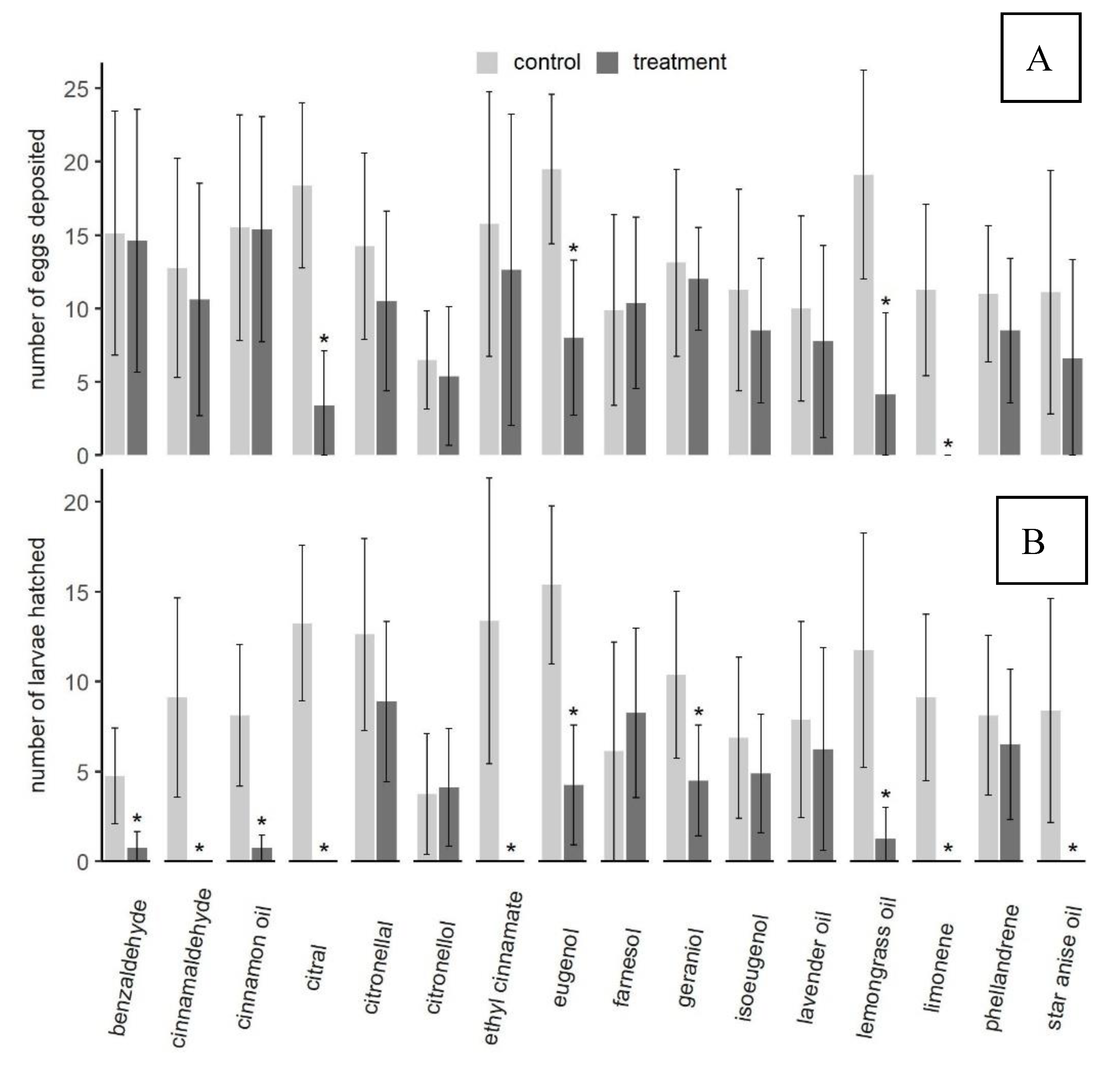
3.1.3. Oviposition Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vogt, H.; Hoffmann, C.; Baufeld, P. Ein neuer Schädling, die Kirschessigfliege, Drosophila suzukii (Matsumura), bedroht Obst-und Weinkulturen. Entomol. Nachr. Ber. 2012, 56, 191–196. [Google Scholar]
- SENASICA—Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria. Mosca del Vinagre de Alas Manchadas (Drosophila Suzukii, Matsumura, 1931); Ficha Técnica 7; Dirección General de Sanidad Vegetal—Sistema Nacional de Vigilancia Epidemiológica Fitosanitaria: Mexico City, Mexico, 2013; 22p. [Google Scholar]
- Deprá, M.; Poppe, J.L.; Schmitz, H.J.; De Toni, D.D.; Valente, V.L. The first record of the invasive pest Drosophila suzukii in the South American continent. J. Pest Sci. 2014, 87, 379–383. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- González, G.; Mary, A.L.; Goñi, B. Drosophila suzukii (Matsumura) found in Uruguay. Dros. Inf. Serv. 2015, 98, 103–107. [Google Scholar]
- Medina-Muñoz, M.C.; Lucero, X.; Severino, C.; Cabrera, N.; Olmedo, D.; Del Pino, F.; Alvarez, E.; Jara, C.; Godoy-Herrera, R. Drosophila suzukii arrived in Chile. Dros. Inf. Serv. 2015, 98, 75. [Google Scholar]
- Hamby, K.A.; Bellamy, D.E.; Chiu, J.C.; Lee, J.C.; Walton, V.M.; Wiman, N.G.; York, R.M.; Biondi, A. Biotic and abiotic factors impacting development, behaviour, phenology, and reproductive biology of Drosophila suzukii. J. Pest Sci. 2016, 89, 605–619. [Google Scholar] [CrossRef]
- Wiman, N.G.; Dalton, D.T.; Anfora, G.; Biondi, A.; Chiu, J.C.; Daane, K.M.; Gerdeman, B.; Gottardello, A.; Hamby, K.A.; Isaacs, R.; et al. Drosophila suzukii population response to environment and management strategies. J. Pest Sci. 2016, 89, 653–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eben, A.; Reifenrath, M.; Briem, F.; Pink, S.; Vogt, H. Response of Drosophila suzukii (Diptera: Drosophilidae) to extreme heat and dryness. Agric. Forest Entomol. 2017, 20, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Briem, F.; Dominic, A.R.; Golla, B.; Hoffmann, C.; Englert, C.; Herz, A.; Vogt, H. Explorative data analysis of Drosophila suzukii trap catches from a seven-year monitoring program in Southwest Germany. Insects 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, H.; Van Timmeren, S.; Wetzel, W.; Isaacs, R. Predicting within-and between-year variation in activity of the invasive spotted wing drosophila (Diptera: Drosophilidae) in a temperate region. Environ. Entomol. 2019, 48, 1223–1233. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Dalton, D.T. Analysis of the invasiveness of spotted wing Drosophila (Drosophila suzukii) in North America, Europe, and the Mediterranean Basin. Biol. Invasions 2016, 18, 3647–3663. [Google Scholar] [CrossRef]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The making of a pest: The evolution of a fruit-penetrating ovipositor in Drosophila suzukii and related species. Proc. R. Soc. B 2014, 281, 20132840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gress, B.E.; Zalom, F.G. Identification and risk assessment of spinosad resistance in a California population of Drosophila suzukii. Pest Manag. Sci. 2018, 75, 1270–1276. [Google Scholar] [CrossRef]
- Biondi, A.; Desneux, N.; Siscaro, G.; Zappalà, L. Using organic-certified rather than synthetic pesticides may not be safer for biological control agents: Selectivity and side effects of 14 pesticides on the predator Orius laevigatus. Chemosphere 2012, 87, 803–812. [Google Scholar] [CrossRef]
- Uhl, P.; Brühl, C.A. The impact of pesticides on flower-visiting insects: A review with regard to European risk assessment. Environ. Tox. Chem. 2019, 38, 2355–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, L.; Moses, J.; Hanson, E.; Fanning, P.; Isaacs, R. Rapid harvest schedules and fruit removal as non-chemical approaches for managing spotted wing drosophila. J. Pest Sci. 2017. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Lee, K.Z.; Otto, S.; Talmann, L.; Stökl, J.; Degenkolb, T.; Vilcinskas, A.; Halitschke, R. Environmentally sustainable pest control options for Drosophila suzukii. J. Appl. Entomol. 2018, 142, 3–17. [Google Scholar] [CrossRef]
- Boehnke, B.; Köppler, K.; Augel, C.; Wichura, A.; Lindstaedt, J.; Wiebusch, J.H.; Engel, A.; Benz, S.; Vogt, H. Demonstration project “Exclusion netting for managing spotted wing Drosophila in fruit crops”. Results 2017. IOBC-WPRS Bull. 2019, 144, 78–84. [Google Scholar]
- Klick, J.; Rodriguez-Saona, C.R.; Hernández Cumplido, J.; Holdcraft, R.J.; Urrutia, W.H.; Da Silva, R.O.; Borges, R.; Nafra-Neto, A.; Seagraves, M.P. Testing a novel attract-and-kill strategy for Drosophila suzukii (Diptera: Drosophilidae) management. J. Insect Sci. 2019, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Kuesel, R.; Scott Hicks, D.; Archer, K.; Sciligo, A.; Bessin, R.; Gonthier, D. Effects of fine-mesh exclusion netting on pests of blackberry. Insects 2019, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Kinjo, H.; Kunimi, Y.; Ban, T.; Makai, M. Oviposition efficacy of Drosophila suzukii (Diptera: Drosophilidae) on different cultivars of blueberry. J. Econ. Entomol. 2013, 106, 1767–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguez-Saona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revadi, S.; Vitagliano, S.; Rossi Stacconi, M.V.; Ramasamy, S.; Mansourian, S.; Carlin, S.; Vrhovsek, U.; Becher, P.G.; Mazzoni, V.; Rota-Stabelli, O.; et al. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Lee, J.C.; Dalton, D.T.; Swoboda-Bhattarai, K.A.; Bruck, D.J.; Burrack, H.J.; Strik, B.C.; Woltz, J.M.; Walton, V.M. Characterization and manipulation of fruit susceptibility to Drosophila suzukii. J. Pest Sci. 2016, 89, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Rossi-Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Anfora, G. Augmentative releases of Trichopria drosophilae for the suppression of early season Drosophila suzukii populations. BioControl 2018, 64, 9–19. [Google Scholar] [CrossRef]
- Lee, J.C.; Wang, X.; Daane, K.M.; Hoelmer, K.A.; Isaacs, R.; Sial, A.A.; Walton, V.M. Biological control of spotted-wing drosophila (Diptera: Drosophilidae)—Current and pending tactics. J. Integr. Pest Manag. 2019, 10, 22. [Google Scholar] [CrossRef]
- Wolf, S.; Boycheva-Woltering, S.; Romeis, J.; Collatz, J. Trichopria drosophilae parasitizes Drosophila suzukii in seven common non-crop fruits. J. Pest Sci. 2020, 93, 627–638. [Google Scholar] [CrossRef]
- Englert, C.; Herz, A. Zur Bedeutung heimischer Drosophila-Parasitoide für die Regulierung von Drosophila suzukii—Vom Freiland über das Labor zum Freiland. In Proceedings of the Deutsche Gesellschaft für allgemeine und angewandte Entomologie, Halle (Saale), Germany, 11–14 March 2019; pp. 45–46. [Google Scholar]
- Vogt, H.; Boehnke, B.; Saltzmann, J.; Eberhardt, G.; Wichura, A.; Wiebusch, J.H.; Lindstaedt, J.; Kramer, K.; Engel, A.; Benz, S. Demonstrationsvorhaben Einnetzen von Obstkulturen zum Schutz gegen die Kirschessigfliege (Drosophila suzukii). Förderkennzeichen: 2815MD010, Ergebnisse des zweiten Projektjahres 2018; Berichtszeitraum 1.März 2018–28.Februar 2019. Open Agrar 2019. [Google Scholar] [CrossRef]
- Basoalto, E.; Hilton, R.; Knight, A. Factors affecting the efficacy of a vinegar trap for Drosophila suzukii (Diptera: Drosophilidae). J. Appl. Entomol. 2013, 137, 561–570. [Google Scholar] [CrossRef]
- Hamby, K.A.; Becher, P.G. Current knowledge of interactions between Drosophila suzukii and microbes, and their potential utility for pest management. J. Pest Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Belutti, N.; Gallmetzer, A.; Innerebner, G.; Schmidt, S.; Zelger, T.; Koschier, E.H. Dietary yeast affects preference and performance in Drosophila suzukii. J. Pest Sci. 2018, 91, 651–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrack, J.H.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C.; et al. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Mori, B.A.; Whitener, A.B.; Leinweber, Y.; Revadi, S.; Beers, E.H.; Witzgall, P.; Becher, P.G. Enhanced yeast feeding following mating facilitates control of the invasive fruit pest Drosophila suzukii. J. Appl. Ecol. 2017, 54, 170–177. [Google Scholar] [CrossRef]
- Tonina, L.; Grassi, A.; Caruso, S.; Mori, M.; Gottardello, A.; Anfora, G.; Giomi, F.; Vaccari, G.; Ioriatti, C. Comparison of attractants for Drosophila suzukii monitoring in sweet cherry orchards in Italy. J. Appl. Entomol. 2018, 142, 8–25. [Google Scholar] [CrossRef]
- Cha, D.H.; Adams, T.; Rogg, H.; Landolt, P.J. Identification and field evaluation of fermentation volatiles from wine and vinegar that mediate attraction of spotted wing Drosophila, Drosophila suzukii. J. Chem. Ecol. 2012, 38, 1419–1431. [Google Scholar] [CrossRef]
- Cha, D.H.; Hesler, S.P.; Cowles, R.S.; Vogt, H.; Loeb, G.M.; Landolt, P.J. Comparison of a synthetic chemical lure and standard fermented baits for trapping Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2013, 42, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Cha, D.H.; Adams, T.; Werle, C.T.; Sampson, B.J.; Adamczyk, J.J.; Rogg, H.; Landolt, P.J. A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest Manag. Sci. 2014, 70, 324–331. [Google Scholar] [CrossRef]
- Cha, D.H.; Landolt, P.J.; Adams, T.B. Effect of chemical ratios of a microbial-based attractant on trap catch of Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2017, 46, 907–915. [Google Scholar] [CrossRef]
- Frewin, A.J.; Enkema, J.R.; Fraser, H.; Hallett, R.H. Evaluation of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2017, 110, 1156–1163. [Google Scholar] [CrossRef]
- Feng, Y.; Bruton, R.; Park, A.; Zhang, A. Identification of attractive blend for spotted wing drosophila, Drosophila suzukii, from apple juice. J. Pest Sci. 2018, 91, 1251–1267. [Google Scholar] [CrossRef] [Green Version]
- Cloonan, K.R.; Hernández-Cumplido, J.; Viana de Sousa, A.L.; Ramalho, D.G.; Burrack, H.J.; Della Rosa, L.; Diepenbrock, L.M.; Ballman, E.; Drummond, F.A.; Gut, L.J.; et al. Laboratory and field evaluation of host-related foraging odor-cue combinations to attract Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2019, 112, 2850–2860. [Google Scholar] [CrossRef] [PubMed]
- Harmon, D.S.; Haseeb, M.; Kanga, L.H.B.; Liburd, O.E. Evaluation of monitoring traps and lures for Drosophila suzukii (Diptera: Drosophilidae) in berry plantings in Florida. Insects 2019, 10, 313. [Google Scholar] [CrossRef] [Green Version]
- Landolt, P.J.; Adams, T.; Rogg, H. Trapping spotted wing drosophila, Drosophila suzukii (Matsumura) (Diptera: Drosophilidae), with combinations of vinegar and wine, and acetic acid and ethanol. J. Appl. Entomol. 2012, 136, 148–154. [Google Scholar] [CrossRef]
- Piñero, J.C.; Barrett, B.A.; Bolton, L.G.; Follett, P.A. ß-cyclocitral synergizes the response of adult Drosophila suzukii (Diptera: Drosophilidae) to fruit juices and isoamyl acetate in a sex-dependent manner. Sci. Rep. 2019, 9, 10574. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isman, M.B.; Machial, C.M. Pesticides based on plant essential oils: From traditional practice to commercialization. In Naturally Occurring Bioactive Compounds; Rai, M., Carpinella, M., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2006; pp. 29–44. [Google Scholar]
- Gökçe, A.; Isaacs, R.; Whalon, M.E. Ovicidal, larvicidal and anti-ovipositional activities of Bifora radians and other plant extracts on the grape berry moth Paralobesia viteana (Clemens). J. Pest Sci. 2011, 84, 487–493. [Google Scholar] [CrossRef]
- Wink, M. Production and application of phytochemicals from an agricultural perspective. In Phytochemistry and Agriculture; Van Beek, T.A., Breteler, H., Eds.; Oxford University Press: Oxford, UK, 1993; Volume 34, pp. 171–213. [Google Scholar]
- Jones, G.; Campbell, C.A.M.; Hardie, J.; Pickett, J.A.; Pye, J.B.; Wadhams, L.J. Integrated management oft wo-spotted spider mite Tetranychus urticae on hops using hop-acids as an antifeedant together with the predaroy mite Pytoseiulus persimilis. Biocontrol Sci. Technol. 2003, 13, 241–252. [Google Scholar] [CrossRef]
- Zimmermann, G. Efficacy and potential use of essential oils in plant protection: A review. J. Kult. 2012, 64, 1–19. [Google Scholar]
- Attia, S.; Grissa, K.; Lognay, G.; Bitume, E.; Hance, T.; Mailleuy, A.C. A review of the major biological aproaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Nenaah, G.E. Chemical composition, insecticidal and repellence activities of essential oils of three Achillea species against the Khapra beetle (Coleoptera: Dermestidae). J. Pest Sci. 2014, 87, 273–283. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Rheault, M.R.; Mahmoud, S.S. Insecticidal and oviposition deterrent effects of essential oils and their constituents against the invasive pest Drosophila suzukii (Matsumura) (Diptera: Drosophilidae). Crop Prot. 2015, 78, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholin-esterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crop Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- Park, C.G.; Jang, M.; Shin, E.; Kim, J. Myrtaceae plant essential oils and their ß-triketone components as insecticides against Drosophila suzukii. Molecules 2017, 22, 1050. [Google Scholar] [CrossRef] [Green Version]
- Cahenzli, F.; Strack, T.; Daniel, C. Screening of 25 different natural crop protection products against Drosophila suzukii. J. Appl. Entomol. 2018, 142, 563–577. [Google Scholar] [CrossRef]
- Pérez-Guerrero, S.; Mateus, C. Field evaluation of commercial plant extracts against Drosophila suzukii. Int. J. Pest Manag. 2019, 65, 53–58. [Google Scholar] [CrossRef]
- Trombin de Souza, M.; Bernardi, D.; Krinski, D.; de Melo, D.J.; da Costa Oliveira, D.; Rakes, M.; Gorgatti Zarbin, P.H.; Lameiro de Noronha, B.H.; Maia, S.; Casilha Zawadneak, M.A. Chemical composition of essential oils of selected species of Piper and their insecticidal activity against Drosophila suzukii and Trichopria anastrephae. Environ. Sci. Pollut. Res. 2020, 27, 1–10. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Cha, D.H.; Linn, C.E., Jr.; Wolfin, M.S.; Loeb, G.M. Robust manipulations of pest insect behavior using repellents and practical application for integrated pest management. Environ. Entomol. 2017, 46, 1041–1050. [Google Scholar] [CrossRef]
- Renkema, J.M.; Buitenhuis, R.; Hallett, R.H. Reduced Drosophila suzukii infestation in berries using deterrent compounds and laminate polymer flakes. Insects 2017, 8, 117. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dong, W.; Zhang, F.; Kenis, M.; Griepink, F.; Zhang, J.; Chen, L.; Xiao, C. Identification of active components from volatiles of Chinese Bayberry, Myrica rubra attractive to Drosophila suzukii. Arthropod-Plant Int. 2018, 12, 435–442. [Google Scholar] [CrossRef]
- Jang, M.; Kim, J.; Yoon, K.A.; Lee, S.H.; Park, C.G. Biological activity of Myrtaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Pest Manag. Sci. 2017, 73. [Google Scholar] [CrossRef]
- Wallingford, A.K.; Hesler, S.P.; Cha, D.H.; Loeb, G.M. Behavioral response of spotted-wing drosophila, Drosophila suzukii Matsumura, to aversive odors and a potential oviposition deterrent in the field. Pest Manag. Sci. 2016, 72, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.B.; Short, B.D.; Leskey, T.C. Development of an attract-and-kill strategy for Drosophila suzukii (Diptera: Drosophilidae): Evaluation of attracticidal spheres under laboratory and field conditions. J. Econ. Entomol. 2017, 110, 535–542. [Google Scholar] [CrossRef]
- Lasa, R.; Toledo-Hernandez, R.A.; Rodriguez, D.; Williams, T. Raspberry as a source for the development of Drosophila suzukii attractants: Laboratory and commercial polytunnel trials. Insects 2019, 10, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallingford, A.K.; Connelly, H.L.; Brind’Amour, G.D.; Boucher, M.T.; Mafra-Neto, A.; Loeb, G.M. Field evaluation of an oviposition deterrent for management of spotted-wing drosophila, Drosophila suzukii, and potential nontarget effects. J. Econ. Entomol. 2016, 109, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.Y.; Hwang, S.T.; Sethi, G.; Fan, L.; Arfuso, F.; Ahn, K.S. Potential anti-inflammatory and anti-cancer proterties of farnesol. Molecules 2018, 23, 2827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedini, S.; Flamini, G.; Cosci, F.; Ascrizzi, R.; Cioni, P.L.; Ioriatti, C.; Conti, B.; Lucchi, A. Repellency of aromatic plant essential oils against Drosophila suzukii. IOBC-WPRS Bull. 2017, 128, 13–17. [Google Scholar]
- Preedy, V.R. Essential Oils in Food Preservation, Flavor and Safety; Academic Press: Amsterdam, The Netherlands, 2016; 930p. [Google Scholar]
- Ja, W.W.; Carvalho, G.B.; Mak, E.M.; De la Rosa, N.N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of Drosophila and the CAFE assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8250–8256. [Google Scholar] [CrossRef] [Green Version]
- Jacas, J.A.; Viñuela, E. Analysis of a laboratory method to test the effects of pesticides on adult females of Opius concolor (Hym., Braconidae), a parasitoid of the olive fruit fly, Bactrocera oleae (Dip., Tephritidae). Biocontrol Sci. Technol. 1984, 4, 147–154. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www-R-project.org/ (accessed on 28 October 2020).
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arppe, A.; Borchers, H.W.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for Descriptive Statistics. Available online: https://cran.r-project.org/web/packages/DescTools/DescTools.pdf (accessed on 12 March 2020).
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Mazzetto, F.; Gonella, E.; Crotti, E.; Vacchini, V.; Syrpas, M.; Pontini, M.; Mangelinckx, S.; Daffonchio, D.; Alma, A. Olfactory attaction of Drosophila suzukii by symbiotic acetic acid bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Sampson, B.J.; Werle, C.T.; Stringer, S.J.; Adamczyk, J.J. Ingestible insecticides for spotted wing Drosophila control: A polyol, Erythritol, and an insect growth regulator, Lefenuron. J. Appl. Entomol. 2016, 141, 8–18. [Google Scholar] [CrossRef]
- Yousef, M.; Aranda-Valera, E.; Quesada-Moraga, E. Lure-and-infect and lure-and-kill devices based on Metarhizium brunneum for spotted wing drosophila control. J. Pest Sci. 2017, 91, 227–235. [Google Scholar] [CrossRef]
- Wollmann, J.; Schlesener, D.H.C.; Vieira, J.G.A.; Bernardi, D.; Garcia, M.S.; Garcia, F.R.M. Evaluation of food baits to capture Drosophila suzukii in the southern of Brazil. An. Acad. Bras. Cienc. 2019, 91, e20180375. [Google Scholar] [CrossRef]
- Spitaler, U.; Bianchi, F.; Eisenstecken, D.; Castellan, I.; Angeli, S.; Dordevic, N.; Robatscher, P.; Vogel, R.F.; Hoschiers, E.H.; Schmidt, S. Yeast species affects feeding and fitness of Drosophila suzukii adults. J. Pest Sci. 2020. [Google Scholar] [CrossRef]
- Sarma, R.; Adhikari, K.; Mahanta, S.; Khanikor, B. Insecticidal activities of Citrus aurantifolia essential oil against Aedes aegypti (Diptera: Culicidae). Toxicol. Rep. 2019, 6, 1091–1096. [Google Scholar] [CrossRef]
- Bedini, S.; Cosci, F.; Tani, C.; Pierattini, E.C.; Venturi, F.; Lucchi, A.; Ioriatti, C.; Ascrizzi, R.; Flamini, G.; Ferroni, G.; et al. Essential oils as post-harvest crop protectants against the fruit fly Drosophila suzukii: Bioactivity and organoleptic profile. Insects 2020, 11, 508. [Google Scholar] [CrossRef]
- Dam, D.; Molitor, D.; Beyer, M. Natural compounds for controlling Drosophila suzukii. A review. Agron. Sustain. Dev. 2019, 39, 53. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Nerio, L.S.; Oliver-Verbel, J.; Stshenko, E. Repellent activity of essential oils: A review. Bioresour. Technol. 2010, 101, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jang, M.; Lee, K.T.; Yoon, K.A.; Park, C.G. Insecticidal and enzyme inhibitory activities of sparassol and its analogues against Drosophila suzukii. J. Agric. Food Chem. 2016, 64, 5479–5483. [Google Scholar] [CrossRef] [PubMed]
- Suwannayod, S.; Sukontason, K.L.; Pitasawat, B.; Junkum, A.; Limsophatham, K.; Jones, M.K.; Somboon, P.; Leksomboon, R.; Chareonviriyaphap, T.; Tawatsin, A.; et al. Synergistic toxicity of plant essential oils combined with pyrethroid insecticides against blowflies and the house fly. Insects 2019, 10, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossentine, J.E.; Ayyanath, M.M. Limited protection of the parasitoid Pachycrepoideus vindemmiae from Drosophila suzukii host-directed spinosad suppression. Entomol. Exp. Appl. 2017, 164, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Schlesener, D.C.H.; Wollmann, J.; De Bastos Pazini, J.; Padilha, A.C.; Grützmacher, A.D.; Mello Garcia, F.R. Insecticide toxicity to Drosophila suzukii (Diptera: Drosophilidae) parasitoids: Trichopria anastrephae (Hymenoptera: Diapriidae) and Pachycrepoideus vindemmiae (Hymenoptera: Pteromalidae). J. Econ. Entomol. 2019, 112, 1197–1206. [Google Scholar] [CrossRef]
- Gowton, C.M.; Reut, M.; Carillo, J. Peppermint esential oil inhibits Drosophila suzukii emergence but reduces Pachycrepoideus vindemmiae parasitism rates. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Hamby, K.A.; Kwok, R.S.; Zalom, F.G.; Chiu, J.H.C. Integrating circadian activity and gene expression profiles to predict chronotoxicity of Drosophila suzukii response to insecticides. PLoS ONE 2013, 8, e68472. [Google Scholar] [CrossRef] [Green Version]
- Wallingford, A.K.; Cha, D.H.; Loeb, G.M. Evaluating a push–pull strategy for management of Drosophila suzukii Matsumura in red raspberry. Pest Manag. Sci. 2018, 74, 120–125. [Google Scholar] [CrossRef]
- Cloyd, R.A.; Galle, C.L.; Ketith, S.R.; Kalscheur, N.A.; Kemp, K.E. Effect of commercially available plant-derived essential oil products on arthropod pests. J. Econ. Entomol. 2009, 102, 1567–1579. [Google Scholar] [CrossRef]
- Cloonan, K.R.; Abraham, J.; Angeli, S.; Zainulabeuddin, S.; Rodriguez-Saona, C. Advances in the chemical ecology of the spotted wing drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 2018, 44, 922–939. [Google Scholar] [CrossRef]
- Wong, J.S.; Wallingford, A.K.; Loeb, G.M.; Lee, J.C. Physiological status of Drosophila suzukii (Diptera: Drosophilidae) affects their response to attractive odours. J. Appl. Entomol. 2018, 142, 473–482. [Google Scholar] [CrossRef]
- Clymans, R.; Van Kerckvoorde, V.; Banghes, E.; Akkermans, W.; Alhmedi, A.; DeClercq, P.; Beliën, T.; Bylemans, D. Olfactory preference of Drosophila suzukii shifts between fruit and fermentation cues over the season: Effects of physiological status. Insects 2019, 10, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, D.M.; Leach, H.L.; Xu, P.; Dong, K.; Isaacs, R.; Gut, L.J. Comparative antennal and behavioral responses of summer and winter morph Drosophila suzukii (Diptera: Drosophilidae) to ecologically relevant volatiles. Environ. Entomol. 2018, 47, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Burkness, E.C.; Hutchison, W.D. Evaluation of high tunnels for management of Drosophila suzukii in fall-bearing red raspberries: Potential for reducing insecticide use. J. Pest Sci. 2016, 89, 815–821. [Google Scholar] [CrossRef]
- Augel, C.; Boehnke, B.; Wichura, A.; Lindstaedt, J.; Wiebusch, J.H.; Engel, A.; Benz, S.; Saltzmann, J.; Eberhardt, G.; Vogt, H.; et al. Demonstration project exclusion netting for managing spotted wing drosophila in fruit crops—Results 2017–2019. In Proceedings of the Ecofruit. 19th International Conference on Organic Fruit Growing, Hohenheim, Germany, 17–19 February 2020; pp. 166–169. [Google Scholar]
- Fanning, P.D.; Grieshop, M.J.; Isaacs, R. Efficacy of biopesticides on spotted wing drosophila, Drosophila suzukii Matsumura, in fall red raspberries. J. Appl. Entomol. 2017, 142, 26–32. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compound | Company | Purity |
|---|---|---|
| Benzaldehyde | Merck | >99% |
| Cinnamon oil | Fluka | Pure essential oil |
| Cinnamon alcohol | Sigma Aldrich | 98% |
| Cinnamaldehyde | Fluka | 99% |
| Citral | Fluka | >99% |
| Citronellal | Fluka | >80% |
| Citronellol | Fluka | >95% |
| Ethyl cinnamate | Merck | >98% |
| Eugenol | Sigma Aldrich | >99% |
| Farnesol | Sigma Aldrich | >95% |
| Geraniol | Fluka | >96% |
| Isoeugenol | Sigma Aldrich | Cis-trans-mix, 98% |
| Lavender oil | Taomed | 100% |
| Lemongrass oil | Fluka | Pure essential oil |
| R (+)—limonene | Sigma Aldrich | >95% |
| Phellandrene | Fluka | >99% |
| Star anise oil | Dragoco | Pure essential oil |
| Test Compounds | LC90 after 1 h (95% CI) | LC90 after 4 h (95% CI) | LC90 after 24 h (95% CI) | |||
|---|---|---|---|---|---|---|
| Cinnamaldehyde | 3.84 | (2.73–4.95) | 2.92 | (2.18–3.67) | 2.80 | (2.10–3.50) |
| Cinnamon alcohol | 3.66 | (2.59–4.73) | 2.14 | (0.34–3.95) | 2.12 | (0.92–3.32) |
| Cinnamon oil | 5.62 | (4.58–6.65) | 4.26 | (3.34–5.18) | 3.98 | (3.11–4.84) |
| Citral | 7.54 | (6.45–8.62) | 8.17 | (7.07–9.26) | 7.64 | (6.47–8.82) |
| Citronellol | >10.00 | (10.04–14.82) | >10.00 | (10.29–19.28) | >10.00 | (9.95–28.49) |
| Ethyl cinnamate | 2.68 | (0.95–4.40) | <2.00 | (0.00–1.34) | 2.90 | (1.99–3.81) |
| Eugenol | 4.01 | (3.34–4.67) | 4.95 | (4.05–5.86) | 4.85 | (3.97–5.72) |
| Farnesol | >10.00 | (10.26–22.79) | 9.47 | (7.20–11.73) | 6.65 | (5.24–8.06) |
| Feraniol | 7.87 | (7.31–8.42) | 8.28 | (7.70–8.87) | 8.74 | (7.68–9.81) |
| Isoeugenol | 2.05 | (1.50–2.60) | 1.93 | (1.08–2.78) | 2.05 | (1.50–2.60) |
| Lemongrass oil | 9.77 | (8.26–11.27) | >10.00 | (9.08–3.41) | 9.46 | (7.96–10.96) |
| Star anise oil | >10.00 | (7.35–19.88) | >10.00 | (0.00–99.47) | >10.00 | (8.36–40.05) |
| Test Compounds | LC90 after 1 h (95% CI) | LC90 after 4 h (95% CI) | LC90 after 24 h (95% CI) | |||
|---|---|---|---|---|---|---|
| Cinnamaldehyde—♂ | 2.75 * | (1.65–3.85) | 2.06 * | (1.54–2.57) | 2.06 * | (1.56–2.56) |
| Cinnamaldehyde—♀ | 4.60 | (3.22–5.98) | 3.55 | (2.56–4.54) | 3.32 | (2.40–4.25) |
| Cinnamyl alcohol—♂ | 2.77 * | (1.97–3.57) | 2.13 | (1.01–3.25) | 2.12 | (0.87–3.38) |
| Cinnamyl alcohol—♀ | 4.50 | (2.85–6.15) | 2.17 | (0.08–4.25) | 2.13 | (0.93–3.32) |
| Cinnamon oil—♂ | 5.58 | (4.29–6.87) | 3.62 * | (2.67–4.56) | 3.62 | (2.67–4.56) |
| Cinnamon oil—♀ | 5.59 | (4.52–6.66) | 4.78 | (3.66–5.90) | 4.29 | (3.29–5.29) |
| Citral—♂ | 6.79 | (5.64–7.93) | 7.59 | (6.33–8.84) | 6.08 * | (5.06–7.10) |
| Citral—♀ | 8.03 | (6.79–9.27) | 8.31 | (7.23–9.38) | 8.56 | (7.16–9.96) |
| Citronellol—♂ | >10.00 | (9.49–14.0) | >10.00 | (9.88–15.13) | >10.00 | (9.56–22.32) |
| Citronellol—♀ | >10.00 | (9.82–16.17) | >10.00 | (8.76–27.59) | >10.00 | (6.30–42.41) |
| Farnesol—♂ | >10.00 | (9.50–17.37) | 5.87 * | (4.50–7.24) | 4.74 * | (3.55–5.92) |
| Farnesol—♀ | >10.00 | (7.41–33.69) | >10.00 | (8.17–12.24) | 7.6 | (6.06–9.14) |
| Lemongrass oil—♂ | 9.41 | (7.52–11.30) | >10.00 | (7.89–12.24) | 8.07 | (6.62–9.53) |
| Lemongrass oil—♀ | 9.37 | (8.27–10.47) | >10.00 | (9.17–12.69) | 9.55 | (8.34–10.75) |
| Test Compounds | Control | 0.01% | 0.10% | 1.00% |
|---|---|---|---|---|
| Cinnamon oil | 4.3 (±1.4) | 84.4 (±0.7) | 63.0 (±1.8) | 30.1 (±0.4) |
| Cinnamaldehyde | 3.9 (±1.5) | 84.3 (±1.6) | 52.5 (±1.5) | 80.4 (±0.5) |
| Cinnamyl alcohol | 94.8 (±0.6) | 73.8 (±1.1) | 73.9 (±1.2) | 21.3 (±0.9) |
| Ethyl cinnamate | 4.7 (±0.7) | 94.7 (±0.7) | 63.4 (±1.6) | 60.3 (±0.5) |
| Eugenol | 4.3 (±0.7) | 4.6 (±0.7) | 3.8 (±1.2) | 1.5 (±1.0) |
| Isoeugenol | 84.3 (±0.7) | 4.6 (±0.5) | 4.6 (±0.5) | 31.8 (±1.2) |
| Lemongrass oil | 4.3 (±1.6) | 94.8 (±0.4) | 94.9 (±0.3) | 42.1 (±1.6) |
| Limonene | 4.2 (±1.9) | 84.4 (±0.7) | 94.8 (±0.6) | 63.3 (±1.3) |
| Citral | 3.3 (±1.0) | 52.8 (±1.7) | 63.3 (±1.6) | 42.1 (±1.6) |
| Citronellal | 4.8 (±0.4) | 15.0 (±0.0) | 94.7 (±0.9) | 63.2 (±1.1) |
| Citronellol | 3.8 (±1.6) | 94.7 (±0.5) | 84.4 (±1.1) | 21.4 (±1.2) |
| Geraniol | 3.8 (±1.8) | 52.9 (±2.0) | 73.5 (±2.0) | 21.3 (±0.9) |
| Farnesol | 4.8 (±0.4) | 94.9 (±0.3) | 94.7 (±0.7) | 31.9 (±1.7) |
| Lavender oil | 4.8 (±0.4) | 94.7 (±0.5) | 94.7 (±0.5) | 84.3 (±0.7) |
| Phellandrene | 4.0 (±1.5) | 4.1 (±1.6) | 84.1 (±1.6) | 42.2 (±2.2) |
| Benzaldehyde | 4.0 (±1.2) | 84.2 (±0.9) | 94.6 (±1.0) | 63.3 (±1.1) |
| Star anise oil | 3.5 (±1.7) | 84.1 (±1.5) | 15.0 (±0.0) | 31.5 (±1.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eben, A.; Sporer, F.; Vogt, H.; Wetterauer, P.; Wink, M. Search for Alternative Control Strategies of Drosophila suzukii (Diptera: Drosophilidae): Laboratory Assays Using Volatile Natural Plant Compounds. Insects 2020, 11, 811. https://doi.org/10.3390/insects11110811
Eben A, Sporer F, Vogt H, Wetterauer P, Wink M. Search for Alternative Control Strategies of Drosophila suzukii (Diptera: Drosophilidae): Laboratory Assays Using Volatile Natural Plant Compounds. Insects. 2020; 11(11):811. https://doi.org/10.3390/insects11110811
Chicago/Turabian StyleEben, Astrid, Frank Sporer, Heidrun Vogt, Pille Wetterauer, and Michael Wink. 2020. "Search for Alternative Control Strategies of Drosophila suzukii (Diptera: Drosophilidae): Laboratory Assays Using Volatile Natural Plant Compounds" Insects 11, no. 11: 811. https://doi.org/10.3390/insects11110811
APA StyleEben, A., Sporer, F., Vogt, H., Wetterauer, P., & Wink, M. (2020). Search for Alternative Control Strategies of Drosophila suzukii (Diptera: Drosophilidae): Laboratory Assays Using Volatile Natural Plant Compounds. Insects, 11(11), 811. https://doi.org/10.3390/insects11110811