Colonization Patterns, Phenology and Seasonal Abundance of the Nearctic Leafhopper Erasmoneura vulnerata (Fitch), a New Pest in European Vineyards

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
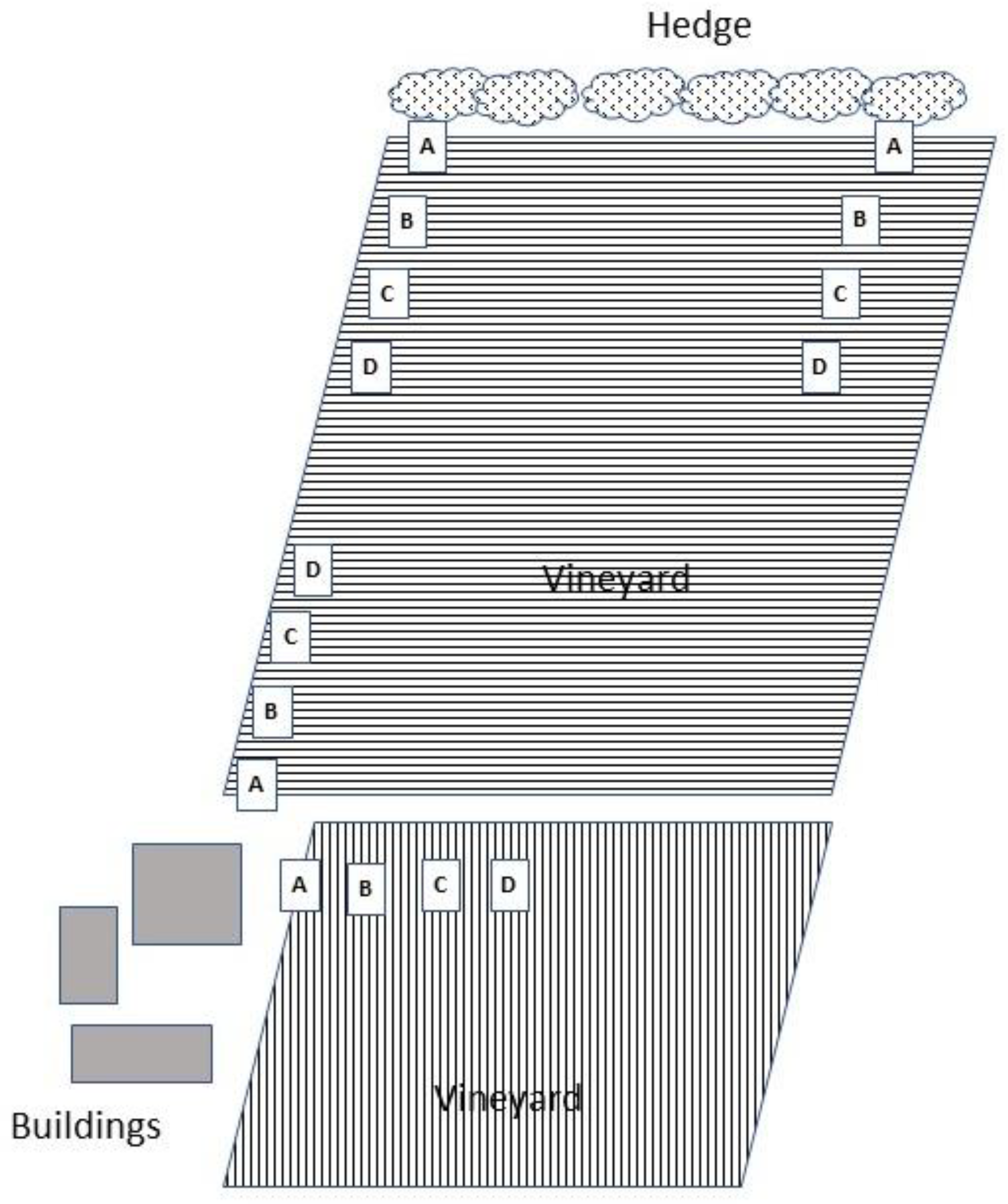
2.1. Study Sites
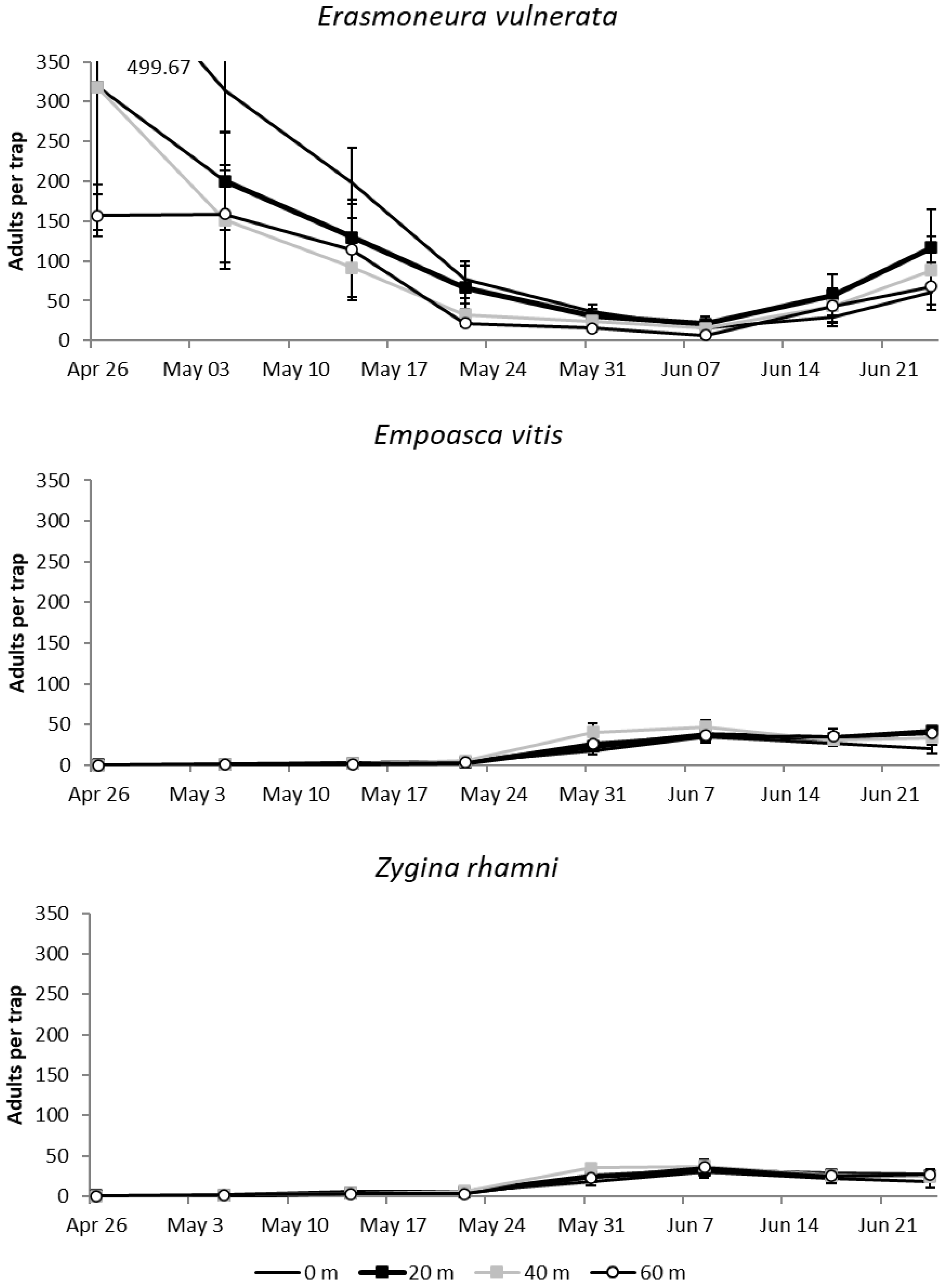
2.2. Effects of Distance from Vineyard Margins on Erasmoneura vulnerata Colonization Patterns
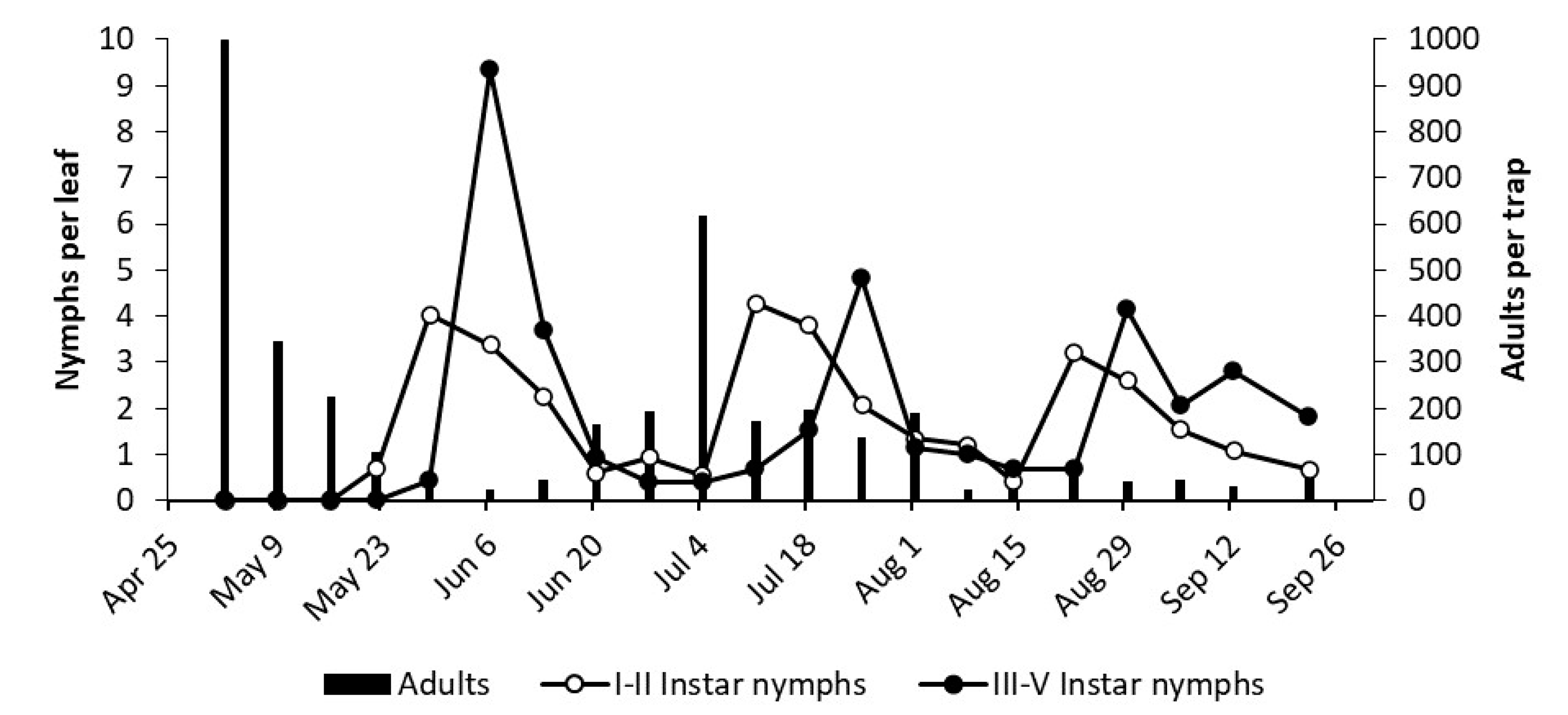
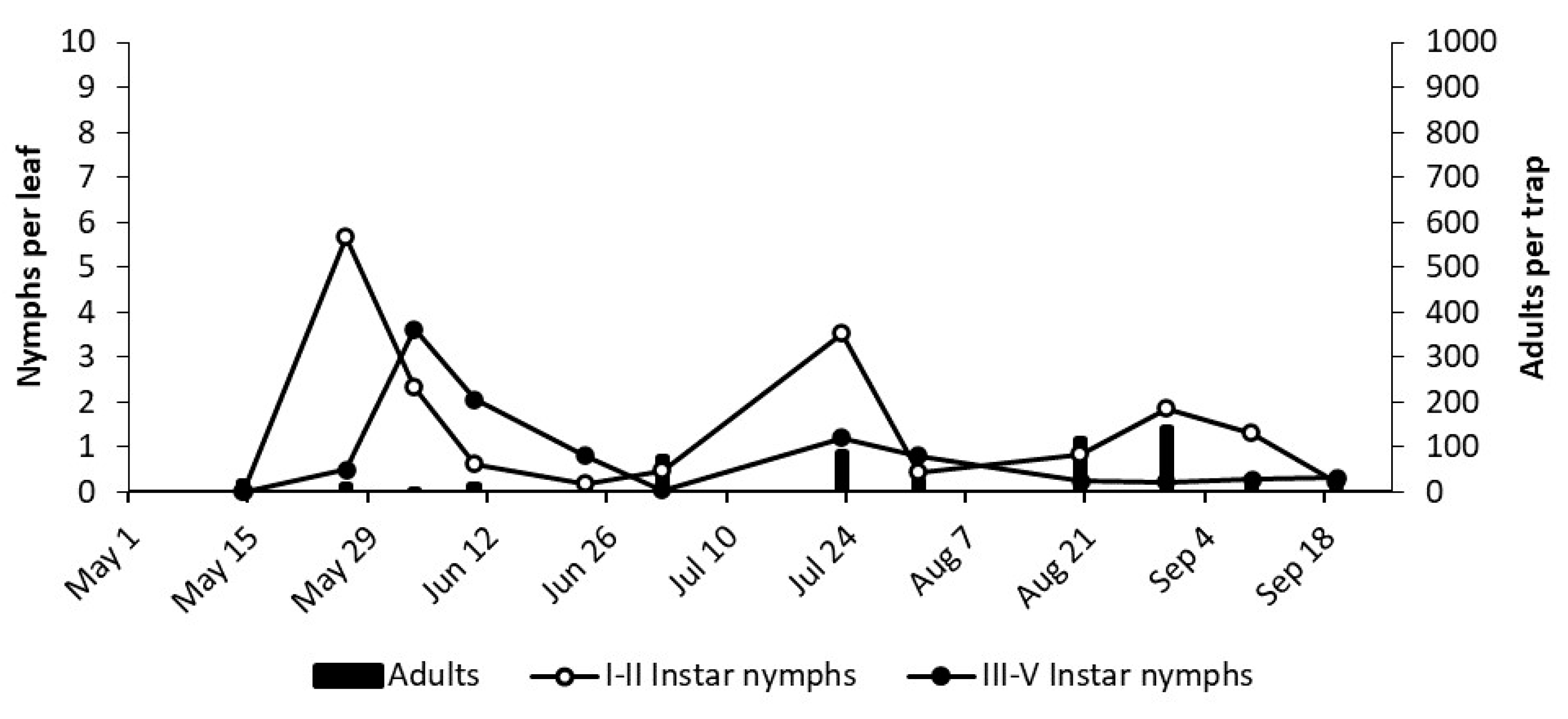
2.3. Phenology of Erasmoneura vulnerata
2.4. Effects of Vineyard Management on Erasmoneura vulnerata Seasonal Abundance
2.5. Statistical Analyses
3. Results
3.1. Effects of Distance from Vineyard Margins on Erasmoneura vulnerata Colonization Patterns
3.1.1. 2017
3.1.2. 2018
3.1.3. 2019
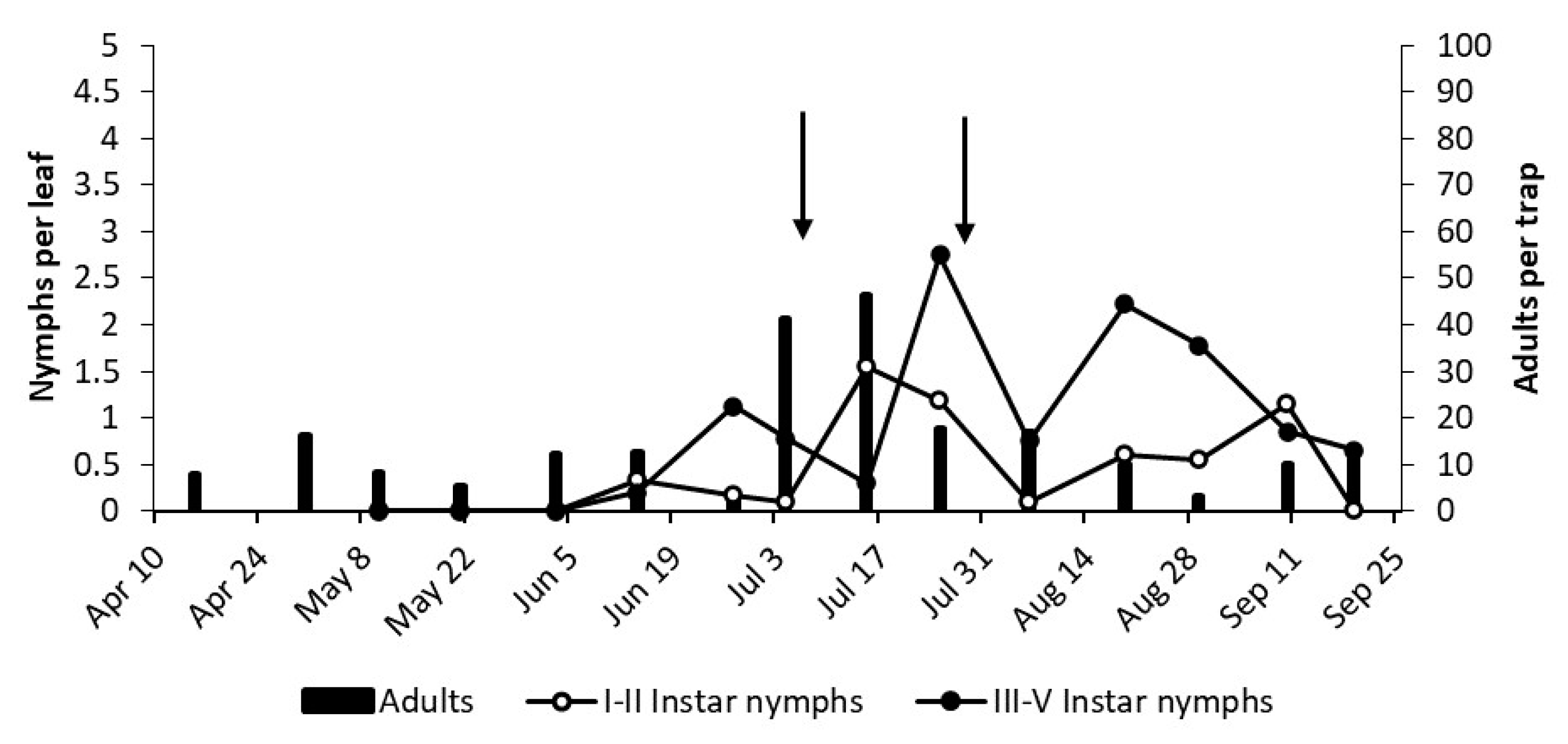
3.2. Phenology of Erasmoneura vulnerata
3.2.1. 2017
3.2.2. 2018
3.2.3. 2019
3.3. Natural Control
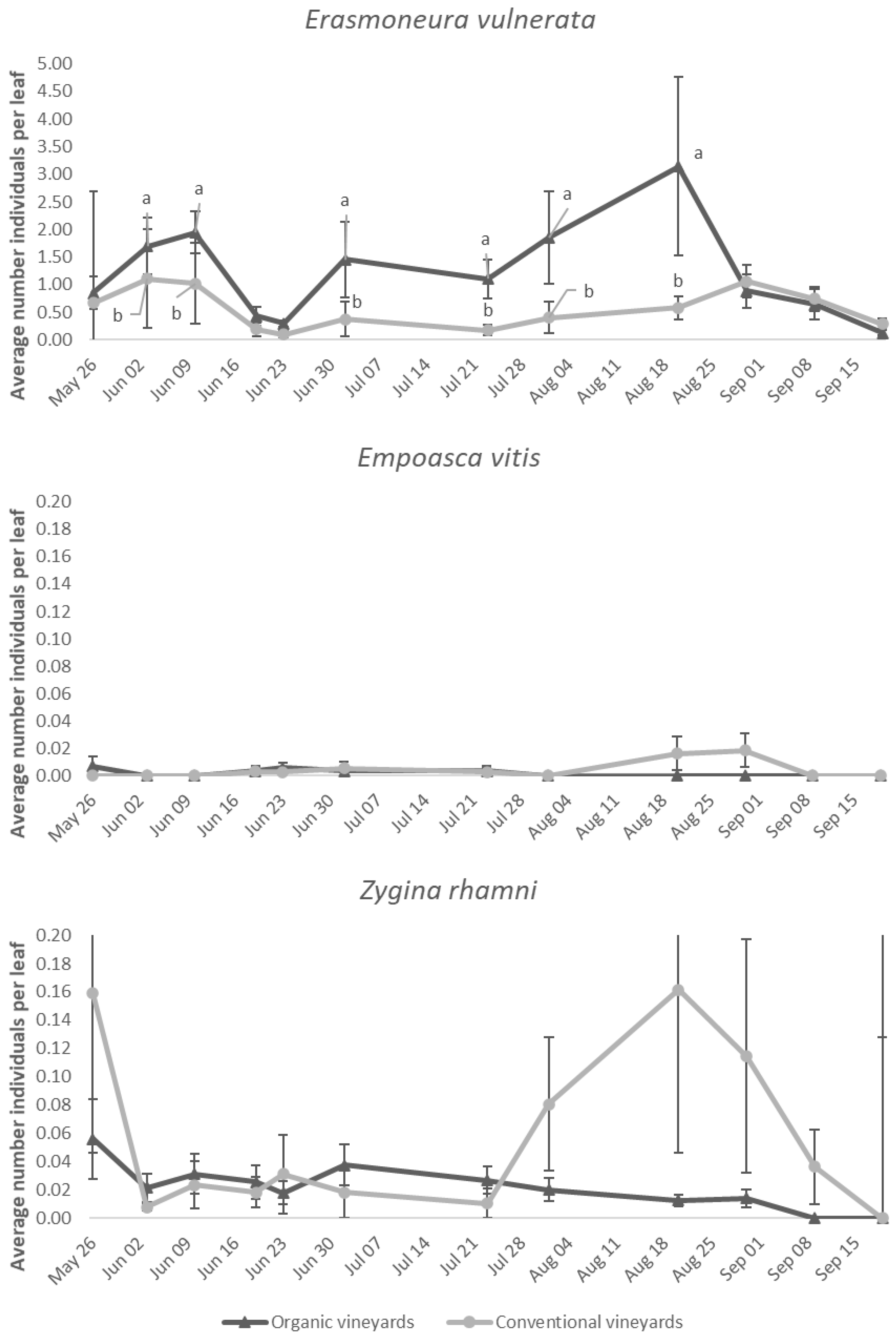
3.4. Effect of Vineyard Management
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McAtee, W.L. Key to the nearctic species and varieties of Erythroneura (Homoptera; Eupterygidae). Trans. Am. Entomol. Soc. 1920, 46, 267–321. [Google Scholar]
- Johnson, D.M. Leafhoppers of Ohio. Subfamily Typhlocybinae (Homoptera: Cicadellidae). Bull. Ohio Biol. Surv. 1935, 6, 39–122. [Google Scholar]
- Metcalf, Z.P. General Catalogue of the Homoptera. VI. Cicadelloidea. Part 17; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1968; Volume 6, p. 1513. [Google Scholar]
- Dmitriev, D.A.; Dietrich, C.H. Review of the New World Erythroneurini (Hemiptera: Cicadellidae: Typhlocybinae). I. Genera Erythroneura, Erasmoneura, Rossmoneura, and Hymetta. Ill. Nat. Hist. Surv. 2007, 38, 59–128. [Google Scholar]
- Wilson, L.T.; Barnes, M.M.; Flaherty, D.L.; Andris, H.L.; Leavitt, G.M. Variegated Grape Leafhopper. In Grape Pest Management; University of California, Division of Agriculture and Natural Resources: Oakland, CA, USA, 1992; Volume 3343, pp. 202–213. [Google Scholar]
- Daane, K.M.; Yokota, G.Y.; Zheng, Y.; Hagen, K.S. Inundative release of common green lacewings (Neuroptera: Chrysopidae) to suppress Erythroneura variabilis and E. elegantula. (Homoptera: Cicadellidae) in vineyards. Environ. Entomol. 1996, 25, 1224–1234. [Google Scholar] [CrossRef]
- Costello, M.J. Regulated deficit irrigation and density of Erythroneura spp. (Hemiptera: Cicadellidae) on grape. J. Econ. Entomol. 2008, 101, 1287–1294. [Google Scholar] [PubMed] [Green Version]
- Dmitriev, D.A. 3I Interactive Keys and Taxonomic Databases. Auchenorrhyncha Database Search. Available online: http://dmitriev.speciesfile.org (accessed on 19 November 2018).
- Robinson, W. The genus Erythroneura north of Mexico (Homoptera, Cicadellidae). Sci. Bull. Univ. Kans. 1926, 16, 101–155. [Google Scholar] [CrossRef]
- Beamer, R.H. The Erythroneura of the vulnerata group (Homoptera—Cicadellidae). J. Kans. Entomol. Soc. 1946, 19, 15–22. [Google Scholar]
- Martinson, T.E.; Dennehy, T.J. Varietal preferences of Erythroneura leafhoppers (Homoptera: Cicadellidae) feeding on grapes in New York. Environ. Entomol. 1995, 24, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Paxton, D.W.; Thorvilson, H.G. Oviposition of three Erythroneura species on grape leaves in Western Texas. Southwest. Entomol. 1996, 21, 141–144. [Google Scholar]
- Zimmerman, R.; Kondratieff, B.; Nelson, E.; Sclar, C. The life history of two species of grape leafhoppers on wine grapes in western Colorado. J. Kans. Entomol. Soc. 1996, 69, 337–345. [Google Scholar]
- Duso, C.; Bressan, A.; Mazzon, L.; Girolami, V. First record of the grape leafhopper Erythroneura vulnerata Fitch (Hom., Cicadellidae) in Europe. J. Appl. Entomol. 2005, 129, 170–172. [Google Scholar] [CrossRef]
- Girolami, V.; Duso, C.; Mazzon, L.; Bressan, A. Nuova cicalina della vite in Italia. Inf. Agric. 2006, 62, 65–67. [Google Scholar]
- Duso, C.; Moret, R.; Marchegiani, G.; Pozzebon, A. Notes on the distribution and the phenology of Erasmoneura vulnerata (Fitch) (Homoptera: Cicadellidae) in North-eastern Italy. IOBC/WPRS Bull. 2008, 36, 251–254. [Google Scholar]
- Duso, C.; Moret, R.; Manera, A.; Berto, D.; Fornasiero, D.; Marchegiani, G.; Pozzebon, A. Investigations on the grape leafhopper Erasmoneura vulnerata in North-eastern Italy. Insects 2019, 10, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seljak, G. First record of the Nearctic leafhopper Erasmoneura vulnerata (Fitch, 1851) (Hemiptera, Cicadomorpha: Cicadellidae) in Slovenia. Acta Entomol. Slov. 2011, 19, 37–42. [Google Scholar]
- Duso, C.; Borgo, M.; Pozzebon, A.; Mazzon, L.; Mori, N.; Pavan, F.; Fornasiero, D.; Marchesini, E.; Martinez-Sañudo, I.; Zanettin, G. Vite: Erasmoneura vulnerata, una minaccia da valutare. Inf. Agric. 2017, 44, 31–35. [Google Scholar]
- Rizzoli, A.; Battelli, R.; Conedera, M.; Jermini, M. First record of Erasmoneura vulnerata Fitch, 1851 (Hemiptera, Cicadellidae, Typhlocybinae) in Switzerland. Alp. Entomol. 2020, 4, 151. [Google Scholar]
- Šćiban, M.; Kosovac, A. New records and updates on alien Auchenorrhyncha species in Serbia. Pesticidi i Fitomedicina 2020, 35, 9–16. [Google Scholar]
- Zanolli, P.; Pavan, F. Autumnal emergence of Anagrus wasps, egg parasitoids of Empoasca vitis, from grapevine leaves and their migration towards brambles. Agric. For. Entomol. 2011, 13, 423–433. [Google Scholar] [CrossRef]
- Vidano, C. Le cicaline italiane della vite. Hemiptera Typhlocibinae. Boll. Zool. Agr. Bachic. 1958, 1, 61–115. [Google Scholar]
- Patwary, M.U.; Kenchington, E.L.; Bird, C.J.; Zouros, E. The use of random amplified polymorphic DNA markers in genetic studies of the sea scallop Placopecten magellanicus (Gmelin, 1791). J. Shellfish Res. 1994, 13, 547–553. [Google Scholar]
- Martinez-Sañudo, I.; Mazzon, L.; Simonato, M.; Avtzis, D.; Pujade-Villar, J.; Faccoli, M. Tracking the origin and dispersal of the Asian chestnut gall wasp Dryocosmus kuriphilus Yasumatsu (Hymenoptera, Cynipidae) in Europe with molecular markers. Bull. Entomol. Res. 2019, 109, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Viggiani, G. On the misidentification of Anagrus ustulatus Haliday (Hymenoptera: Mymaridae). Zootaxa 2014, 3826, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzon, L.; Martinez-Sañudo, I.; (DAFNAE University of Padova, Legnaro, Italy). Personal communication, 2019.
- Olivier, C.; Vincent, C.; Saguez, J.; Galka, B.; Weintraub, P.G.; Maixner, M. Leafhoppers and planthoppers: Their bionomics, pathogen transmission and management in vineyards. In Arthropod Management in Vineyards; Springer: Dordrecht, The Netherlands, 2012; pp. 253–270. [Google Scholar]
- Triapitsyn, S.V.; Rugman-Jones, P.F.; Jeong, G.; Morse, J.G.; Stouthamer, R. Morphological and molecular differentiation of the Anagrus epos species complex (Hymenoptera: Mymaridae), egg parasitoids of leafhoppers (Hemiptera: Cicadellidae) in North America. Zootaxa 2010, 2428, 1–21. [Google Scholar] [CrossRef]
- Zanolli, P.; Martini, M.; Mazzon, L.; Pavan, F. Morphological and Molecular identification of Anagrus ‘atomus’ Group (Hymenoptera: Mymaridae) individuals from different geographic areas and plant hosts in Europe. J. Insect Sci. 2016, 16, 38. [Google Scholar] [CrossRef] [Green Version]
- Ponti, L.; Ricci, C.; Veronesi, F.; Torricelli, R. Natural hedges as an element of functional biodiversity in agroecosystems: The case of a Central Italy vineyard. Bull. Insectol. 2005, 58, 19–23. [Google Scholar]
- Tacoli, F.; Pavan, F.; Cargnus, E.; Tilatti, E.; Pozzebon, A.; Zandigiacomo, P. Efficacy and mode of action of kaolin in the control of Empoasca vitis and Zygina rhamni (Hemiptera: Cicadellidae) in vineyards. J. Econ. Entomol. 2017, 110, 1164–1178. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Vineyard | Management | Locality | Cultivar | Insecticides Used (n° Applicat.) | Colonization Patterns | Phenology |
|---|---|---|---|---|---|---|---|
| 2017 | AO | Organic | Alonte | Merlot, Glera, | Pyrethrins (1) | ● | ● |
| 45°22′0″ N, 11°25′41″ E | Pinot gris, Incrocio Manzoni 6013 | ||||||
| 2017 | LO1 | Organic | Lonigo | Merlot, Trebbiano T. | Insecticide-free | ● | |
| 45°23′18″ N, 11°23′24″ E | |||||||
| 2017 | AC | Conventional | Alonte | Cabernet S., Pinot gris, Glera | Insecticide-free | ● | ● |
| 45°21′33.6″ N, 11°26′11.3″ E | |||||||
| Lonigo | Garganega, | Insecticide-free | ● | ||||
| 2017 | LC | Conventional | 45°24′16.2″ N 11°22′32.2″ E | Chardonnay, | |||
| Merlot | |||||||
| 2018 | AO | Organic | Alonte | Merlot, Glera | Insecticide-free | ● | |
| 45°22′38.6″ N 11°24′30.6″ E | |||||||
| 2018 | LO2 | Organic | Lonigo | Trebbiano T., Cabernet S. | Insecticide-free | ● | ● |
| 45°22′41.2″ N 11°24′34.4″ E | |||||||
| 2018 | MO | Organic | Monteforte | Garganega | Mineral oil (1) | ● | |
| 45°25′5” N, 11°17′4″ E | |||||||
| 2018 | LC | Conventional | Lonigo | Garganega | Insecticide-free | ● | |
| 2018 | MC | Conventional | Monteforte | Trebbiano S. | Insecticide-free | ● | |
| 45°27′16.6″ N, 11°16′59.2″ E | |||||||
| 2019 | LO1 | Organic | Lonigo | Merlot | Pyrethrins (2) | ● | ● |
| 2019 | MO | Organic | Monteforte | Garganega | Insecticide-free | ● | ● |
| 2019 | AC | Conventional | Alonte | Cabernet S., Glera | Insecticide-free | ● | |
| 2019 | GC | Conventional | Gambellara | Garganega | Insecticide-free | ● | |
| 45°24′17.9″ N 11°22′31.8″ E |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duso, C.; Zanettin, G.; Gherardo, P.; Pasqualotto, G.; Raniero, D.; Rossetto, F.; Tirello, P.; Pozzebon, A. Colonization Patterns, Phenology and Seasonal Abundance of the Nearctic Leafhopper Erasmoneura vulnerata (Fitch), a New Pest in European Vineyards. Insects 2020, 11, 731. https://doi.org/10.3390/insects11110731
Duso C, Zanettin G, Gherardo P, Pasqualotto G, Raniero D, Rossetto F, Tirello P, Pozzebon A. Colonization Patterns, Phenology and Seasonal Abundance of the Nearctic Leafhopper Erasmoneura vulnerata (Fitch), a New Pest in European Vineyards. Insects. 2020; 11(11):731. https://doi.org/10.3390/insects11110731
Chicago/Turabian StyleDuso, Carlo, Giulia Zanettin, Pamela Gherardo, Giulia Pasqualotto, Damiano Raniero, Filippo Rossetto, Paola Tirello, and Alberto Pozzebon. 2020. "Colonization Patterns, Phenology and Seasonal Abundance of the Nearctic Leafhopper Erasmoneura vulnerata (Fitch), a New Pest in European Vineyards" Insects 11, no. 11: 731. https://doi.org/10.3390/insects11110731
APA StyleDuso, C., Zanettin, G., Gherardo, P., Pasqualotto, G., Raniero, D., Rossetto, F., Tirello, P., & Pozzebon, A. (2020). Colonization Patterns, Phenology and Seasonal Abundance of the Nearctic Leafhopper Erasmoneura vulnerata (Fitch), a New Pest in European Vineyards. Insects, 11(11), 731. https://doi.org/10.3390/insects11110731