Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina

 ,
, 

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sites, Traps and Sampling Protocol
2.2. Data Analyses: Effectiveness in Trapping V. velutina
2.3. Data Analyses: Bycatch of Insects and Differences between Trap-Bait Combinations
2.4. Data Analyses: Selectiveness of the Traps
3. Results
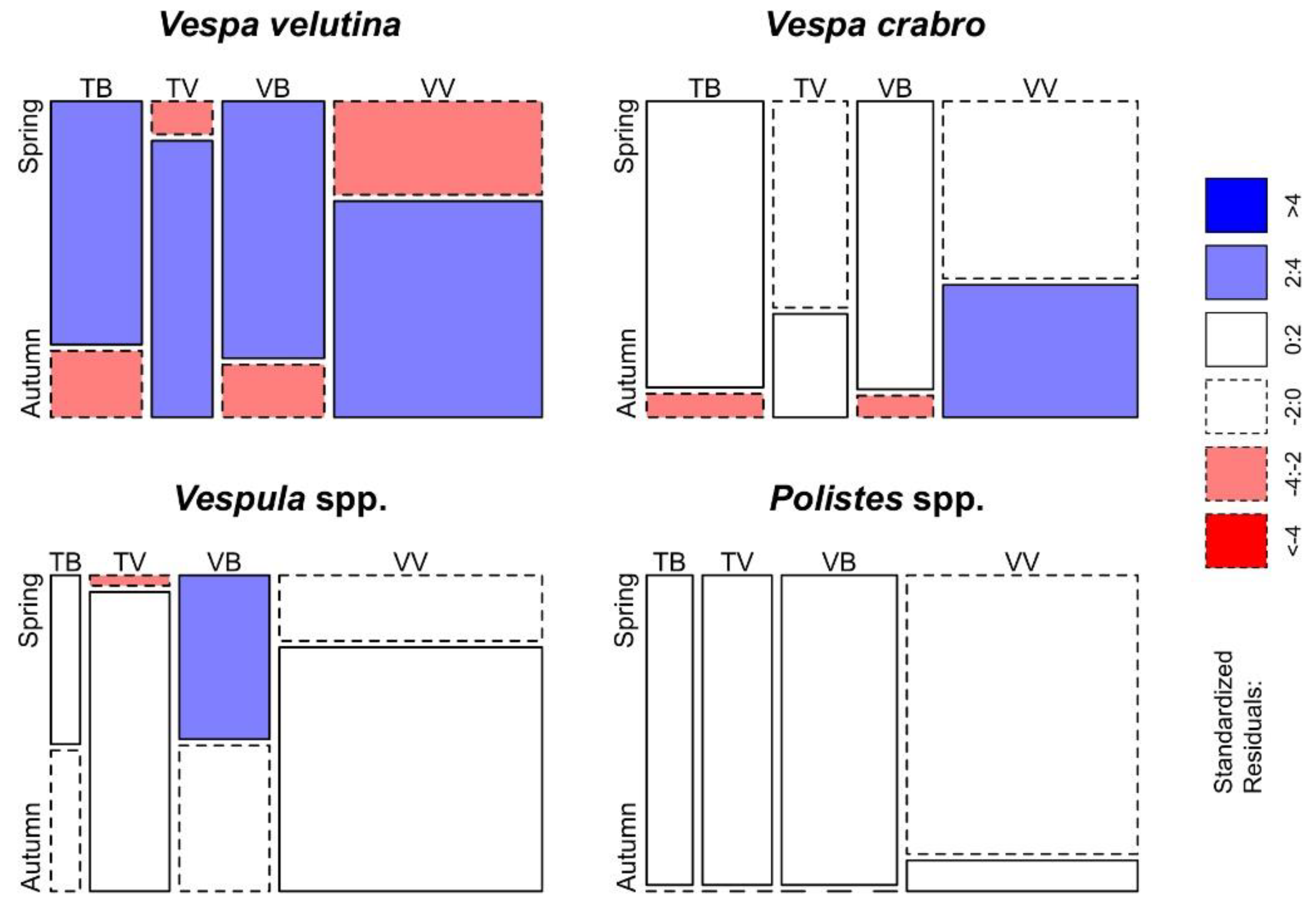
3.1. Effectiveness in Trapping V. velutina
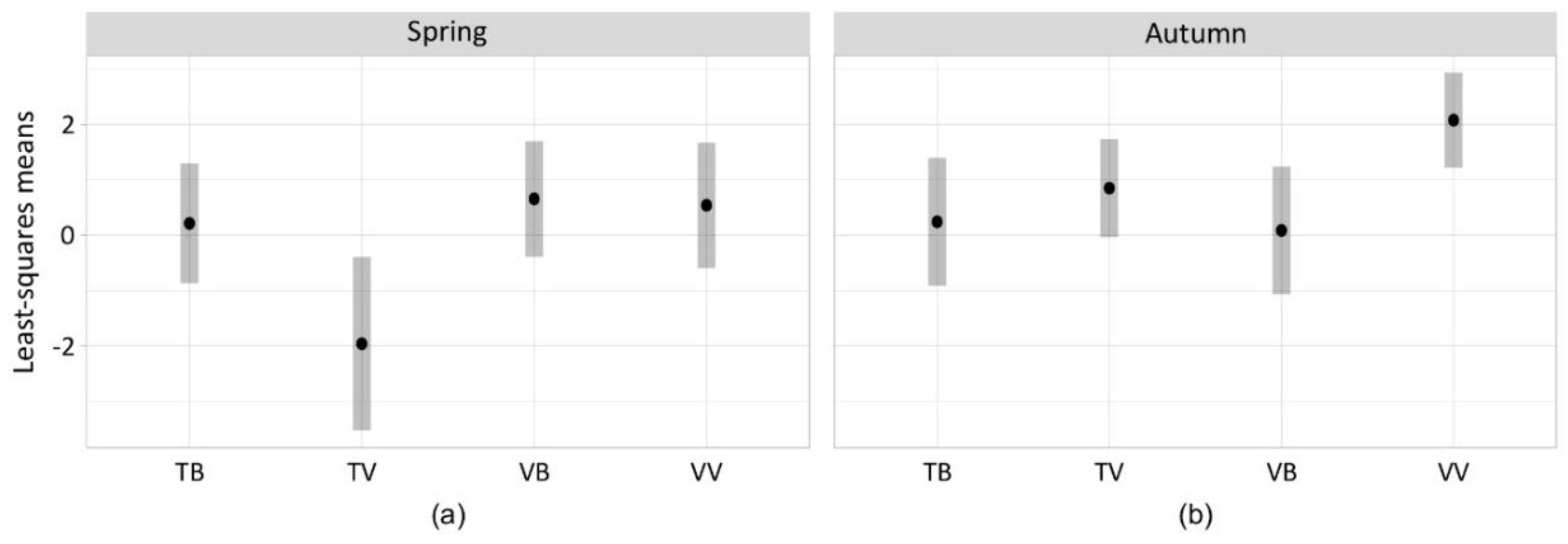
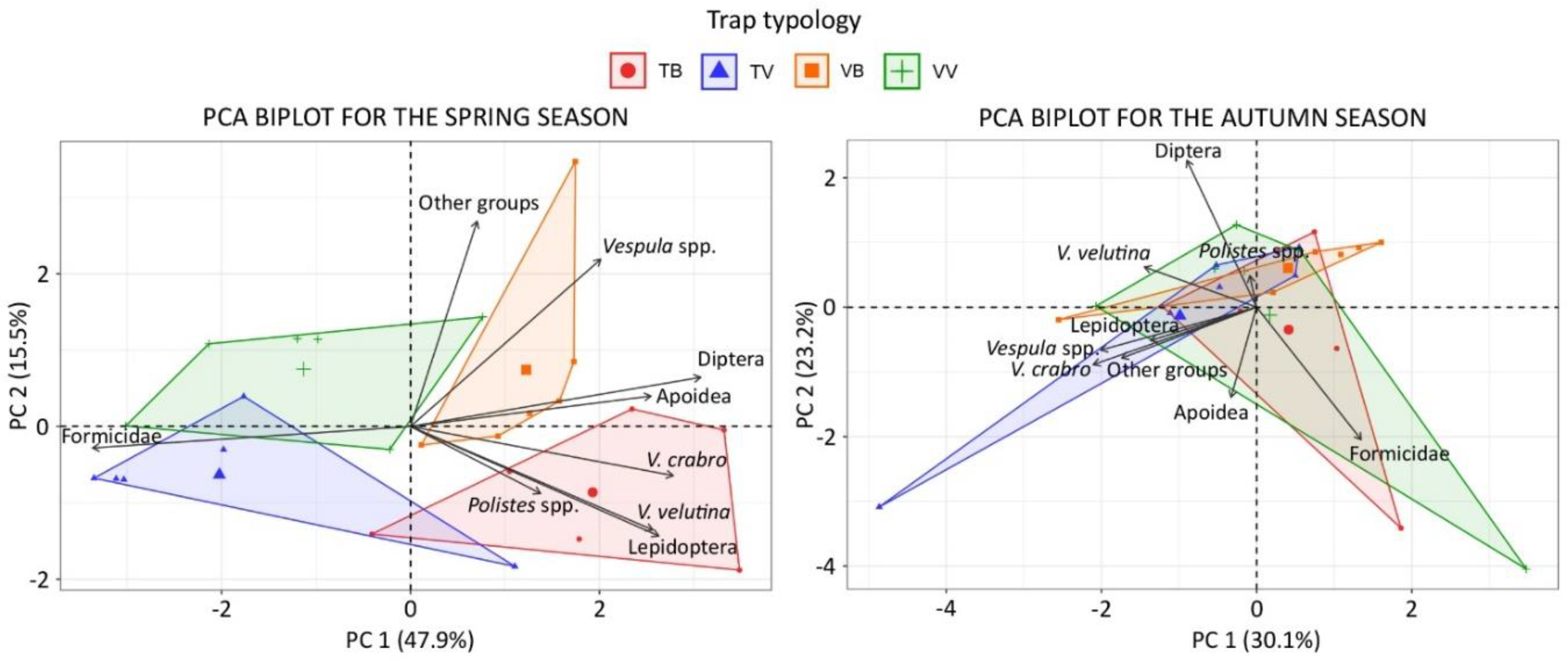
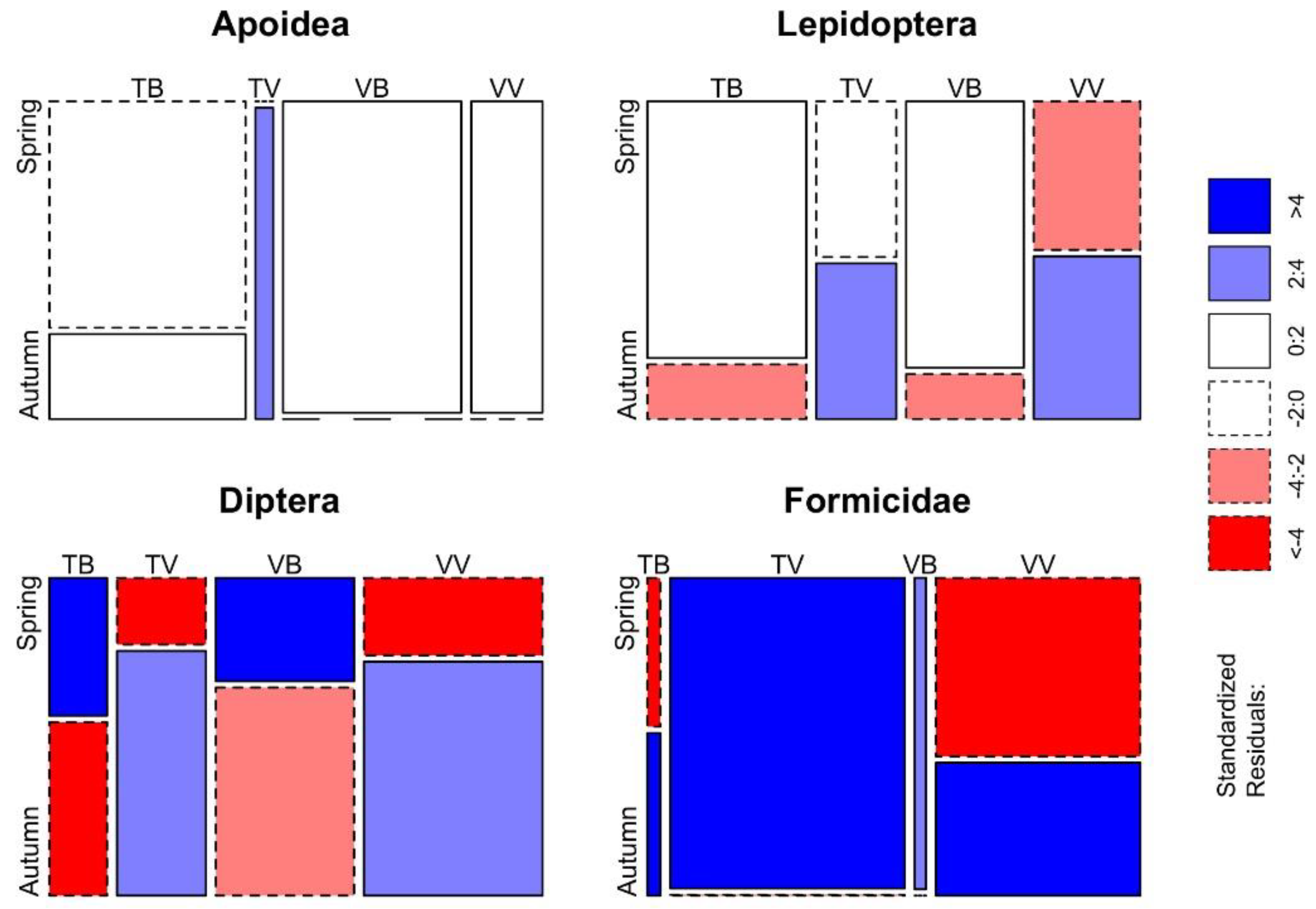
3.2. Bycatch of Insects and Differences between Trap-Bait Combinations
3.3. Selectiveness of the Traps
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Monceau, K.; Bonnard, O.; Thiéry, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Choi, M.B.; Martin, S.J.; Lee, J.W. Distribution, spread, and impact of the invasive hornet Vespa velutina in South Korea. J. Asia-Pac. Entomol. 2012, 15, 473–477. [Google Scholar] [CrossRef]
- Takahashi, J.; Okuyama, H.; Kiyoshi, T.; Takeuchi, T.; Martin, S.J. Origins of Vespa velutina hornets that recently invaded Iki Island, Japan and Jersey Island, UK. Mitochondrial DNA Part A 2019, 30, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An Alien Driver of Honey Bee Colony Losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef]
- Villemant, C.; Muller, F.; Haubois, S.; Perrard, A.; Darrouzet, E.; Rome, Q. Bilan des travaux (MNHN et IRBI) sur l’invasion en france de Vespa velutina, le frelon asiatique prédateur d’abeilles. In Proceedings of the Journée Scientifique Apicole JSA, Arles, France, 11 February 2011; pp. 3–12. [Google Scholar]
- Monceau, K.; Bonnard, O.; Moreau, J.; Thiéry, D. Spatial distribution of Vespa velutina individuals hunting at domestic honeybee hives: Heterogeneity at a local scale. Insect Sci. 2014, 21, 765–774. [Google Scholar] [CrossRef]
- Requier, F.; Rome, Q.; Chiron, G.; Decante, D.; Marion, S.; Menard, M.; Muller, F.; Villemant, C.; Henry, M. Predation of the invasive Asian hornet affects foraging activity and survival probability of honey bees in Western Europe. J. Pest Sci. 2019, 92, 567–578. [Google Scholar] [CrossRef]
- Arca, M.; Papachristoforou, A.; Mougel, F.; Rortais, A.; Monceau, K.; Bonnard, O.; Tardy, P.; Thiéry, D.; Silvain, J.-F.; Arnold, G. Defensive behaviour of Apis mellifera against Vespa velutina in France: Testing whether European honeybees can develop an effective collective defence against a new predator. Behav. Process. 2014, 106, 122–129. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Calviño-Cancela, M. The invasive hornet Vespa velutina affects pollination of a wild plant through changes in abundance and behaviour of floral visitors. Biol. Invasions 2020, 22, 2609–2618. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Salles, J.-M.; Courchamp, F. The economic cost of control of the invasive yellow-legged Asian hornet. NeoBiota 2020, 55, 11–25. [Google Scholar] [CrossRef]
- Budge, G.E.; Hodgetts, J.; Jones, E.P.; Ostojá-Starzewski, J.C.; Hall, J.; Tomkies, V.; Semmence, N.; Brown, M.; Wakefield, M.; Stainton, K. The invasion, provenance and diversity of Vespa velutina Lepeletier (Hymenoptera: Vespidae) in Great Britain. PLoS ONE 2017, 12, e0185172. [Google Scholar] [CrossRef]
- Leza, M.; Miranda, M.Á.; Colomar, V. First detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A challenging study case. Biol. Invasions 2018, 20, 1643–1649. [Google Scholar] [CrossRef]
- Lioy, S.; Manino, A.; Porporato, M.; Laurino, D.; Romano, A.; Capello, M.; Bertolino, S. Establishing surveillance areas for tackling the invasion of Vespa velutina in outbreaks and over the border of its expanding range. NeoBiota 2019, 46, 51–69. [Google Scholar] [CrossRef]
- Demichelis, S.; Manino, A.; Minuto, G.; Mariotti, M.; Porporato, M. Social wasp trapping in north west Italy: Comparison of different bait-traps and first detection of Vespa velutina. Bull. Insectol. 2014, 67, 307–317. [Google Scholar]
- Porporato, M.; Manino, A.; Laurino, D.; Demichelis, S. Vespa velutina Lepeletier (Hymenoptera Vespidae): A first assessment two years after its arrival in Italy. Redia 2014, 97, 189–194. [Google Scholar]
- Turchi, L.; Derijard, B. Options for the biological and physical control of Vespa velutina nigrithorax (Hym.: Vespidae) in Europe: A review. J. Appl. Entomol. 2018, 142, 553–562. [Google Scholar] [CrossRef]
- Sorvari, J. Social wasp (Hymenoptera: Vespidae) beer trapping in Finland 2008–2012: A German surprise. Entomol. Fenn. 2013, 24, 156–164. [Google Scholar] [CrossRef]
- Wegner, G.S.; Jordan, K.K. Comparison of Three Liquid Lures for Trapping Social Wasps (Hymenoptera: Vespidae). J. Econ. Entomol. 2005, 98, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Monceau, K.; Bonnard, O.; Thiéry, D. Chasing the queens of the alien predator of honeybees: A water drop in the invasiveness ocean. Open J. Ecol. 2012, 2, 183–191. [Google Scholar] [CrossRef]
- Bacandritsos, N.; Papanastasiou, I.; Saitanis, C.; Roinioti, E. Three non-toxic insect traps useful in trapping wasps enemies of honey bees. Bull. Insectol. 2006, 59, 135–145. [Google Scholar]
- Rodríguez-Flores, M.S.; Seijo-Rodríguez, A.; Escuredo, O.; del Seijo-Coello, M.C. Spreading of Vespa velutina in northwestern Spain: Influence of elevation and meteorological factors and effect of bait trapping on target and non-target living organisms. J. Pest Sci. 2019, 92, 557–565. [Google Scholar] [CrossRef]
- Monceau, K.; Thiéry, D. Vespa velutina nest distribution at a local scale: An 8-year survey of the invasive honeybee predator. Insect Sci. 2017, 24, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Nossa, S.V.; Novoa, N.; Serrano, A.; Calviño-Cancela, M. Performance of baited traps used as control tools for the invasive hornet Vespa velutina and their impact on non-target insects. Apidologie 2018, 49, 872–885. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org (accessed on 15 May 2020).
- Rome, Q.; Muller, F.J.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste differentiation and seasonal changes in Vespa velutina (Hym.: Vespidae) colonies in its introduced range. J. Appl. Entomol. 2015, 139, 771–782. [Google Scholar] [CrossRef]
- Vick, K.W.; Mankin, R.W.; Cogburn, R.R.; Mullen, M.; Throne, J.E.; Wright, V.F.; Cline, L.D. Review of Pheromone-Baited Sticky Traps for Detection of Stored-Product Insects. J. Kans. Entomol. Soc. 2020, 63, 526–532. [Google Scholar]
- Welzel, K.F.; Choe, D.-H. Development of a Pheromone-Assisted Baiting Technique for Argentine Ants (Hymenoptera: Formicidae). J. Econ. Entomol. 2016, 109, 1303–1309. [Google Scholar] [CrossRef]
- Short, B.D.; Khrimian, A.; Leskey, T.C. Pheromone-based decision support tools for management of Halyomorpha halys in apple orchards: Development of a trap-based treatment threshold. J. Pest Sci. 2017, 90, 1191–1204. [Google Scholar] [CrossRef]
- Wen, P.; Cheng, Y.-N.; Dong, S.-H.; Wang, Z.-W.; Tan, K.; Nieh, J.C. The sex pheromone of a globally invasive honey bee predator, the Asian eusocial hornet, Vespa velutina. Sci. Rep. 2017, 7, 12956. [Google Scholar] [CrossRef]
- Couto, A.; Monceau, K.; Bonnard, O.; Thiéry, D.; Sandoz, J.-C. Olfactory Attraction of the Hornet Vespa velutina to Honeybee Colony Odors and Pheromones. PLoS ONE 2014, 9, e115943. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |







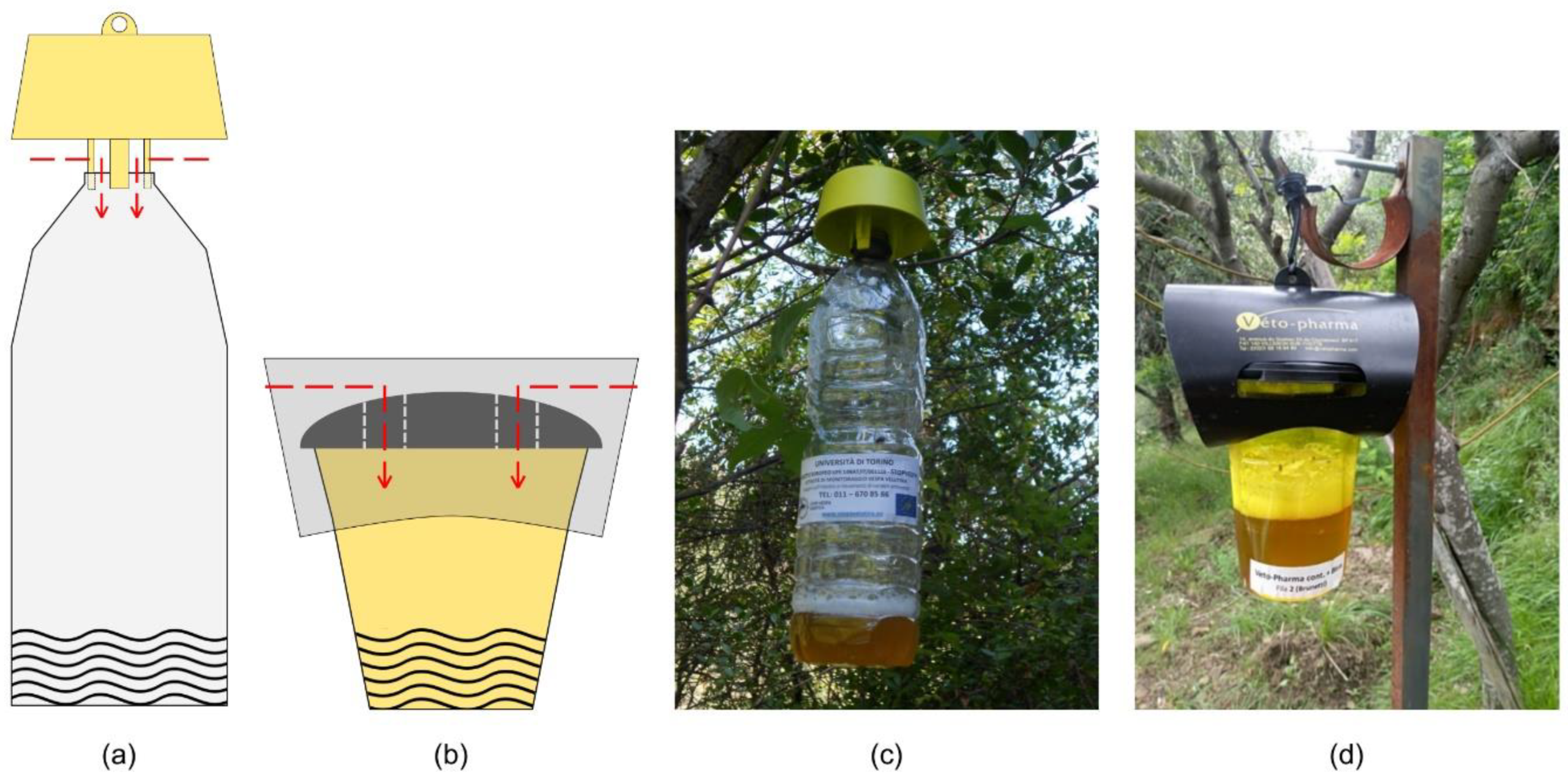{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TOTAL | SPRING | AUTUMN | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TB | TV | VB | VV | TB | TV | VB | VV | ||||||||||||
| N. of Individuals | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | Mean per Trap (SD) | % of Insects | |
| Hymenoptera | |||||||||||||||||||
| Vespoidea | |||||||||||||||||||
| Vespa velutina | 213 | 1.48 (2.64) | 1.02 | 1.38 (1.78) | 3.65 | 0.13 (0.33) | 0.06 | 1.63 (2.36) | 2.72 | 1.21 (2.36) | 0.80 | 0.75 (1.09) | 1.02 | 2.08 (1.71) | 1.53 | 0.67 (1.11) | 0.39 | 5.58 (5.22) | 1.35 |
| Vespa crabro | 261 | 1.81 (3.16) | 1.25 | 2.54 (2.74) | 6.74 | 1.17 (2.36) | 0.52 | 1.67 (2.25) | 2.79 | 2.63 (4.09) | 1.74 | 0.42 (0.95) | 0.57 | 1.17 (1.99) | 0.86 | 0.25 (0.60) | 0.15 | 3.92 (5.72) | 0.95 |
| Vespula spp. | 174 | 1.21 (3.08) | 0.84 | 0.25 (0.52) | 0.66 | 0.04 (0.20) | 0.02 | 0.75 (1.13) | 1.26 | 0.88 (1.92) | 0.58 | 0.42 (0.64) | 0.57 | 2.42 (3.64) | 1.78 | 1.33 (1.84) | 0.79 | 6.50 (7.16) | 1.57 |
| Polistes spp. | 20 | 0.14 (0.40) | 0.10 | 0.08 (0.28) | 0.22 | 0.13 (0.33) | 0.06 | 0.21 (0.41) | 0.35 | 0.38 (0.70) | 0.25 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.08 (0.28) | 0.02 |
| Formicidae | 9641 | 67.0 (258.5) | 46.35 | 5.4 (7.7) | 14.26 | 202.8 (518.8) | 90.95 | 9.9 (14.5) | 16.55 | 101.6 (173.8) | 67.50 | 11.8 (38.7) | 15.99 | 1.2 (3.3) | 0.86 | 0.0 (0.0) | 0.00 | 151.3 (358.7) | 36.63 |
| Hymenoptera Apoidea | |||||||||||||||||||
| Apis mellifera | 18 | 0.13 (0.45) | 0.09 | 0.29 (0.68) | 0.77 | 0.00 (0.00) | 0.00 | 0.33 (0.75) | 0.56 | 0.08 (0.28) | 0.06 | 0.08 (0.28) | 0.11 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 |
| Bombus spp. | 6 | 0.04 (0.20) | 0.03 | 0.04 (0.20) | 0.11 | 0.00 (0.00) | 0.00 | 0.08 (0.28) | 0.14 | 0.08 (0.28) | 0.06 | 0.00 (0.00) | 0.00 | 0.08 (0.28) | 0.06 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 |
| Other Apoidea | 2 | 0.01 (0.12) | 0.01 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.17 (0.37) | 0.23 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 | 0.00 (0.00) | 0.00 |
| Diptera | 9993 | 69.4 (111.3) | 48.04 | 23.0 (16.5) | 60.89 | 17.1 (23.6) | 7.68 | 41.2 (27.1) | 68.99 | 39.9 (45.3) | 26.52 | 57.8 (56.7) | 78.56 | 125.7 (117.2) | 92.52 | 165.9 (170.0) | 97.93 | 241.1 (200.8) | 58.39 |
| Lepidoptera | 382 | 2.65 (3.97) | 1.84 | 4.50 (5.09) | 11.93 | 1.38 (2.29) | 0.62 | 3.46 (4.68) | 5.80 | 1.75 (2.74) | 1.16 | 1.92 (2.36) | 2.61 | 2.75 (3.39) | 2.02 | 1.17 (1.28) | 0.69 | 3.83 (5.62) | 0.93 |
| Other groups | 90 | 0.63 (1.49) | 0.43 | 0.29 (0.54) | 0.77 | 0.21 (0.41) | 0.09 | 0.50 (1.15) | 0.84 | 2.00 (2.86) | 1.33 | 0.25 (0.43) | 0.34 | 0.50 (0.87) | 0.37 | 0.08 (0.28) | 0.05 | 0.67 (0.94) | 0.16 |
| Variables | β | SE | Z | p |
|---|---|---|---|---|
| Season (Autumn) | 0.03 | 0.71 | 0.04 | 0.968 |
| Bait (VespaCatch) | −2.18 | 0.63 | −3.44 | <0.001 |
| Trap_model (VespaCatch) | 0.44 | 0.26 | 1.71 | 0.088 |
| Season (Autumn): Bait (VespaCatch) | 2.78 | 0.79 | 3.52 | <0.001 |
| Season (Autumn): Trap_model (VespaCatch) | −0.60 | 0.64 | −0.94 | 0.348 |
| Bait (VespaCatch): Trap typology (VespaCatch) | 2.06 | 0.69 | 2.97 | <0.01 |
| Season (Autumn): Bait (VespaCatch): Trap typology (VespaCatch) | −0.68 | 0.95 | −0.71 | 0.475 |
| SPRING | AUTUMN | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | TB | TV | VB | VV | p | TB | TV | VB | VV | |||||||||
| Apoidea | 0.763 | 0.24 | a | 0.00 | a | 0.26 | a | 0.14 | a | <0.01 | 0.33 | b | 0.04 | ab | 0.00 | ab | 0.00 | a * |
| Vespa crabro | <0.001 | 1.85 | a * | 9.33 | b | 1.03 | a * | 2.17 | ab | 0.798 | 0.56 | a | 0.56 | a | 0.38 | a | 0.70 | a |
| Vespula spp. | 0.047 | 0.18 | a | 0.33 | a | 0.46 | a | 0.72 | a | 0.339 | 0.56 | a | 1.16 | a | 2.00 | a | 1.16 | a |
| Polistes spp. | 0.017 | 0.06 | a | 1.00 | a | 0.13 | a | 0.31 | a | 1.000 | 0.00 | a | 0.00 | a | 0.00 | a | 0.01 | a |
| Formicidae | <0.001 | 3.91 | a * | 1622 | c | 6.08 | a * | 84.10 | b | <0.001 | 15.67 | b | 0.56 | a * | 0.00 | a * | 27.09 | b |
| Diptera | <0.001 | 16.7 | a * | 137 | b | 25.33 | a * | 33.03 | ab | <0.001 | 77.00 | ab | 60.32 | a * | 248 | b | 43.18 | a * |
| Lepidoptera | <0.01 | 3.27 | ab | 11.00 | b | 2.13 | a * | 1.45 | a * | <0.05 | 2.56 | b | 1.32 | ab | 1.75 | ab | 0.69 | a * |
| Other groups | <0.001 | 0.21 | a * | 1.67 | ab | 0.31 | a * | 1.66 | b | 0.365 | 0.33 | a | 0.24 | a | 0.13 | a | 0.12 | a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina. Insects 2020, 11, 706. https://doi.org/10.3390/insects11100706
Lioy S, Laurino D, Capello M, Romano A, Manino A, Porporato M. Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina. Insects. 2020; 11(10):706. https://doi.org/10.3390/insects11100706
Chicago/Turabian StyleLioy, Simone, Daniela Laurino, Michela Capello, Andrea Romano, Aulo Manino, and Marco Porporato. 2020. "Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina" Insects 11, no. 10: 706. https://doi.org/10.3390/insects11100706
APA StyleLioy, S., Laurino, D., Capello, M., Romano, A., Manino, A., & Porporato, M. (2020). Effectiveness and Selectiveness of Traps and Baits for Catching the Invasive Hornet Vespa velutina. Insects, 11(10), 706. https://doi.org/10.3390/insects11100706