The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate

 , , ,
, , ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
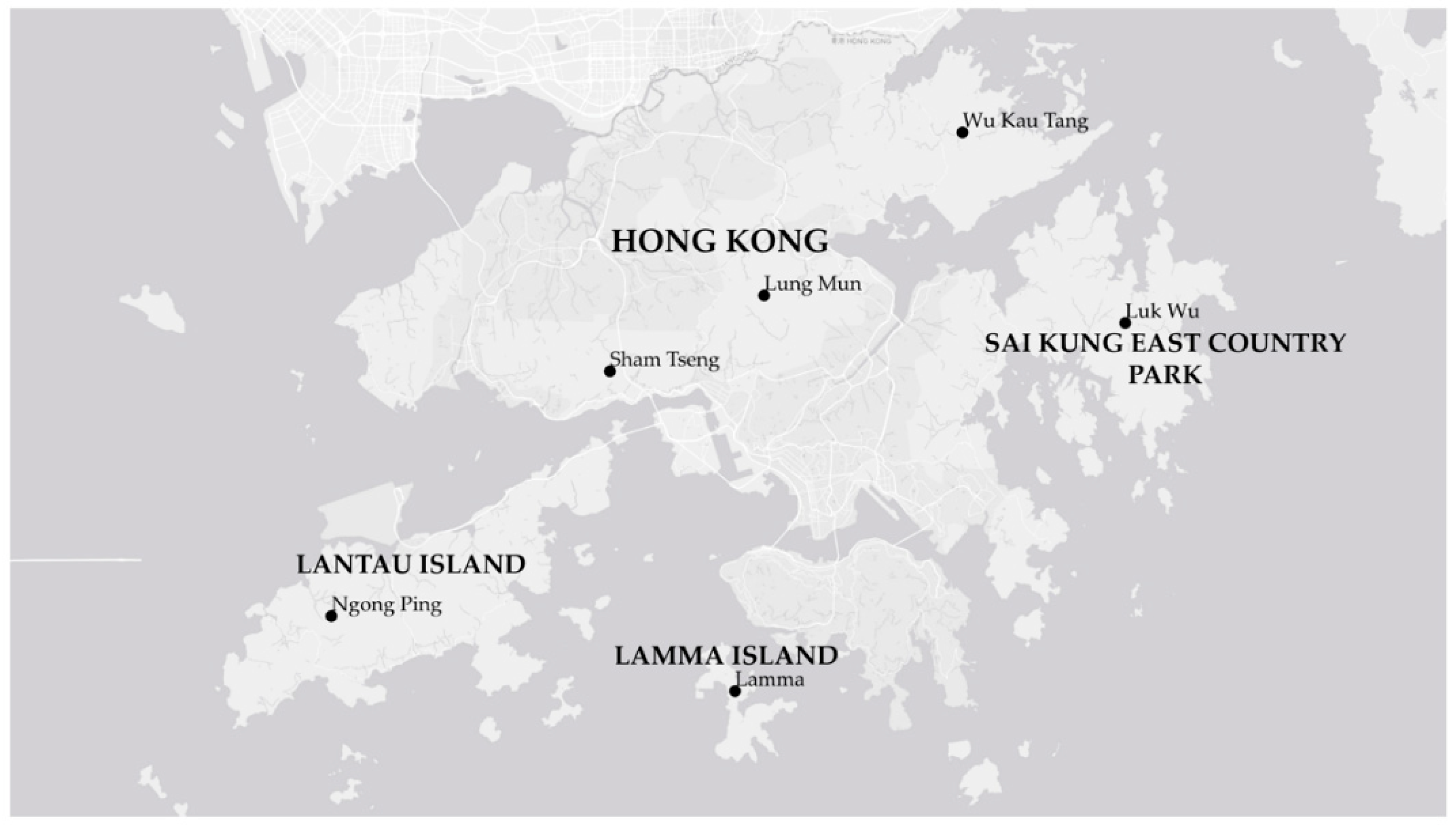
2.1. Collection
2.2. Sample Preparation for Descriptions
2.3. Rearing
2.4. Preliminary Host Range Examination
2.5. Tracking Larval Size and Development
3. Results
3.1. Species Description
3.1.1. Description of Adult Female
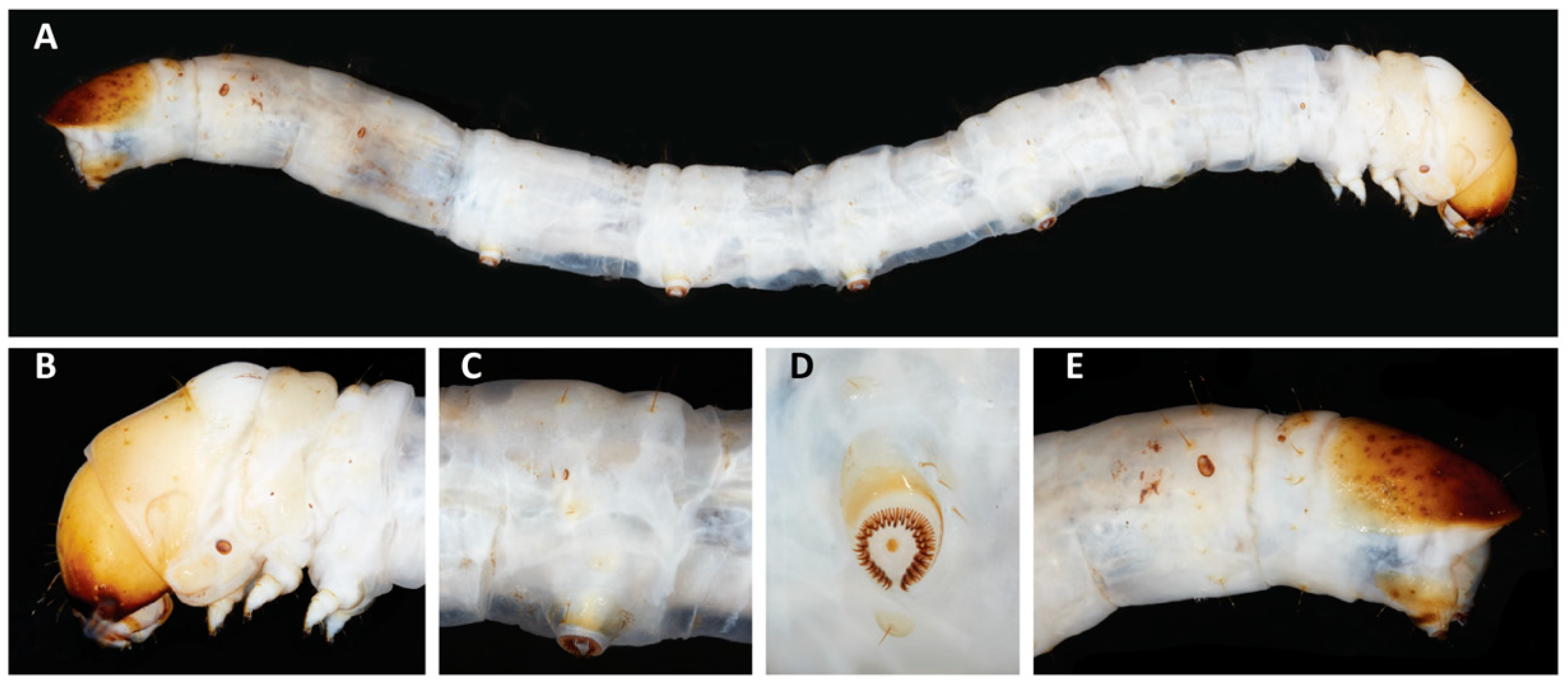
3.1.2. Larval Diagnosis
3.1.3. Pupa
3.1.4. Egg
3.1.5. Specimens Examined
3.2. Casmara Subagronoma Development and Initial Host Range Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Langeland, K.A.; Craddock Burks, K.C. Identification and Biology of Non-Native Plants in Florida’s Native Areas; University of Florida Institute of Food and Agriculture Sciences: Gainesville, FL, USA, 1998. [Google Scholar]
- University of Georgia EddMaps-Early Detection and Distribution Mapping System. Available online: www.eddmaps.org (accessed on 19 August 2020).
- Gordon, D.R.; Thomas, K.P. Florida’s invasion by nonindigenous plants: History, screening, and regulation. In Strangers in Paradise: Impact and Management of Nonindigenous Species in Florida; Simberloff, D., Schmitz, D., Brown, T.C., Eds.; Island Press: Washington, DC, USA, 1997; pp. 21–37. [Google Scholar]
- Kusuma, I.W.; Ainiyati, N.; Suwinarti, W. Search for biological activities from an invasive shrub species rosemyrtle (Rhodomyrtus tomentosa). Nusant. Biosci. 2016, 8. [Google Scholar] [CrossRef]
- Hosaka, E.Y.; Thistle, A. Noxious plants of the Hawaiian ranges (Bulletin 52). In USDA Extension; University of Hawaii: Manoa, HI, USA, 1954. [Google Scholar]
- Krauss, N.L.H. Biological control investigations on some Hawaiian weeds. Proc. Hawaii. Entomol. Soc. 1966, 19, 223–231. [Google Scholar]
- Winotai, A.; Wright, T.; Goolsby, J.A. Herbivores in Thailand on Rhodomyrtus tomentosa (Myrtaceae), an invasive weed in Florida. Fla. Entomol. 2005, 88, 104–105. [Google Scholar] [CrossRef]
- Adamski, D.; Makinson, J.R.; Brown, B.T.; Wright, S.A.; Prattand, P.D.; Brown, J.W. Description and evaluation of Metharmostis multilineata (Cosmopterigidae) and Idiophantis soreuta (Gelechiidae) (Lepidoptera: Gelechioidea) for biocontrol of downy rose myrtle, Rhodomyrtus tomentosa (Myrtaceae). J. Lepid. Soc. 2013, 67, 111–127. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Mattison, E.D.; Metz, M.A.; Pratt, P.D.; Purcell, M.F. Host range of the defoliator Strepsicrates sp. is too broad for biological control of the invasive weed Rhodomyrtus tomentosa. Biocontrol Sci. Technol. 2016, 26, 861–865. [Google Scholar] [CrossRef]
- Pratt, P.D.; Makinson, J.; Purcell, M.; Pogue, M.G.; Rayamajhi, M.B.; Center, T.D. Rhodomyrtus tomentosa (Myrtales: Myrtaceae): New host records for Metanastria gemella (Lepidoptera: Lasiocampidae) and Arna bipunctapex (Lepidoptera: Lymantriidae). Flen 2013, 96, 641–642. [Google Scholar] [CrossRef]
- Pratt, P.D.; Wright, S.A.; Makinson, J.R.; Purcell, M.F.; Center, T.D.; Rayamajhi, M.B. The broad host range of Metharmostis multilineata precludes its use as a biological control agent of Rhodomyrtus tomentosa. Biocontrol Sci. Technol. 2014, 24, 1188–1191. [Google Scholar] [CrossRef]
- Pratt, P.D.; Herdocia, K.; Valentin, V.; Makinson, J.; Purcell, M.F.; Mattison, E.; Rayamajhi, M.B.; Moran, P.; Raghu, S. Development rate, consumption, and host fidelity of Neostauropus alternus (Walker, 1855) (Lepidoptera: Notodontidae). Panp 2016, 92, 200–209. [Google Scholar] [CrossRef]
- Smith, M.C.; Wheeler, G.S.; Barr, K.L.; Brown, B.T.; Makinson, J.R. Non-selective feeding and oviposition behaviour of Cryptocephalus trifasciata (Coleoptera: Chrysomelidae: Cryptocephalinae) precludes its use as a biological control agent for downy rose myrtle (Rhodomyrtus tomentosa). Biocontrol Sci. Technol. 2017, 27, 445–448. [Google Scholar] [CrossRef]
- Caradja, A. New descriptions written by Mr. Edward Meyrick. Acad. Rom. Mem. Sect. Stiint. 1925, 3, 125. [Google Scholar]
- Caradja, A. Second contribution to our knowledge about the Pyralidae and Microlepidoptera of Kwanhsien. Bull. Sect. Sci. Acad. Roum. 1931, 14, 59–75. [Google Scholar]
- Caradja, A.; Meyrick, E. Materialien zu einer microlepidopteren-fauna Kwangtungs. Deutsch. Ent. Zeitschr. Iris 1933, 1, 37. [Google Scholar]
- Chen, J.T. Morphology and habit of six main borers of the tea bush. Acta Entomol. Sin. 1958, 8, 272–280. [Google Scholar]
- Jaros, J.; Spitzer, K.; Havelka, J.; Park, K.T. Synecological and biogeographical outlines of Lepidoptera communities in North Korea. Insecta Koreana 1992, 9, 78–104. [Google Scholar]
- Wang, S.X. Oecophoridae of China (Insecta, Lepidoptera); Science Press: Beijing, China, 2006. [Google Scholar]
- Wang, S.; Kendrick, R.C.; Sterling, P. Microlepidoptera of Hong Kong: Oecophoridae I: The genus Promalactis Meyrick. Zootaxa 2009, 2239, 31–44. [Google Scholar] [CrossRef]
- Byun, B.-K.; Park, Y.M.; Kim, S.; Lee, B.-W.; Bang, N. Notes on Casmara agronoma Meyrick (Lepidoptera, Oecophoridae) in Korea: Casmara agronoma in Korea. Entomol. Res. 2012, 42, 281–283. [Google Scholar] [CrossRef]
- Lvovsky, A.L. New and little known species of the genus Casmara Walker, 1863. Rossica 2013, 22, 107–110. [Google Scholar] [CrossRef]
- Meyrick, E. Descriptions of Indian micro-lepidoptera. J. Bombay Nat. Hist. Soc. 1907, 17, 742. [Google Scholar]
- Meyrick, E. Exotic Microlepidoptera; Biodiversity Heritage Library: London, UK, 1914; Volume 1, p. 237. [Google Scholar]
- Meyrick, E. Exotic Microlepidoptera; Biodiversity Heritage Library: London, UK, 1916; Volume 2, p. 544. [Google Scholar]
- Bradley, J.D. XXIX–A new species of Casmara Walker, 1863, from Malaya—(Lepidoptera: Oecophoridae). Ann. Mag. Nat. Hist. 1953, 6, 319–320. [Google Scholar] [CrossRef]
- Diakonoff, A. Records and descriptions of South Asiatic Microlepidoptera. Tijdschr. Entomol. 1966, 109, 49–81. [Google Scholar]
- Moruati, S. A new and an unrecorded species of Casmara Walker (Lepidoptera: Oecophoridae) from Thailand. Tinea 1985, 12, 11–16. [Google Scholar]
- Robinson, G.S.; Tuck, K.R.; Shaffer, M. A Field Guide to The Smaller Moths of South-East Asia; Natural History Museum of London: London, UK, 1994. [Google Scholar]
- Common, I.F. Oecophorine Genera of Australia III: The Barea Group and Unplaced Genera (Lepidoptera: Oecophoridae); CSIRO Publishing: Clayton, Australia, 2000; ISBN 978-0-643-10585-0. [Google Scholar]
- Kalshoven, L.G.E. The biology of Casmara kalshoveni Diakonhoff, an oecophorid borer. Tijdschr. Entomol. 1966, 109, 87–89. [Google Scholar]
- Li, M.M.; Shu, J.P.; Wang, H.J.; Xu, T.S.; Wang, J.T.; Liu, D.F.; Hua, Z.Y. Biological characteristics of Casmara patrona (Lepidoptera: Oecophoridae). For. Res. 2015, 28, 900–905. [Google Scholar]
- Wang, S.; Zhang, L.; Wang, J. Four new species of the genus Casmara Walker, 1863 (Lepidoptera: Oecophoridae) from China. Zootaxa 2012, 3239, 58–63. [Google Scholar] [CrossRef]
- Xiong, P.C. Optimal top-clipping dates for control of Casmara patrona. Plant Protect. 1987, 13, 29. [Google Scholar]
- Das, G.M. Pests of Tea in North East India and their Control; Tocklaj Experimental Station: Assam, India, 1965. [Google Scholar]
- van Driem, G.L. The Tale of Tea: A Comprehensive History of Tea from Prehistoric Times to the Present Day; BRILL: Leiden, The Netherlands, 2019; ISBN 978-90-04-39360-8. [Google Scholar]
- Zwölfer, H.; Harris, P. Host Specificity determination of insects for biological control of weeds. Ann. Rev. Entomol. 1971, 16, 159–178. [Google Scholar] [CrossRef]
- Wapshere, A.J. A strategy for evaluating the safety of organisms for biological weed control. Ann. Appl. Biol. 1974, 77, 201–211. [Google Scholar] [CrossRef]
- Briese, D. The centrifugal phylogenetic method used to select plants for host-specificity testing for weed biological control agents: Can and should it be modernized? Centrif. Phylogenet. Method Used Select Plants Host-Specif. Test. Weed Biol. Control Agents 2003, 7, 23–33. [Google Scholar]
- Hinz, H.L.; Winston, R.L.; Schwarzländer, M. How safe is weed biological control? a global review of direct nontarget attack. Quart. Rev. Biol. 2019, 94, 1–27. [Google Scholar] [CrossRef]
- Walcott, S.M. Tea production in South Carolina. Southeastern Geogr. 1999, 39, 61–74. [Google Scholar] [CrossRef]
- Charleston Tea Garden. Available online: www.charlestonteagarden.com (accessed on 8 March 2019).
- USDA APHIS. Technical Advisory Group for Biological Control Agents of Weeds Manual, 1st ed.; U.S. Dept. of Agriculture: Washington, DC, USA, 2016.
- Clarke, J.F.G. The preparation of slides of genitalia of Lepidoptera. Bull. Brooklyn Entomol. Soc. 1941, 36, 149–161. [Google Scholar]
- Robinson, G.S. The preparation of slides of Lepidoptera genitalia with special reference to the Microlepidoptera. Entomol. Gaz. 1976, 27, 127–132. [Google Scholar]
- Hodges, R.W. Gelechioidea: Oecophoridae. The Moths of America North of Mexico; E.W. Classey Ltd. & R.B.D. Publications: London, UK, 1974. [Google Scholar]
- Hodges, R.W. The Gelechioidea. In Handbook of Zoology, Arthropoda: Insecta, Part 35, Lepidoptera, Moths and Butterflies; Kristensen, N.P., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; Volume 4, pp. 131–158. [Google Scholar]
- Kristensen, N.P. Skeleton and muscles: Adults. In Handbook of Zoology, Arthropoda: Insecta, Part 35, Lepidoptera, Moths and Butterflies: Morphology, Physiology and Development; Kristensen, N.P., Ed.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 2, pp. 39–131. [Google Scholar]
- Patočka, J.; Turčáni, M. Lepidoptera Pupae. Central European Species. Plate Volume; Apollo Books: Stenstrup, Denmark, 2005. [Google Scholar]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.V.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef]
- Smith, M.C.; Lake, E.C.; Wheeler, G.S. Oviposition choice and larval performance of Neomusotima conspurcatalis on leaflet types of the invasive fern, Lygodium microphyllum. Entomol. Exp. Appl. 2016, 160, 11–17. [Google Scholar] [CrossRef]
- Muirhead, K.A.; Murphy, N.P.; Sallam, N.; Donnellan, S.C.; Austin, A.D. Phylogenetics and genetic diversity of the Cotesia flavipes complex of parasitoid wasps (Hymenoptera: Braconidae), biological control agents of lepidopteran stemborers. Mol. Phylogenet. Evolut. 2012, 63, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, G.R. Biological Control of Alligatorweed, Alternanthera philoxeroides, the world’s first aquatic weed success story. Castanea 1996, 61, 232–243. [Google Scholar]
- Jarvis, P.J.; Fowler, S.V.; Paynter, Q.; Syrett, P. Predicting the economic benefits and costs of introducing new biological control agents for Scotch broom Cytisus scoparius into New Zealand. Biol. Control 2006, 39, 135–146. [Google Scholar] [CrossRef]
- Paynter, Q.; Fowler, S.V.; Gourlay, A.H.; Peterson, P.G.; Smith, L.A.; Winks, C.J. Relative performance on test and target plants in laboratory tests predicts the risk of non-target attack in the field for arthropod weed biocontrol agents. Biol. Control 2015, 80, 133–142. [Google Scholar] [CrossRef]
- Smith, C.S.; Wilson, C.G. Close to the Edge: Microhabitat Selection by Neurostrota gunniella (Busck) (Lepidoptera: Gracillariidae), a Biological Control Agent for Mimosa pigra L. in Australia. Aust. J. Entomol. 1995, 34, 177–180. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Larva lShipment Number * | Test Initiated | Larva ID | Date Individual Observed Dead | Stage at Death | Cause of Death | Last Date Individual Observed Alive † | Stage at Observation | Date Pupa | Date Adult | Sex | Maximum Lifespan (Days) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rhodomyrtus tomentosa | 2015-1008 | 19-Jan-16 | 1 | 1-Jun-16 | larva | parasitoid | 22-Apr-16 | larva | 134 | |||
| 2015-1008 | 19-Jan-16 | 5 | 17-Feb-16 | larva | transfer injury | 17-Feb-16 | larva | 29 | ||||
| 2015-1008 | 19-Jan-16 | 16 | 22-Mar-18 | larva | diminished feeding | 22-Mar-18 | larva | 793 | ||||
| 2015-1008 | 19-Jan-16 | 31 | 5-Apr-18 | larva | trapped in cage fold | 5-Apr-18 | larva | unknown | unknown | male | 807 | |
| 2015-1008 | 19-Jan-16 | 38 | 30-Sep-17 | larva | failed to bore after transfer | 30-Sep-17 | larva | 620 | ||||
| Myrtus communis | 2015-1008 | 19-Jan-16 | 29 | 1-Apr-16 | larva | diminished feeding | 15-Mar-16 | larva | 73 | |||
| 2015-1008 | 19-Jan-16 | 44 | 22-Feb-16 | larva | rejected stem | 9-Feb-16 | larva | 34 | ||||
| 2015-1008 | 19-Jan-16 | 30 | 12-Dec-16 | larva | diminished feeding | 7-Nov-16 | larva | 328 | ||||
| 2015-1008 | 19-Jan-16 | 9 | 14-Apr-18 | larva | bored to soil level and died | 10-May-17 | larva | 816 | ||||
| 2015-1008 | 19-Jan-16 | 8 | 12-Jun-17 | larva | diminished feeding | 10-May-17 | larva | 510 | ||||
| Rhodomyrtus tomentosa | 2016-1001 | 14-Sep-16 | 12 | 8-Dec-16 | larva | parasitized | 15-Nov-16 | larva | 85 | |||
| 2016-1001 | 14-Sep-16 | 9 | 19-Sep-16 | larva | unknown | 12-Sep-16 | larva | 5 | ||||
| 2016-1001 | 14-Sep-16 | 22 | 11-Jun-18 | larva | unknown | 27-Sep-17 | larva | 635 | ||||
| Myrtus communis | 2016-1001 | 14-Sep-16 | 1 | 12-Nov-16 | larva | parasitized | 14-Sep-16 | larva | 59 | |||
| 2016-1001 | 14-Sep-16 | 3 | 16-Dec-16 | larva | parasitized | 27-Oct-16 | larva | 93 | ||||
| 2016-1001 | 14-Sep-16 | 6 | 27-Sep-17 | larva | failed to bore after transfer | 25-Jun-17 | larva | 378 | ||||
| Rhodomyrtus tomentosa | 2015-1008 | 17-Feb-16 | 6 | 29-Jun-16 | larva | parasitoids | 6-Jun-16 | larva | 133 | |||
| 2015-1008 | 16-Feb-16 | 28 | 18-Oct-16 | larva | parasitoids | 27-Sep-16 | larva | 245 | ||||
| 2015-1008 | 17-Feb-16 | 33 | 17-Aug-17 | larva | diminished feeding | 25-Jul-17 | larva | 547 | ||||
| Myrcianthes fragrans | 2015-1008 | 17-Feb-16 | 4 | 26-Jul-16 | adult | 17-Jul-16 | adult | 17-Jun-16 | 17-Jul-16 | male | 160 | |
| 2015-1008 | 16-Feb-16 | 25 | 6-Jun-16 | larva | diminished feeding | 3-Jun-16 | larva | 111 | ||||
| 2015-1008 | 17-Feb-16 | 43 | 6-Jun-17 | pupa | pupal orientation ‡ | 14-Apr-17 | pupa | 475 | ||||
| Rhodomyrtus tomentosa | 2016-1001 | 14-Sep-16 | 8 | 28-Sep-17 | larva | failed to bore after transfer | 7-Jun-17 | larva | 379 | |||
| 2016-1001 | 14-Sep-16 | 18 | 12-Jun-18 | larva | unknown | 6-Jun-17 | larva | 636 | ||||
| 2016-1001 | 14-Sep-16 | 10 | 11-Jun-18 | larva | plant died | 6-Jun-17 | larva | 635 | ||||
| Myrcianthes fragrans | 2016-1001 | 14-Sep-16 | 2 | 27-Sep-17 | larva | failed to bore after transfer | 26-Sep-17 | larva | 378 | |||
| 2016-1001 | 14-Sep-16 | 19 | 5-Aug-17 | adult | adult | 13-Jul-17 | 5-Aug-17 | male | 325 | |||
| 2016-1001 | 14-Sep-16 | 15 | 8-Jun-17 | adult | adult | 11-May-17 | 8-Jun-17 | male | 267 |
| Hong Kong Collection Sites 12 March 2018 | Larva ID | Measurements of Hong Kong Damaged Stems Containing Larvae | Host Plant | Dried Frass Weight (g) After Transfer to Florida Host Material | Larval Lifespan in Laboratory (Weeks) | ||
|---|---|---|---|---|---|---|---|
| Stem Diam (mm) | Larval Bore Diam (mm) | 15 March to 13 April | 14 April to 27 April | ||||
| Lantau | 2 | 3.75 | 1.13 | R. tomentosa | 0.03 | 0.03 | 39 |
| 4 | 2.98 | 1.37 | 0.14 | 0.03 | >24 | ||
| 9 | 3.79 | 1.51 | 0.24 | 0.36 | >24 | ||
| 12 | 3.86 | 1.84 | 0.27 | 0.18 | 23 | ||
| 15 | 4.6 | 2.07 | 0.57 | 0.78 | 30 | ||
| 20 | 3.19 | 1.55 | 0.33 | 0.22 | 39 | ||
| 24 | 3.96 | 1.58 | 0.25 | 0.14 | >24 | ||
| 25 | 3.96 | 1.27 | 0.30 | 0.39 | >24 | ||
| 10 | 3.68 | 1.68 | 0.38 | 0.29 | 39 | ||
| mean | 3.75 | 1.56 | 0.28 | 0.27 | |||
| stdev | 0.47 | 0.29 | 0.15 | 0.23 | |||
| SE | 0.16 | 0.10 | 0.05 | 0.08 | |||
| Lantau | 1 | 4.43 | 1.67 | C. sinensis | 0 | 0 | <4 weeks |
| 3 | 4.43 | 1.59 | 0.009 | 0 | <6 weeks | ||
| 6 | 3.32 | 1.68 | 0.002 | 0 | <4 weeks | ||
| 11 | 3.62 | 1.63 | 0.001 | 0 | <4 weeks | ||
| 14 | 3 | 1.63 | 0.001 | 0 | <4 weeks | ||
| 16 | 3.06 | 1.32 | 0.002 | 0 | <4 weeks | ||
| 22 | 3.92 | 1.37 | 0 | 0 | <4 weeks | ||
| 27 | 3.79 | 1.15 | 0.002 | 0 | <4 weeks | ||
| Luk Wu | 1 | 3.86 | 2.12 | 0.002 | 0 | <4 weeks | |
| mean | 3.71 | 1.57 | 0.002 | 0 | |||
| std | 0.52 | 0.28 | 0.003 | 0 | |||
| SE | 0.19 | 0.10 | 0.001 | 0 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wineriter-Wright, S.A.; Smith, M.C.; Metz, M.A.; Makinson, J.R.; Brown, B.T.; Purcell, M.F.; Barr, K.L.; Pratt, P.D. The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate. Insects 2020, 11, 653. https://doi.org/10.3390/insects11100653
Wineriter-Wright SA, Smith MC, Metz MA, Makinson JR, Brown BT, Purcell MF, Barr KL, Pratt PD. The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate. Insects. 2020; 11(10):653. https://doi.org/10.3390/insects11100653
Chicago/Turabian StyleWineriter-Wright, Susan A., Melissa C. Smith, Mark A. Metz, Jeffrey R. Makinson, Bradley T. Brown, Matthew F. Purcell, Kane L. Barr, and Paul D. Pratt. 2020. "The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate" Insects 11, no. 10: 653. https://doi.org/10.3390/insects11100653
APA StyleWineriter-Wright, S. A., Smith, M. C., Metz, M. A., Makinson, J. R., Brown, B. T., Purcell, M. F., Barr, K. L., & Pratt, P. D. (2020). The Biology of Casmara subagronoma (Lepidoptera: Oecophoridae), a Stem-Boring Moth of Rhodomyrtus tomentosa (Myrtaceae): Descriptions of the Previously Unknown Adult Female and Immature Stages, and Its Potential as a Biological Control Candidate. Insects, 11(10), 653. https://doi.org/10.3390/insects11100653