Interpreting Morphological Adaptations Associated with Viviparity in the Tsetse Fly Glossina morsitans (Westwood) by Three-Dimensional Analysis

 ,
,  , , ,
, , ,
Simple Summary
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Biological Materials
2.2. Sample Collection
2.3. Sample Preparation, Fixation and Staining
2.4. Phase Contrast Micro-Computed Tomography
2.5. Data Processing, Segmentation, Visualization and Analysis
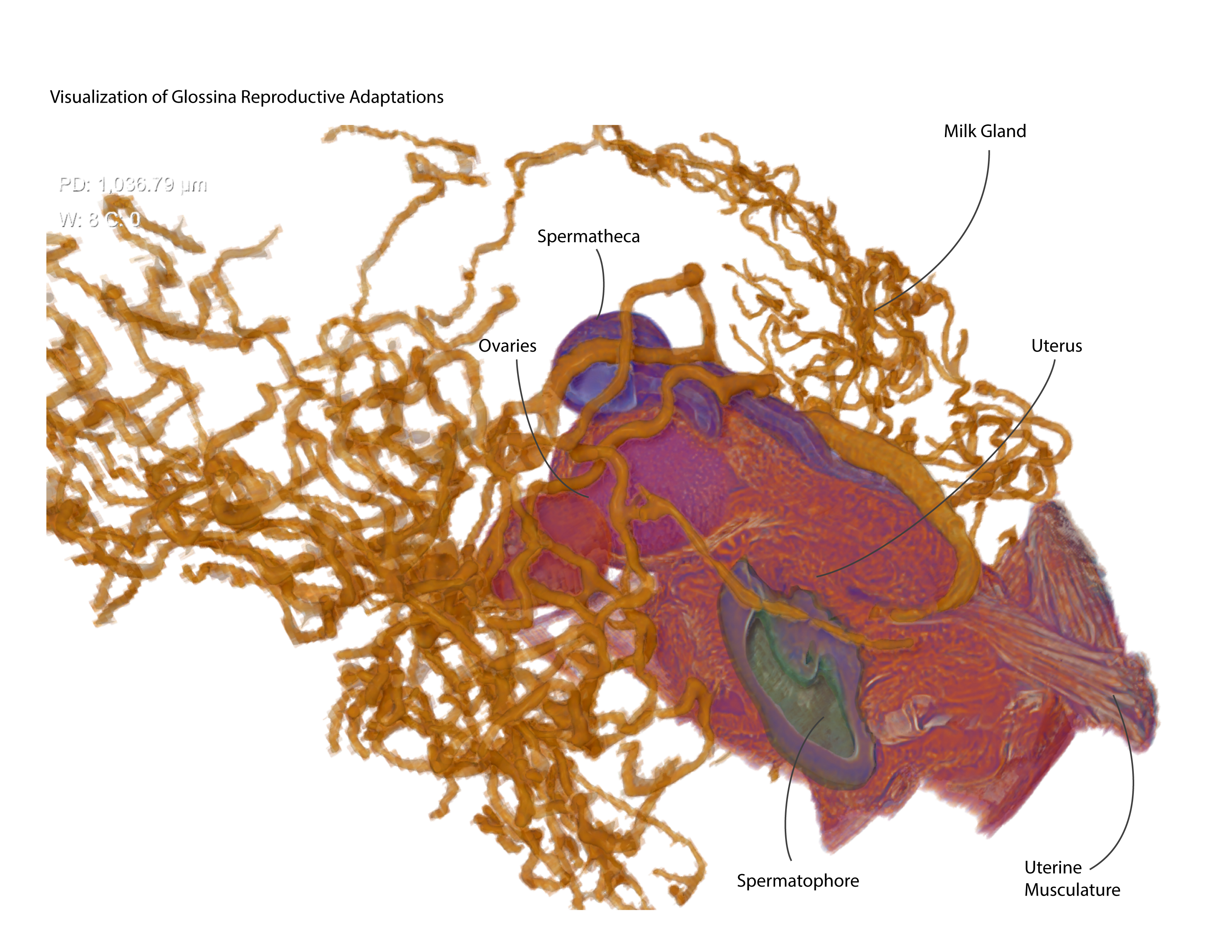
3. Results and Discussion
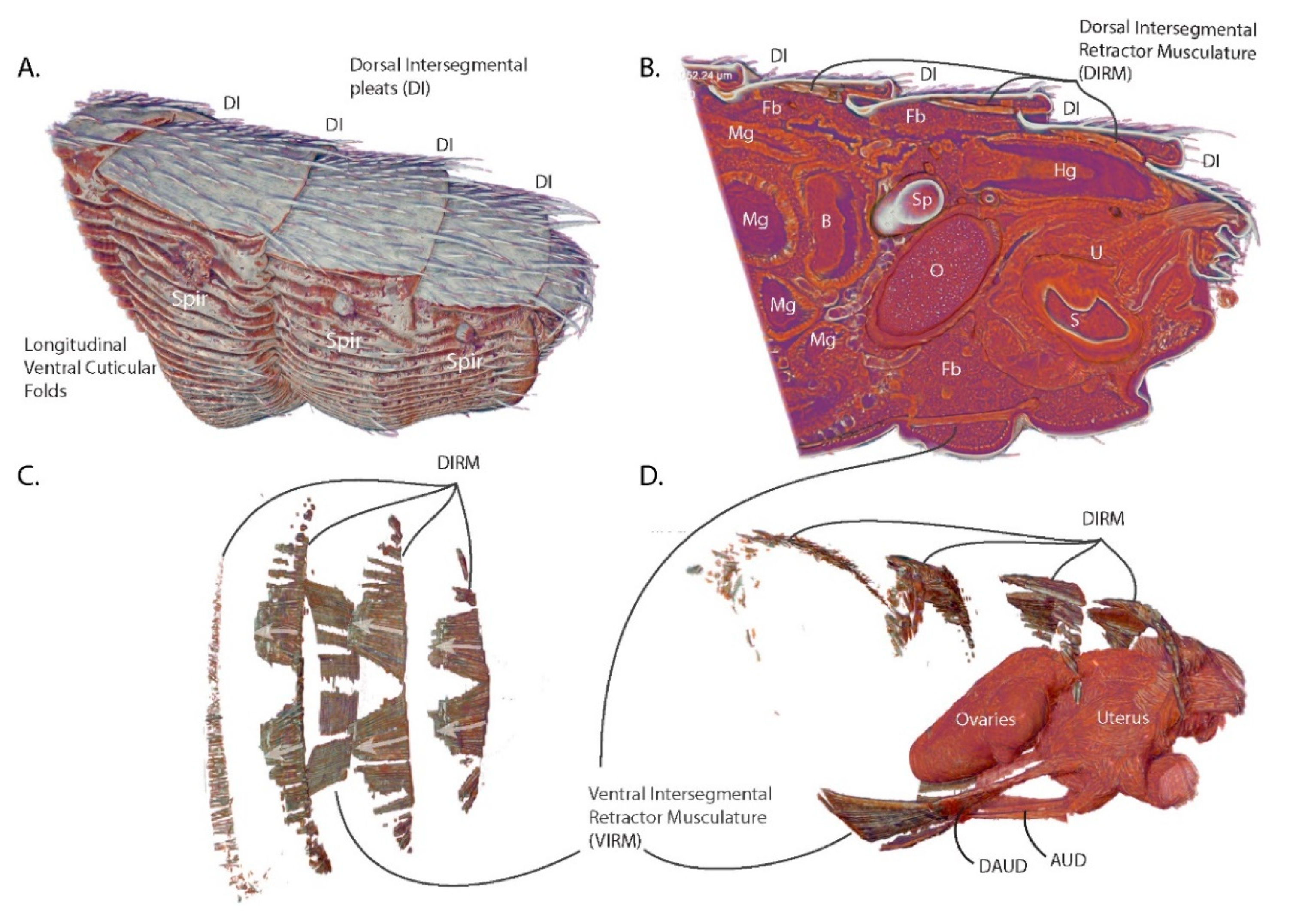
3.1. Abdominal Structural and Cuticular Adaptations for Blood Feeding and Pregnancy
3.2. Functional Aspects of G. morsitans Abdominal Musculature
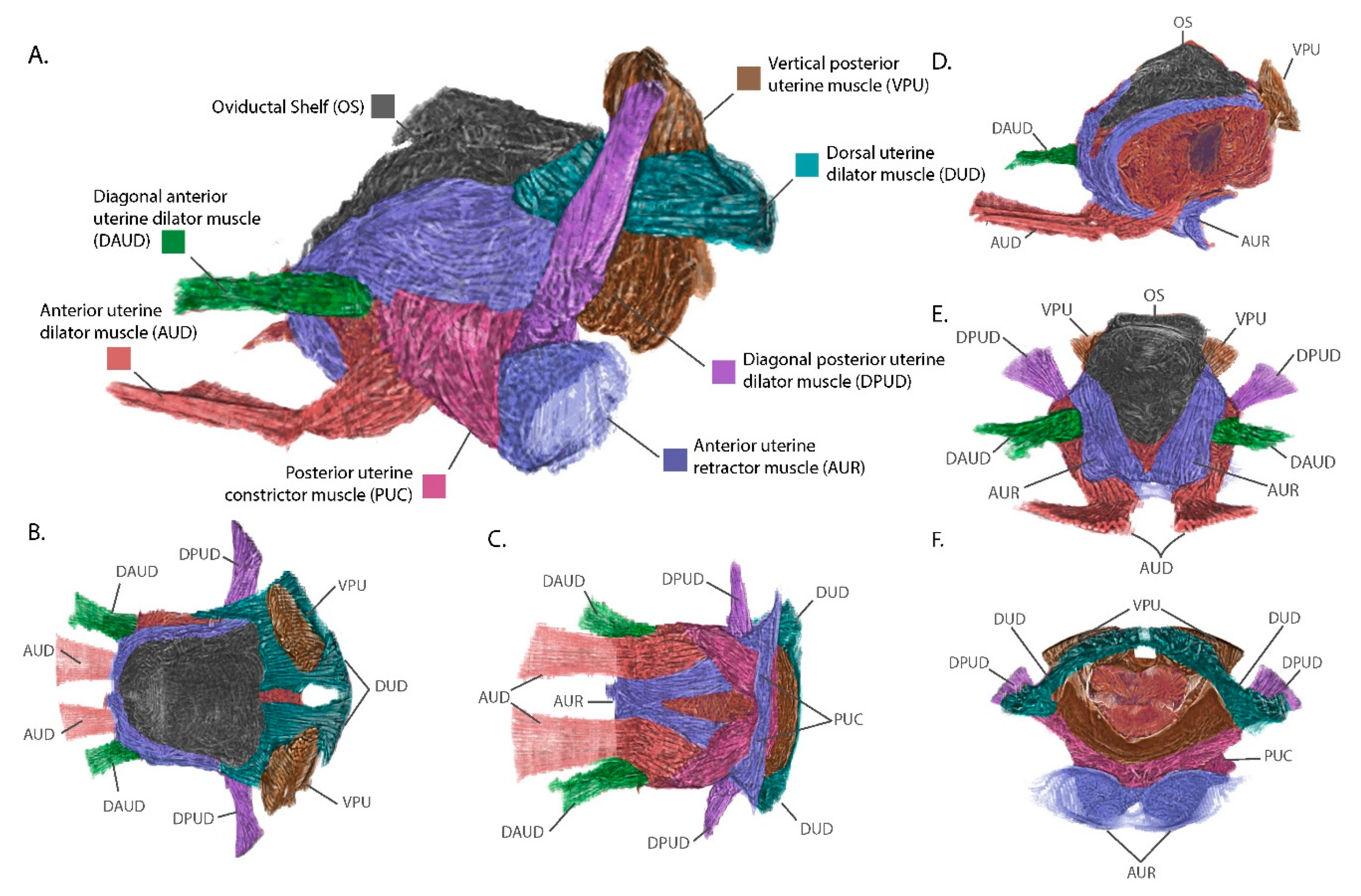
3.3. The External Uterine Musculature is Optimized for Intrauterine Larval Development and Parturition
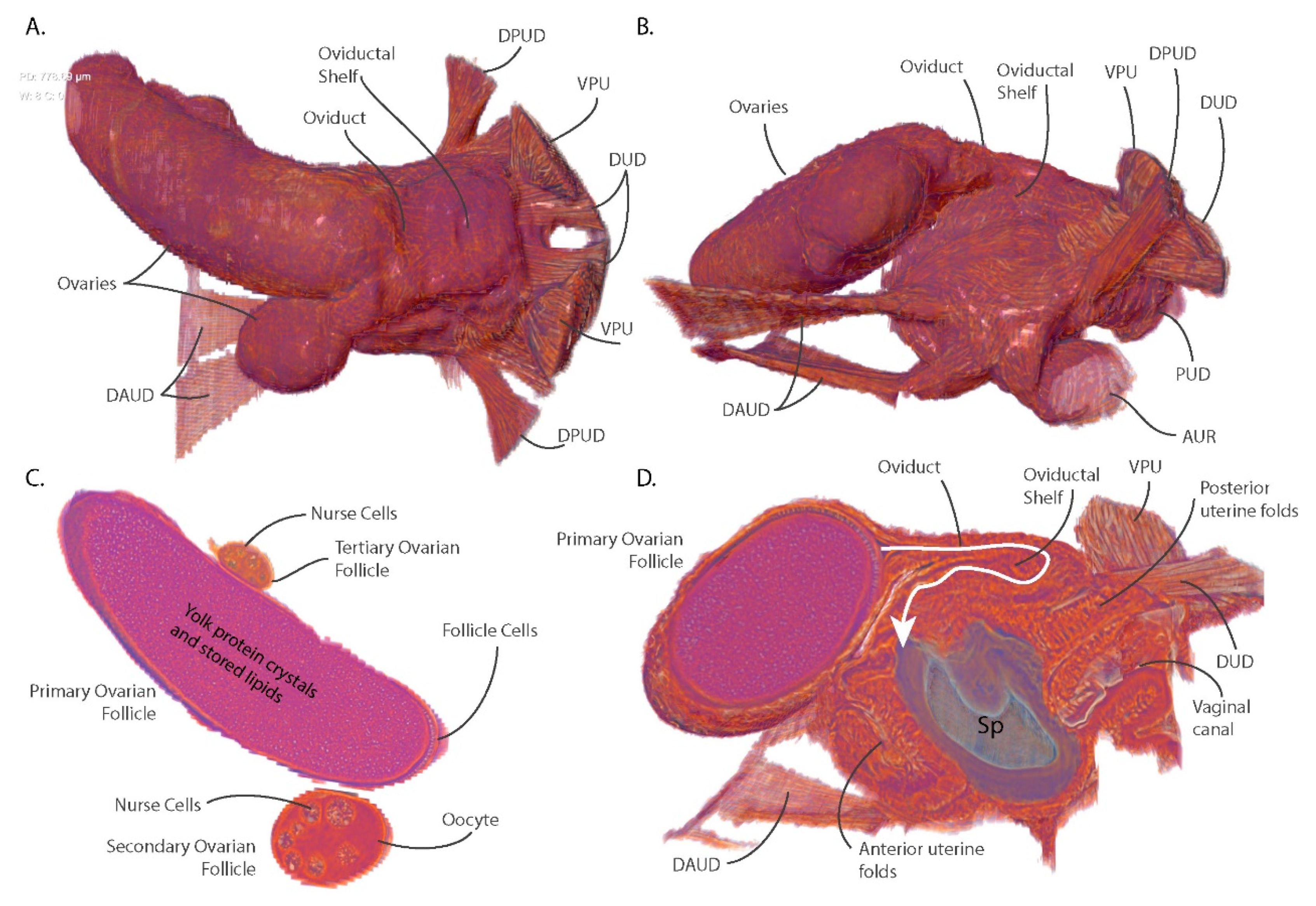
3.4. Ovarian Reduction and Oviduct Folding Reduce Constraints on Blood Meal Volume and Larval Size
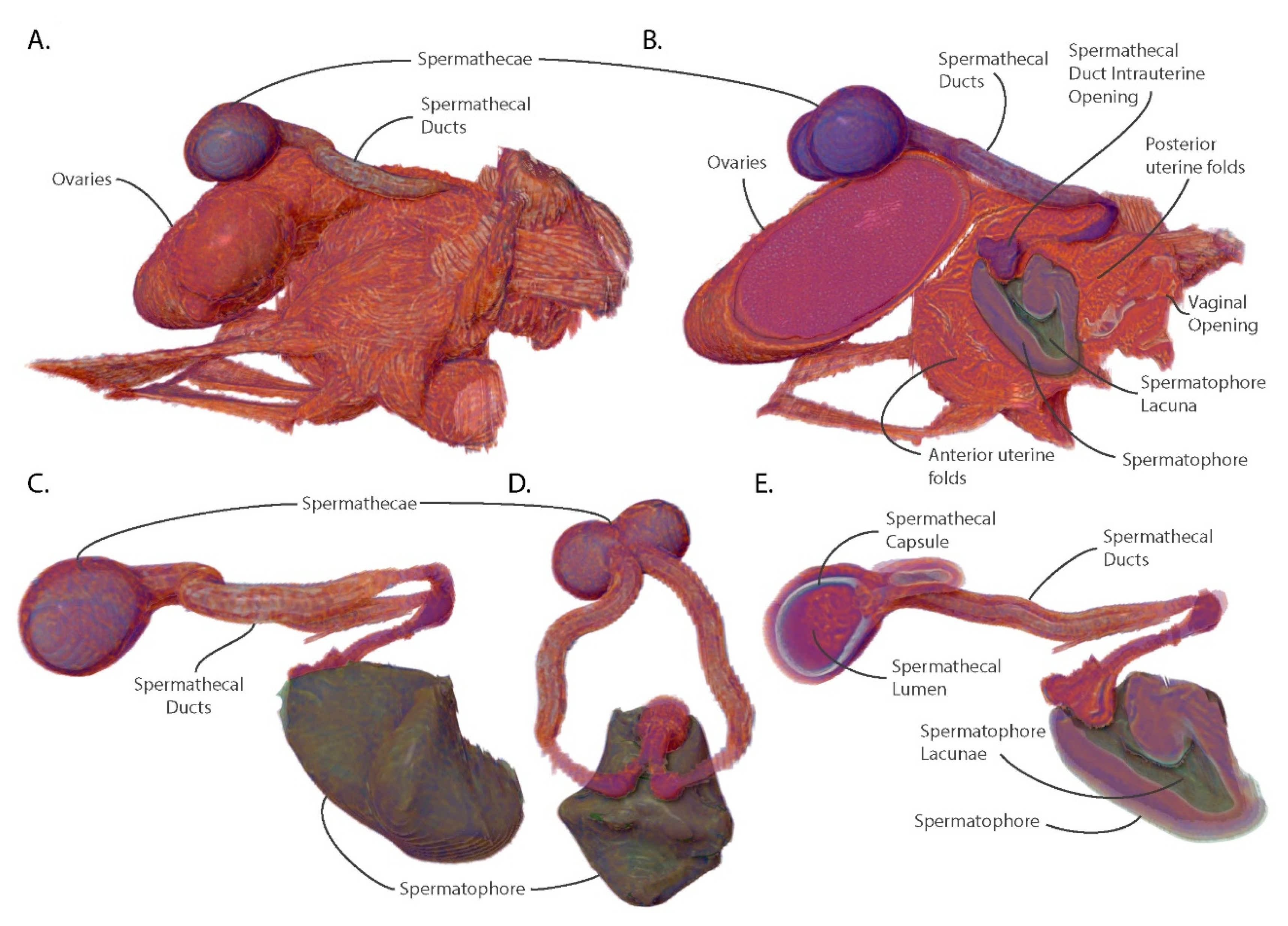
3.5. The Spermatophore Facilitates Sperm Storage within the Spermatheca and Acts as a Physical Barrier to Insemination by Competing Males
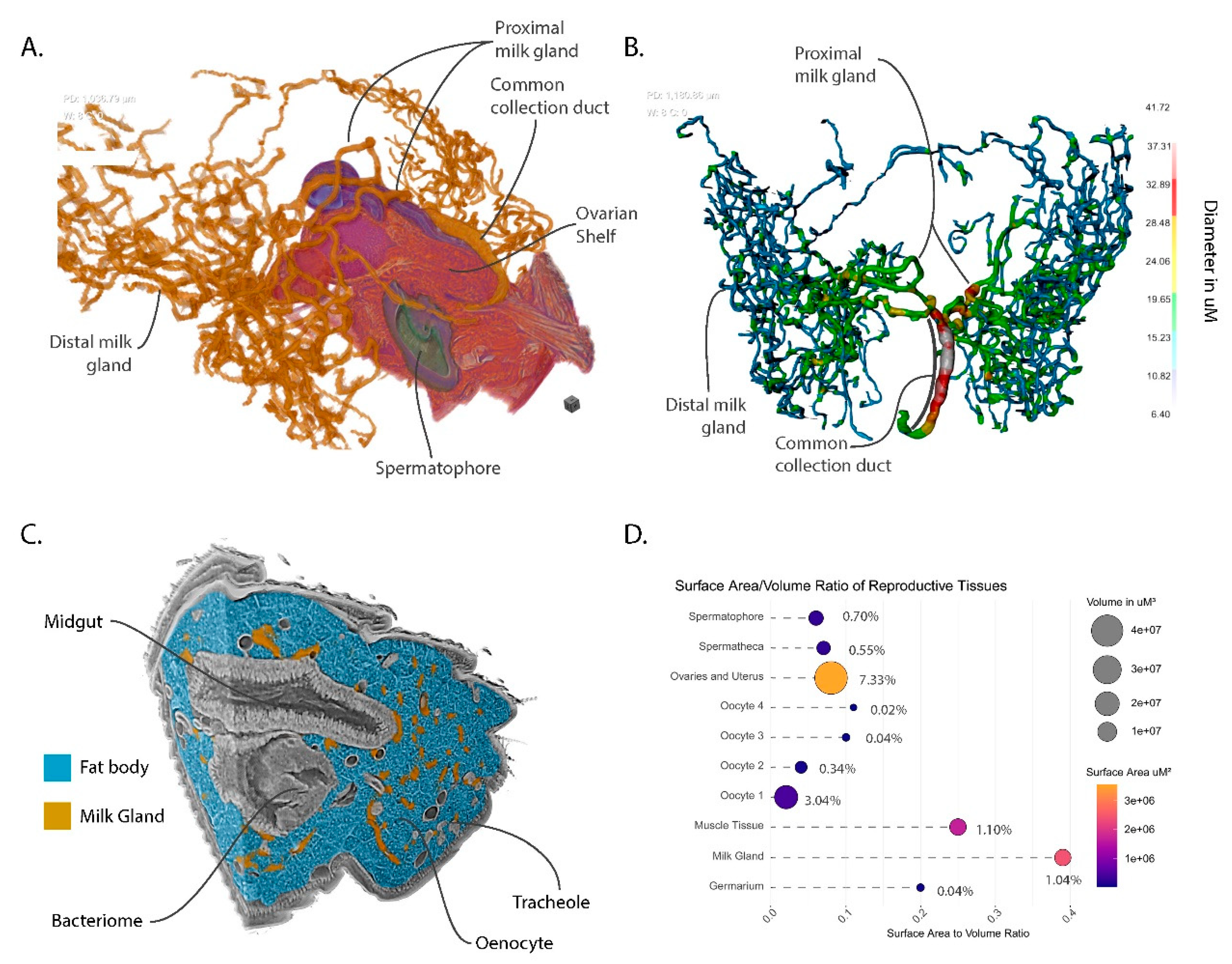
3.6. The Milk Gland Organ Maximizes Nutrient Incorporation via Extensive Ramification for Increased Surface Area and Intimate Contact with Fat Storage Tissues
3.7. Volumetric Relationships and Functional Implications
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aksoy, S.; Buscher, P.; Lehane, M.; Solano, P.; Van Den Abbeele, J. Human African trypanosomiasis control: Achievements and challenges. PLoS Negl. Trop. Dis. 2017, 11, e0005454. [Google Scholar] [CrossRef]
- Tirados, I.; Esterhuizen, J.; Kovacic, V.; Mangwiro, T.N.; Vale, G.A.; Hastings, I.; Solano, P.; Lehane, M.J.; Torr, S.J. Tsetse Control and Gambian Sleeping Sickness; Implications for Control Strategy. PLoS Negl. Trop. Dis. 2015, 9, e0003822. [Google Scholar] [CrossRef]
- Benoit, J.B.; Attardo, G.M.; Baumann, A.A.; Michalkova, V.; Aksoy, S. Adenotrophic viviparity in tsetse flies: Potential for population control and as an insect model for lactation. Annu. Rev. Entomol. 2015, 60, 351–371. [Google Scholar] [CrossRef]
- Langley, P.A. Physiology of tsetse flies (Glossina spp.) (Diptera: Glossinidae): A review. Bull. Entomol. Res. 1977, 67, 523–574. [Google Scholar] [CrossRef]
- Denlinger, D.L.; Ma, W.C. Dynamics of the pregnancy cycle in the tsetse Glossina morsitans. J. Insect Physiol. 1974, 20, 1015–1026. [Google Scholar] [CrossRef]
- Vreysen, M.J.; Saleh, K.M.; Ali, M.Y.; Abdulla, A.M.; Zhu, Z.R.; Juma, K.G.; Dyck, V.A.; Msangi, A.R.; Mkonyi, P.A.; Feldmann, H.U. Glossina austeni (Diptera: Glossinidae) eradicated on the island of Unguja, Zanzibar, using the sterile insect technique. J. Econ. Entomol. 2000, 93, 123–135. [Google Scholar] [CrossRef]
- Vreysen, M.J. Principles of area-wide integrated tsetse fly control using the sterile insect technique. Med. Trop. 2001, 61, 397–411. [Google Scholar]
- Saunders, D.S. Ovaries of Glossina morsitans. Nature 1960, 185, 121–122. [Google Scholar] [CrossRef]
- Roberts, M.J. The role of the choriothete in tsetse flies. Parasitology 1972, 64, 23–36. [Google Scholar] [CrossRef]
- Tobe, S.S.; Davey, K.G.; Huebner, E. Nutrient transfer during the reproductive cycle in Glossina austeni Newst.: Histology and histochemistry of the milk gland, fat body, and oenocytes. Tissue Cell 1973, 5, 633–650. [Google Scholar] [CrossRef]
- Ma, W.C.; Denlinger, D.L. Secretory discharge and microflora of milk gland in tsetse flies. Nature 1974, 247, 301–303. [Google Scholar] [CrossRef]
- Ma, W.C.; Denlinger, D.L.; Jarlfors, U.; Smith, D.S. Structural modulations in the tsetse fly milk gland during a pregnancy cycle. Tissue Cell 1975, 7, 319–330. [Google Scholar] [CrossRef]
- Mattei, A.L.; Riccio, M.L.; Avila, F.W.; Wolfner, M.F. Integrated 3D view of postmating responses by the Drosophila melanogaster female reproductive tract, obtained by micro-computed tomography scanning. Proc. Natl. Acad. Sci. USA 2015, 112, 8475–8480. [Google Scholar] [CrossRef]
- Kapelnikov, A.; Zelinger, E.; Gottlieb, Y.; Rhrissorrakrai, K.; Gunsalus, K.C.; Heifetz, Y. Mating induces an immune response and developmental switch in the Drosophila oviduct. Proc. Natl. Acad. Sci. USA 2008, 105, 13912–13917. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, S. Establishment and Maintenance of Small Scale Tsetse Colonies. In Maintenance of Human, Animal and Plant Pathogen Vectors; Maramorosch, K., Mahmood, F., Eds.; Science Publishers, Inc.: Enfield, NH, USA, 1999; pp. 123–136. [Google Scholar]
- Saunders, D.S.; Dodd, C.W.H. Mating, Insemination, and Ovulation in Tsetse Fly, Glossina morsitans. J. Insect Physiol. 1972, 18, 187–198. [Google Scholar] [CrossRef]
- Pandolfi, R.J.; Allan, D.B.; Arenholz, E.; Barroso-Luque, L.; Campbell, S.I.; Caswell, T.A.; Blair, A.; De Carlo, F.; Fackler, S.; Fournier, A.P.; et al. Xi-cam: A versatile interface for data visualization and analysis. J. Synchrotron Radiat. 2018, 25, 1261–1270. [Google Scholar] [CrossRef] [PubMed]
- Gürsoy, D.; De Carlo, F.; Xiao, X.; Jacobsen, C. TomoPy: A framework for the analysis of synchrotron tomographic data. J. Synchrotron Radiat. 2014, 21, 1188–1193. [Google Scholar] [CrossRef]
- Tobe, S.S.; Davey, K.G. Volume Relationships During Pregnancy Cycle of Tsetse Fly Glossina austeni. Can. J. Zool. 1972, 50, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Pyenson, N.D.; Goldbogen, J.A.; Vogl, A.W.; Szathmary, G.; Drake, R.L.; Shadwick, R.E. Discovery of a sensory organ that coordinates lunge feeding in rorqual whales. Nature 2012, 485, 498–501. [Google Scholar] [CrossRef]
- Kubrakiewicz, J.; Bilinski, S.M.; Mazurkiewicz, M. Diptera—ovary structure and oogenesis in midges and flies. Folia Histochem. Cytobiol. 1998, 36, 197–203. [Google Scholar]
- Saunders, D.S. Mating, ovulation and oocyte development in Glossina morsitans. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 184–185. [Google Scholar] [CrossRef]
- Mellanby, H. Experimental work on reproduction in the tsetse fly, Glossina palpalis. Parasitology 1937, 1, 131–141. [Google Scholar] [CrossRef]
- Gillott, C.; Langley, P.A. The control of receptivity and ovulation in the tsetse fly, Glossina morsitans. Physiol. Entomol. 1981, 6, 269–281. [Google Scholar] [CrossRef]
- Ejezie, G.C.; Davey, K.G. Some effects of mating in female tsetse, Glossina austeni Newst. J. Exp. Zool. 1977, 200, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Odhiambo, T.R. Regulation of Ovulation in Tsetse-Fly, Glossina pallidipes Austen. J. Exp. Zool. 1971, 177, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Wall, R.; Langley, P.A. The Mating-Behavior of Tsetse-Flies (Glossina)—A Review. Physiol. Entomol. 1993, 18, 211–218. [Google Scholar] [CrossRef]
- Chaudhury, M.F.B.; Dhadialla, T.S.; Kunyiha, R.W. Evidence of Neuroendocrine Relationships Between Mating and Ovulation in the Tsetse-Fly, Glossina morsitans morsitans. Insect Sci. Appl. 1981, 1, 161–166. [Google Scholar] [CrossRef]
- Pollock, J.N. Sperm transfer by spermatophores in Glossina austeni newstead. Nature 1970, 225, 1063–1064. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.N. Spermatophores in tsetse flies. Trans. R. Soc. Trop. Med. Hyg. 1972, 66, 315. [Google Scholar] [CrossRef]
- Scolari, F.; Benoit, J.B.; Michalkova, V.; Aksoy, E.; Takac, P.; Abd-Alla, A.M.; Malacrida, A.R.; Aksoy, S.; Attardo, G.M. The Spermatophore in Glossina morsitans morsitans: Insights into Male Contributions to Reproduction. Sci. Rep. 2016, 6, 20334. [Google Scholar] [CrossRef]
- Odhiambo, T.R.; Kokwaro, E.D.; Sequeira, L.M. Histochemical and Ultrastructural Studies of the Male Accessory Reproductive Glands and Spermatophore of the Tsetse, Glossina morsitans morsitans Westwood. Insect Sci. Appl. 1983, 4, 227–236. [Google Scholar] [CrossRef]
- Kokwaro, E.D.; Odhiambo, T.R. Spermatophore of the tsetse, Glossina morsitans morsitans Westwood: An ultrastructural study. Int. J. Trop. Insect Sci. 1981, 1, 185–190. [Google Scholar] [CrossRef]
- Mann, T. Spermatophores: Development, Structure, Biochemical Attributes and Role in the Transfer of Spermatozoa; Springer: Berlin/Heidelberg, Germany, 1984; ISBN 9783540135838. [Google Scholar]
- Briceño, R.D.; Eberhard, W.G. Experimental demonstration of possible cryptic female choice on male tsetse fly genitalia. J. Insect Physiol. 2009, 55, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Abd-Alla, A.M.M.; Acosta-Serrano, A.; Allen, J.E.; Bateta, R.; Benoit, J.B.; Bourtzis, K.; Caers, J.; Caljon, G.; Christensen, M.B.; et al. Comparative genomic analysis of six Glossina genomes, vectors of African trypanosomes. Genome Biol. 2019, 20, 187. [Google Scholar] [CrossRef] [PubMed]
- Cmelik, S.H.; Hurrell, D.P.; Lunat, M. Lipid content and composition of the tse-tse fly, Glossina morsitans Westwood. Comp. Biochem. Physiol. 1969, 31, 65–78. [Google Scholar] [CrossRef]
- Cmelik, S.H.W.; Bursell, E.; Slack, E. Composition of Gut Contents of Third-Instar Tsetse Larvae (Glossina morsitans Westwood). Comp. Biochem. Physiol. 1969, 29, 447–453. [Google Scholar] [CrossRef]
- Attardo, G.M.; Lohs, C.; Heddi, A.; Alam, U.H.; Yildirim, S.; Aksoy, S. Analysis of milk gland structure and function in Glossina morsitans: Milk protein production, symbiont populations and fecundity. J. Insect Physiol. 2008, 54, 1236–1242. [Google Scholar] [CrossRef]
- Pais, R.; Lohs, C.; Wu, Y.; Wang, J.; Aksoy, S. The obligate mutualist Wigglesworthia glossinidia influences reproduction, digestion, and immunity processes of its host, the tsetse fly. Appl. Environ. Microbiol. 2008, 74, 5965–5974. [Google Scholar] [CrossRef]
- Slobounov, S.M.; Ray, W.; Johnson, B.; Slobounov, E.; Newell, K.M. Modulation of cortical activity in 2D versus 3D virtual reality environments: An EEG study. Int. J. Psychophysiol. 2015, 95, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Laha, B.; Sensharma, K.; Schiffbauer, J.D.; Bowman, D.A. Effects of immersion on visual analysis of volume data. IEEE Trans. Vis. Comput. Graph. 2012, 18, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Krokos, E.; Plaisant, C.; Varshney, A. Virtual memory palaces: Immersion aids recall. Virtual Real. 2019, 23, 1–15. [Google Scholar] [CrossRef]
- Metscher, B.D.; Muller, G.B. MicroCT for molecular imaging: Quantitative visualization of complete three-dimensional distributions of gene products in embryonic limbs. Dev. Dyn. 2011, 240, 2301–2308. [Google Scholar] [CrossRef] [PubMed]
- Martín-Vega, D.; Clark, B.; Ferrer, L.M.; López-Tamayo, S.; Panadero, R.; Cepeda-Palacios, R.; Colwell, D.D.; Hall, M.J.R. Major differences in the larval anatomy of the digestive and excretory systems of three Oestridae species revealed by micro-CT. Med. Vet. Entomol. 2020. [Google Scholar] [CrossRef]
- Tiulpin, A.; Finnilä, M.; Lehenkari, P.; Nieminen, H.J.; Saarakkala, S. Deep-Learning for Tidemark Segmentation in Human Osteochondral Tissues Imaged with Micro-computed Tomography. In Lecture Notes in Computer Science, Proceedings of the Advanced Concepts for Intelligent Vision Systems, Auckland, New Zealand, 10–14 February 2020; Springer International Publishing: Cham, Switzerland, 2020; pp. 131–138. [Google Scholar]
- Fredericksen, M.A.; Zhang, Y.; Hazen, M.L.; Loreto, R.G.; Mangold, C.A.; Chen, D.Z.; Hughes, D.P. Three-dimensional visualization and a deep-learning model reveal complex fungal parasite networks in behaviorally manipulated ants. Proc. Natl. Acad. Sci. USA 2017, 114, 12590–12595. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attardo, G.M.; Tam, N.; Parkinson, D.; Mack, L.K.; Zahnle, X.J.; Arguellez, J.; Takáč, P.; Malacrida, A.R. Interpreting Morphological Adaptations Associated with Viviparity in the Tsetse Fly Glossina morsitans (Westwood) by Three-Dimensional Analysis. Insects 2020, 11, 651. https://doi.org/10.3390/insects11100651
Attardo GM, Tam N, Parkinson D, Mack LK, Zahnle XJ, Arguellez J, Takáč P, Malacrida AR. Interpreting Morphological Adaptations Associated with Viviparity in the Tsetse Fly Glossina morsitans (Westwood) by Three-Dimensional Analysis. Insects. 2020; 11(10):651. https://doi.org/10.3390/insects11100651
Chicago/Turabian StyleAttardo, Geoffrey M, Nicole Tam, Dula Parkinson, Lindsey K Mack, Xavier J Zahnle, Joceline Arguellez, Peter Takáč, and Anna R Malacrida. 2020. "Interpreting Morphological Adaptations Associated with Viviparity in the Tsetse Fly Glossina morsitans (Westwood) by Three-Dimensional Analysis" Insects 11, no. 10: 651. https://doi.org/10.3390/insects11100651
APA StyleAttardo, G. M., Tam, N., Parkinson, D., Mack, L. K., Zahnle, X. J., Arguellez, J., Takáč, P., & Malacrida, A. R. (2020). Interpreting Morphological Adaptations Associated with Viviparity in the Tsetse Fly Glossina morsitans (Westwood) by Three-Dimensional Analysis. Insects, 11(10), 651. https://doi.org/10.3390/insects11100651