Effects of Host Plants Reared under Elevated CO2 Concentrations on the Foraging Behavior of Different Stages of Corn Leaf Aphids Rhopalosiphum maidis

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. CO2 Condition Chambers
2.2. Plant Material
2.3. Aphid Rearing
2.4. Foraging Behavior Bioassay
2.5. Headspace Analysis of Volatiles from Plants by GC-MS
2.6. Statistical Analyses
3. Results
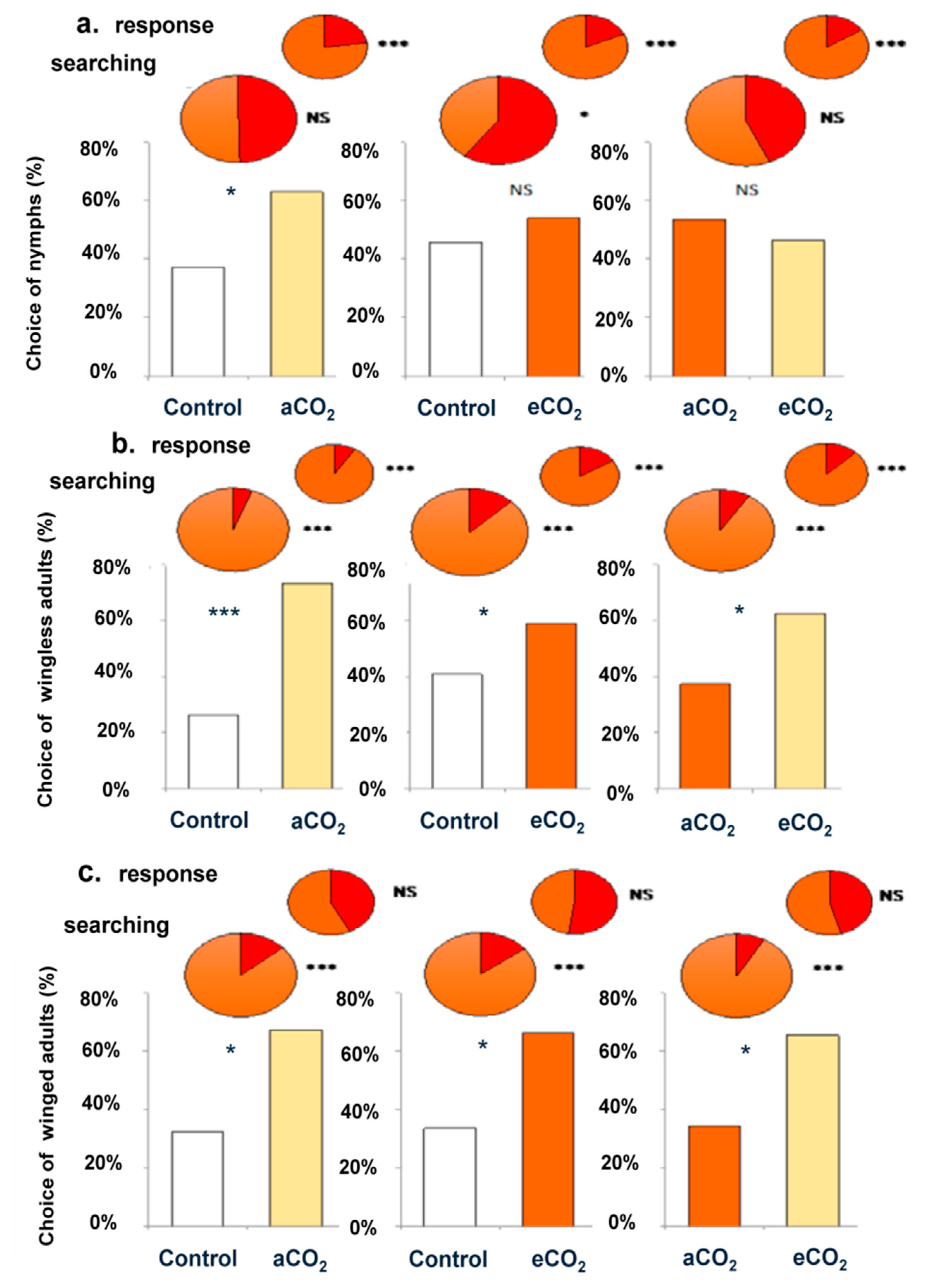
3.1. Foraging Behaviors of Aphids
3.2. Residence Duration for Searching and Selection Behaviors
3.3. Volatiles Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stocker, T.; Qin, D.; Plattner, G.; Tignor, M.; Allen, S.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, V.; Midgley, P. IPCC, 2013: Climate Change 2013: The physical science basis. contributionof working group i to the fifth assessment report of the intergovernmental panel on climate change. Comput. Geom. 2013, 18, 95–123. [Google Scholar]
- Hartley, S.E.; Jones, C.G.; Couper, G.C.; Jones, T.H. Biosynthesis of plant phenolic compounds in elevated atmospheric CO2. Glob. Chang. Biol. 2000, 6, 497–506. [Google Scholar] [CrossRef]
- Bae, H.; Sicher, R. Changes of soluble protein expression and leaf metabolite levels in Arabidopsis thaliana grown in elevated atmospheric carbon dioxide. Field Crops Res. 2004, 90, 61–73. [Google Scholar] [CrossRef]
- Seneweera, S.P.; Conroy, J.P. Enhanced leaf elongation rates of wheat at elevated CO2: Is it related to carbon and nitrogen dynamics within the growing leaf blade? Environ. Exp. Bot. 2005, 54, 174–181. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising CO2: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Johns, C.V.; Hughes, L. Interactive effects of elevated CO2 and temperature on the leaf-miner Dialectica scalariella Zeller (Lepidoptera: Gracillariidae) in Paterson’s Curse, Echium plantagineum (Boraginaceae). Glob. Chang. Biol. 2002, 8, 142–152. [Google Scholar] [CrossRef]
- Chen, F.; Ge, F.; Parajulee, M.N. Impact of elevated CO2 on tri-trophic interaction of Gossypium hirsutum, Aphis gossypii, and Leis axyridis. Environ. Entomol. 2005, 34, 37–46. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, H.; Yuan, E.; Ge, F. Elevated CO2 increases R gene-dependent resistance of Medicago truncatula against the pea aphid by up-regulating a heat shock gene. New Phytol. 2018, 217, 1696–1711. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, Y.; Wan, G.; Liu, B.; Xing, G.; Chen, F. Effects of elevated CO2 on plant chemistry, growth, yield of resistant soybean, and feeding of a target lepidoptera pest, Spodoptera litura (Lepidoptera: Noctuidae). Environ. Entomol. 2018, 47, 848–856. [Google Scholar]
- Drake, B.G.; Azcon-Bieto, J.; Berry, J.; Bunce, J.; Dijkstra, P.; Farrar, J.; Gifford, R.; Gonzalez-meler, M.A.; Koch, G.; Lambers, H. Does elevated atmospheric CO2 concentration inhibit mitochondrial respiration in green plants? Plant Cell Environ. 1999, 22, 649–657. [Google Scholar] [CrossRef]
- Ziska, L.H.; Namuco, O.; Moya, T.; Quilang, J. Growth and yield response of field-grown tropical rice to increasing carbon dioxide and air temperature. Agron. J. 1997, 89, 45–53. [Google Scholar] [CrossRef]
- Kim, K.N.; Cheong, Y.H.; Grant, J.J.; Pandey, G.K.; Luan, S. CIPK3, a calcium sensor-associated protein kinase that regulates abscisic acid and cold signal transduction in Arabidopsis. Plant Cell 2003, 15, 411–423. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Karowe, D.N.; Seimens, D.H.; Mitchell-Olds, T. Species-specific response of glucosinolate content to elevated atmospheric CO2. J. Chem. Ecol. 1997, 23, 2569–2582. [Google Scholar] [CrossRef]
- Coviella, C.E.; Stipanovic, R.D.; Trumble, J.T. Plant allocation to defensive compounds: Interactions between elevated CO2 and nitrogen in transgenic cotton plants. J. Exp. Bot. 2002, 53, 323–331. [Google Scholar] [CrossRef]
- Karowe, D.N.; Grubb, C. Elevated CO2 increases constitutive phenolics and trichomes, but decreases inducibility of phenolics in Brassica rapa (Brassicaceae). J. Chem. Ecol. 2011, 37, 1332–1340. [Google Scholar] [CrossRef]
- Stiling, P.; Cornelissen, T. How does elevated carbon dioxide CO2 affect plant–herbivore interactions? A field experiment and meta-analysis of CO2-mediated changes on plant chemistry and herbivore performance. Glob. Chang. Biol. 2007, 13, 1823–1842. [Google Scholar] [CrossRef]
- Robinson, M.T.; Weeks, A.R.; Hoffmann, A.A. Geographic patterns of clonal diversity in the earth mite species Penthaleus major with particular emphasis on species margins. Evolution 2002, 56, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Bidart-Bouzat, M.G.; Imeh-Nathaniel, A. Global change effects on plant chemical defenses against insect herbivores. J Integr. Plant Biol. 2008, 50, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B.; Wicher, D. Chemical ecology in insects. In Chemosensory Transduction: The Detection of Odors, Tastes, and other Chemostimuli; Zufall, F., Munger, S.D., Eds.; Academic press: London, UK, 2006; pp. 29–45. [Google Scholar]
- Qualley, A.V.; Dudareva, N. Metabolomics of plant volatiles. Methods Mol. Biol. 2009, 553, 329–343. [Google Scholar]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents-a checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Holopainen, J.K.; Heijari, J.; Oksanen, E.; Alessio, G.A. Leaf volatile emissions of Betula pendula during autumn coloration and leaf fall. J. Chem. Ecol. 2010, 36, 1068–1075. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Martínez, E.; Bosque-Pérez, N.; Berger, P.; Zemetra, R. Life history of the bird cherry-oat aphid, Rhopalosiphum padi (Homoptera: Aphididae), on transgenic and untransformed wheat challenged with Barley yellow dwarf virus. J. Econ. Entomol. 2004, 97, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. Sites of synthesis, biochemistry and functional role of plant volatiles. S. Afr. J. Bot. 2010, 76, 612–631. [Google Scholar] [CrossRef]
- Boullis, A.; Detrain, C.; Francis, F.; Verheggen, F.J. Will climate change affect insect pheromonal communication? Curr. Opin. Insect Sci. 2016, 17, 87–91. [Google Scholar] [CrossRef]
- Harrewijn, P.; Minks, A.K. Aphids: Their Biology, Natural Enemies, and Control, Vol. A.; Elsevier: Amsterdam, The Netherlands, 1987. [Google Scholar]
- Nault, L. Arthropod transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Braendle, C.; Davis, G.K.; Brisson, J.A.; Stern, D.L. Wing dimorphism in aphids. Heredity 2006, 97, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Pickett, J.; Wadhams, L.; Woodcock, C.; Hardie, J. The chemical ecology of aphids. Annu. Rev. Entomol. 1992, 37, 67–90. [Google Scholar] [CrossRef]
- Pettersson, J.; Ninkovic, V.; Glinwood, R.; Al Abassi, S.; Birkett, M.; Pickett, J.; Wadhams, L. Chemical stimuli supporting foraging behaviour of Coccinella septempunctata L. (Coleoptera: Coccinellidae): Volatiles and allelobiosis. Appl. Entomol. Zool. 2008, 43, 315–321. [Google Scholar] [CrossRef][Green Version]
- Webster, B. The role of olfaction in aphid host location. Physiol. Entomol. 2012, 37, 10–18. [Google Scholar] [CrossRef]
- Quiroz, A.; Niemeyer, H. Olfactometer-assessed responses of aphid Rhopalosiphum padi to wheat and oat volatiles. J. Chem. Ecol. 1998, 24, 113–124. [Google Scholar] [CrossRef]
- Boullis, A.; Francis, F.; Verheggen, F.J. Climate change and tritrophic interactions: Will modifications to greenhouse gas emissions increase the vulnerability of herbivorous insects to natural enemies? Environ. Entomol. 2015, 44, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Boullis, A.; Fassotte, B.; Sarles, L.; Lognay, G.; Heuskin, S.; Vanderplanck, M.; Bartram, S.; Haubruge, E.; Francis, F.; Verheggen, F.J. Elevated carbon dioxide concentration reduces alarm signaling in aphids. J. Chem. Ecol. 2017, 43, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Boullis, A.; Blanchard, S.; Francis, F.; Verheggen, F. Elevated CO2 concentrations impact the semiochemistry of aphid honeydew without having a cascade effect on an aphid predator. Insects 2018, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, S.; Lognay, G.; Verheggen, F.; Detrain, C. Today and tomorrow: Impact of climate change on aphid biology and potential consequences on their mutualism with ants. Physiol. Entomol. 2019, 44, 77–86. [Google Scholar] [CrossRef]
- El-Ibrashy, M.T.; El-ziadys, S.; Riad, A.A. Laboratory studies on the biology of the corn leaf aphid, Rhopalosiphum maidis (Homoptera: Aphididae). Entomol. Exp. Appl. 1972, 15, 166–174. [Google Scholar] [CrossRef]
- Everly, R.T. Loss in corn yield associated with the abundance of the corn leaf aphid, Rhopalosiphum maidis, in Indiana. J. Econ. Entomol. 1960, 53, 924–932. [Google Scholar] [CrossRef]
- Foott, W.H.; Timmins, P.R. Effects of infestations by the corn leaf aphid, Rhopalosiphum maidis (Homoptera: Aphididae), on field corn in southwestern Ontario. Can. Entomol. 1973, 105, 449–458. [Google Scholar] [CrossRef]
- Bing, J.W.; Guthrie, W.D.; Dicke, F.F.; Obrycki, J.J. Seedling stage feeding by corn leaf aphid (Homoptera: Aphididae): Influence on plant development in maize. J. Econ. Entomol. 1991, 84, 625–632. [Google Scholar] [CrossRef]
- Tottman, D.; Makepeace, R.; Broad, H. An explanation of the decimal code for the growth stages of cereals, with illustrations. Ann. Appl. Biol. 1979, 93, 221–234. [Google Scholar] [CrossRef]
- Powell, G.; Hardie, J.; Pickett, J. Behavioural evidence for detection of the repellent polygodial by aphid antennal tip sensilla. Physiol. Entomol. 1995, 20, 141–146. [Google Scholar] [CrossRef]
- Pettersson, J.; Tjallingii, W.F.; Hardie, J. Host-Plant Selection and Feeding. In Aphids as Crop Pests; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2007; pp. 87–113. [Google Scholar]
- Park, K.C.; Hardie, J. Electrophysiological characterisation of olfactory sensilla in the black bean aphid, Aphis fabae. J. Insect Physiol. 2004, 50, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Wohlers, P.; Tjallingii, W. Electroantennogram responses of aphids to the alarm pheromone (E)-β-farnesene. Entomol. Exp. Appl. 1983, 33, 79–82. [Google Scholar] [CrossRef]
- Babikova, Z.; Gilbert, L.; Randall, K.C.; Bruce, T.J.; Pickett, J.A.; Johnson, D. Increasing phosphorus supply is not the mechanism by which arbuscular mycorrhiza increase attractiveness of bean (Vicia faba) to aphids. J. Exp. Bot. 2014, 65, 5231–5241. [Google Scholar] [CrossRef] [PubMed]
- Harmel, N.; Almohamad, R.; Fauconnier, M.L.; Du Jardin, P.; Verheggen, F.; Marlier, M.; Haubruge, E.; Francis, F. Role of terpenes from aphid-infested potato on searching and oviposition behavior of Episyrphus balteatus. Insect Sci. 2007, 14, 57–63. [Google Scholar] [CrossRef]
- Halbert, S.; Corsini, D.; Wiebe, M.; Vaughn, S. Plant-derived compounds and extracts with potential as aphid repellents. Ann. Appl. Biol. 2009, 154, 303–307. [Google Scholar] [CrossRef]
- Bukovinszky, T.; Gols, R.; Posthumus, M.; Vet, L.; Van Lenteren, J. Variation in plant volatiles and attraction of the parasitoid Diadegma semiclausum (Hellen). J. Chem. Ecol. 2005, 31, 461–480. [Google Scholar] [CrossRef] [PubMed]
- Wenda-Piesik, A.; Piesik, D.; Ligor, T.; Buszewski, B. Volatile organic compounds (VOCs) from cereal plants infested with crown rot: Their identity and their capacity for inducing production of VOCs in uninfested plants. Int. J. Pest Manag. 2010, 56, 377–383. [Google Scholar] [CrossRef]
- Piesik, D.; Łyszczarz, A.; Tabaka, P.; Lamparski, R.; Bocianowski, J.; Delaney, K. Volatile induction of three cereals: Influence of mechanical injury and insect herbivory on injured plants and neighbouring uninjured plants. Ann. Appl. Biol. 2010, 157, 425–434. [Google Scholar] [CrossRef]
- Visser, J.; Taanman, J. Odour-conditioned anemotaxis of apterous aphids (Cryptomyzus korschelti) in response to host plants. Physiol. Entomol. 1987, 12, 473–479. [Google Scholar] [CrossRef]
- Nottingham, S.F.; Hardie, J.; Dawson, G.W.; Hick, A.J.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Behavioral and electrophysiological responses of aphids to host and nonhost plant volatiles. J. Chem. Ecol. 1991, 17, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Storer, J.R.; Young, S.; Hardie, J. Three-dimensional analysis of aphid landing behaviour in the laboratory and field. Physiol. Entomol. 1999, 24, 271–277. [Google Scholar] [CrossRef]
- Vargas, R.R.; Troncoso, A.J.; Tapia, D.H.; Olivares-Donoso, R.; Niemeyer, H.M. Behavioural differences during host selection between alate virginoparae of generalist and tobacco-specialist Myzus persicae. Entomol. Exp. Appl. 2005, 116, 43–53. [Google Scholar] [CrossRef]
- Goldansaz, S.H.; McNeil, J.N. Effect of wind speed on the pheromone-mediated behavior of sexual morphs of the potato aphid, Macrosiphum euphorbiae (Thomas) under laboratory and field conditions. J. Chem. Ecol. 2006, 32, 1719. [Google Scholar] [CrossRef] [PubMed]
- Hardie, J.; Gao, N.; Timar, T.; Sebok, P.; Honda, K.I. Precocene derivatives and aphid morphogenesis. Arch. Insect Biochem. Physiol. 1996, 32, 493–501. [Google Scholar] [CrossRef]
- Kennedy, J. Behavioural post-inhibitory rebound in aphids taking flight after exposure to wind. Anim. Behav. 1990, 39, 1078–1088. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 2003, 6, 1025–1037. [Google Scholar] [CrossRef]
- Gish, M.; Inbar, M. Host location by apterous aphids after escape dropping from the plant. J. Insect Behav. 2006, 19, 143. [Google Scholar] [CrossRef]
- Zhang, W.; Chang, X.Q.; Hoffmann, A.A.; Zhang, S.; Ma, C.S. Impact of hot events at different developmental stages of a moth: The closer to adult stage, the less reproductive output. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Webster, B.; Bruce, T.; Pickett, J.; Hardie, J. Volatiles functioning as host cues in a blend become nonhost cues when presented alone to the black bean aphid. Anim. Behav. 2010, 79, 451–457. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Model | DF | χ2 | p |
|---|---|---|---|---|
| Control vs aCO2 | Life stages | 2 | 60.03 | <0.0001 |
| Choices | 1 | 124.81 | <0.0001 | |
| Life stages * Choices | 2 | 22.83 | <0.0001 | |
| Control vs eCO2 | Life stages | 2 | 48.43 | <0.0001 |
| Choices | 1 | 2.62 | 0.1063 | |
| Life stages * Choices | 2 | 0.79 | 0.4557 | |
| eCO2 vs aCO2 | Life stages | 2 | 50.60 | <0.0001 |
| Choices | 1 | 130.45 | <0.0001 | |
| Life stages * Choices | 2 | 13.37 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Martin, C.; Fingu Mabola, J.C.; Verheggen, F.; Wang, Z.; He, K.; Francis, F. Effects of Host Plants Reared under Elevated CO2 Concentrations on the Foraging Behavior of Different Stages of Corn Leaf Aphids Rhopalosiphum maidis. Insects 2019, 10, 182. https://doi.org/10.3390/insects10060182
Chen Y, Martin C, Fingu Mabola JC, Verheggen F, Wang Z, He K, Francis F. Effects of Host Plants Reared under Elevated CO2 Concentrations on the Foraging Behavior of Different Stages of Corn Leaf Aphids Rhopalosiphum maidis. Insects. 2019; 10(6):182. https://doi.org/10.3390/insects10060182
Chicago/Turabian StyleChen, Yu, Clément Martin, Junior Corneille Fingu Mabola, François Verheggen, Zhenying Wang, KangLai He, and Frederic Francis. 2019. "Effects of Host Plants Reared under Elevated CO2 Concentrations on the Foraging Behavior of Different Stages of Corn Leaf Aphids Rhopalosiphum maidis" Insects 10, no. 6: 182. https://doi.org/10.3390/insects10060182
APA StyleChen, Y., Martin, C., Fingu Mabola, J. C., Verheggen, F., Wang, Z., He, K., & Francis, F. (2019). Effects of Host Plants Reared under Elevated CO2 Concentrations on the Foraging Behavior of Different Stages of Corn Leaf Aphids Rhopalosiphum maidis. Insects, 10(6), 182. https://doi.org/10.3390/insects10060182