Hygiene Defense Behaviors Used by a Fungus-Growing Ant Depend on the Fungal Pathogen Stages

 , and
, and 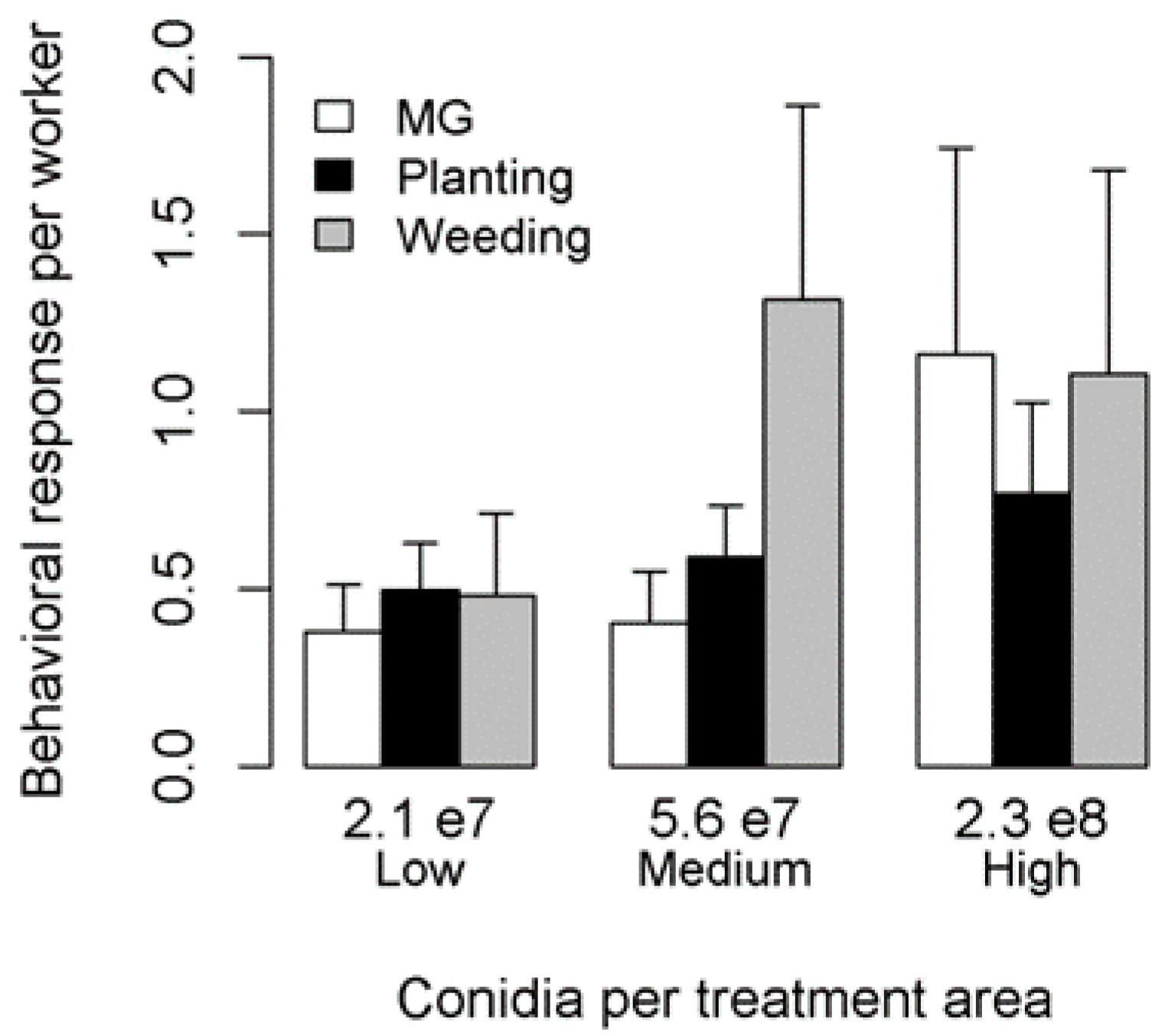{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Groups and Study Behaviors
2.2. Hygienic Responses to Different Metarhizium brunneum Pathogen Concentrations
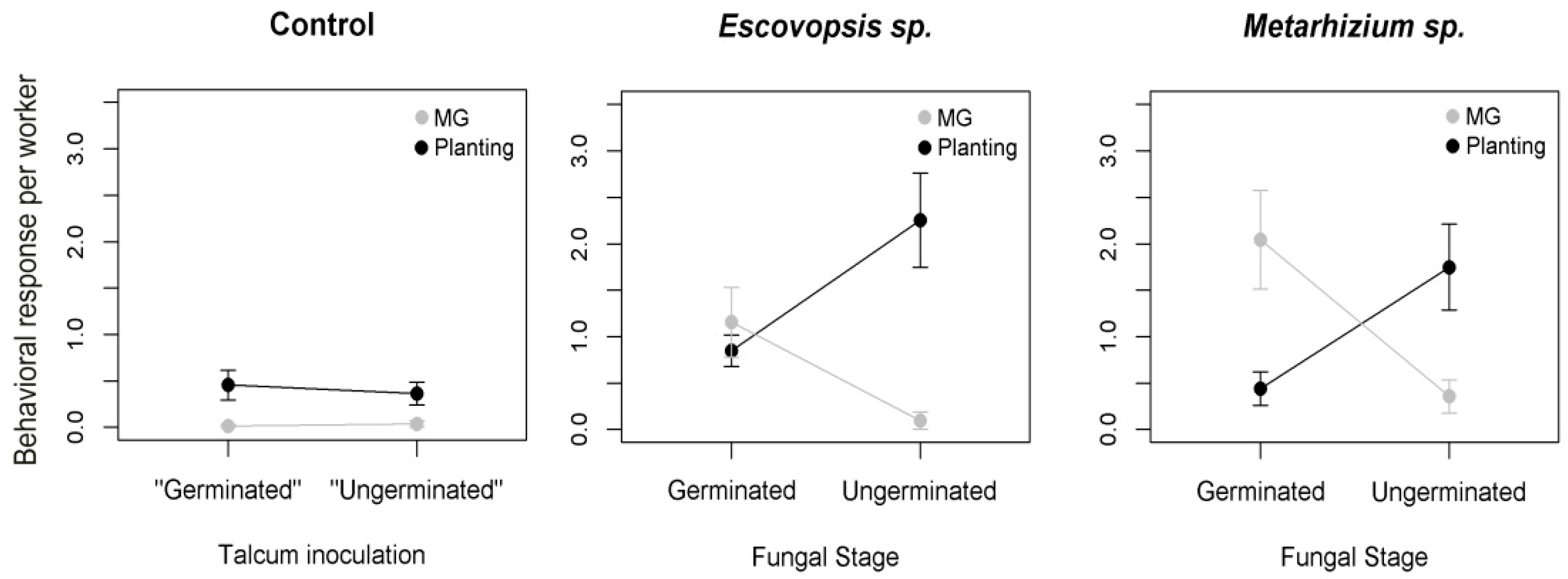
2.3. Hygienic Responses to Different Stages of the Pathogen
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hamilton, W.D. Kinship, recognition, disease, and intelligence: Constraints of social evolution. In Animal Societies: Theories and Facts; Itô, Y., Brown, J.L., Kikkawa, J., Eds.; Japan Science Society Press: Tokyo, Japan, 1987; pp. 81–102. [Google Scholar]
- Godfrey, S.S.; Murray, K.; Bull, C.M.; Gardner, M.G. Transmission mode and distribution of parasites among groups of the social lizard Egernia stokesii. Parasitol. Res. 2006, 99, 223–230. [Google Scholar] [CrossRef]
- Bot, A.; Currie, C.; Hart, A.; Boomsma, J. Waste management in leaf-cutting ants. Ethol. Ecol. Evol. 2001, 13, 225–237. [Google Scholar]
- Diez, L.; Lejeune, P.; Detrain, C. Keep the nest clean: Survival advantages of corpse removal in ants. Boil. Lett. 2014, 10, 20140306. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, A.; Shimizu, S. Resistance of the termite, Coptotermes formosanus Shiraki to Metarhizium anisopliae due to grooming. BioControl 2007, 52, 75–85. [Google Scholar] [CrossRef]
- Bos, N.; Sundström, L.; Fuchs, S.; Freitak, D. Ants medicate to fight disease. Evolution 2015, 69, 2979–2984. [Google Scholar] [CrossRef] [PubMed]
- Castella, G.; Chapuisat, M.; Christe, P. Prophylaxis with resin in wood ants. Anim. Behav. 2008, 75, 1591–1596. [Google Scholar] [CrossRef]
- Bot, A.N.M.; Ortius-Lechner, D.; Finster, K.; Maile, R.; Boomsma, J. Variable sensitivity of fungi and bacteria to compounds produced by the metapleural glands of leaf-cutting ants. Insectes Sociaux 2002, 49, 363–370. [Google Scholar] [CrossRef]
- Cremer, S.; Armitage, S.A.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, R693–R702. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.G.; Gerardo, N.M.; Aanen, D.K.; Six, D.L.; Schultz, T.R. The evolution of agriculture in insects. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 563–595. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Eilenberg, J.; Boomsma, J.J. Trade-offs in group living: Transmission and disease resistance in leaf-cutting ants. Proc. R. Soc. B: Boil. Sci. 2002, 269, 1811–1819. [Google Scholar] [CrossRef]
- Fernández-Marín, H.; Zimmerman, J.K.; Rehner, S.; Wcislo, W.T. Active use of the metapleural glands by ants in controlling fungal infection. Proc. R. Soc. B: Boil. Sci. 2006, 273, 1689–1695. [Google Scholar]
- Currie, C.R.; Scott, J.A.; Summerbell, R.C.; Malloch, D. Fungus-growing ants use antibiotic-producing bacteria to control garden parasites. Nature 1999, 398, 701–704. [Google Scholar] [CrossRef]
- Fernández-Marín, H.; Zimmerman, J.K.; Nash, D.R.; Boomsma, J.J.; Wcislo, W.T. Reduced biological control and enhanced chemical pest management in the evolution of fungus farming in ants. Proc. R. Soc. B Boil. Sci. 2009, 276, 2263–2269. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Marín, H.; Bruner, G.; Gomez, E.B.; Nash, D.R.; Boomsma, J.J.; Wcislo, W.T. Dynamic Disease Management in Trachymyrmex Fungus-Growing Ants (Attini: Formicidae). Am. Nat. 2013, 181, 571–582. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutational Multivariate Analysis of Variance (PERMANOVA). In Wiley StatsRef: Statistics Reference Online; Wiley: Hoboken, NJ, USA, 2017; pp. 1–15. [Google Scholar]
- De Lima Mendonça, A.; Da Silva, C.E.; de Mesquita, F.L.T.; da Silva Campos, R.; Do Nascimento, R.R.; de Azevedo Ximenes, E.C.P.; Sant’Ana, A.E.G. Antimicrobial activities of components of the glandular secretions of leaf cutting ants of the genus Atta. Antonie Van Leeuwenhoek 2009, 95, 295–303. [Google Scholar] [CrossRef]
- Fernández-Marín, H.; Nash, D.R.; Higginbotham, S.; Estrada, C.; Van Zweden, J.S.; D’Ettorre, P.; Wcislo, W.T.; Boomsma, J.J. Functional role of phenylacetic acid from metapleural gland secretions in controlling fungal pathogens in evolutionarily derived leaf-cutting ants. Proc. R. Soc. B Boil. Sci. 2015, 282, 20150212. [Google Scholar] [CrossRef]
- Hughes, W.O.H.; Pagliarini, R.; Madsen, H.B.; Dijkstra, M.B.; Boomsma, J.J. Antimicrobial defense Shows an abrupt evolutionary transition in the fungus-growing ants. Evolution 2008, 62, 1252–1257. [Google Scholar] [CrossRef]
- Poulsen, M.; Bot, A.; Nielsen, M.; Boomsma, J. Experimental evidence for the costs and hygienic significance of the antibiotic metapleural gland secretion in leaf-cutting ants. Behav. Ecol. Sociobiol. 2002, 52, 151–157. [Google Scholar] [CrossRef]
- Richard, F.-J.; Poulsen, M.; Drijfhout, F.; Jones, G.; Boomsma, J.J. Specificity in Chemical Profiles of Workers, Brood and Mutualistic Fungi in Atta, Acromyrmex, and Sericomyrmex Fungus-growing Ants. J. Chem. Ecol. 2007, 33, 2281–2292. [Google Scholar] [CrossRef]
- Van Bael, S.A.; Fernández-Marín, H.; Valencia, M.C.; Rojas, E.I.; Wcislo, W.T.; Herre, E.A. Two fungal symbioses collide: Endophytic fungi are not welcome in leaf-cutting ant gardens. Proc. R. Soc. Lond. B 2009, 276, 2419–2426. [Google Scholar] [CrossRef] [PubMed]
- Gerardo, N.M.; Jacobs, S.R.; Currie, C.R.; Mueller, U.G. Ancient Host–Pathogen Associations Maintained by Specificity of Chemotaxis and Antibiosis. PLoS Boil. 2006, 4, e235. [Google Scholar] [CrossRef] [PubMed]
- Armitage, S.A.O.; Fernández-Marín, H.; Wcislo, W.T.; Boomsma, J.J.; Fernández-Marín, H. An evaluation of the possible adaptive function of fungal brood covering by attine ants. Evolution 2012, 66, 1966–1975. [Google Scholar] [CrossRef] [PubMed]
- Armitage, S.A.O.; Fernández-Marín, H.; Boomsma, J.J.; Wcislo, W.T.; Fernández-Marín, H.; Fernández-Marín, H. Slowing them down will make them lose: A role for attine ant crop fungus in defending pupae against infections? J. Anim. Ecol. 2016, 85, 1210–1221. [Google Scholar] [CrossRef]
- Braga, G.U.; Destéfano, R.H.; Messias, C.L. Oxygen Consumption by Metarhizium anisopliae during Germination and Growth on Different Carbon Sources. J. Invertebr. Pathol. 1999, 74, 112–119. [Google Scholar] [CrossRef]
- Axelrod, R.; Axelrod, D.E.; Pienta, K.J. Evolution of cooperation among tumor cells. Proc. Natl. Acad. Sci. USA 2006, 103, 13474–13479. [Google Scholar] [CrossRef]
- Davis, H.E.; Meconcelli, S.; Radek, R.; McMahon, D.P. Termites shape their collective behavioural response based on stage of infection. Sci. Rep. 2018, 8, 14433. [Google Scholar] [CrossRef]
- Palumbi, S.R. Humans as the world’s greatest evolutionary force. Science 2001, 293, 1786–1790. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonadies, E.; Wcislo, W.T.; Gálvez, D.; Hughes, W.O.H.; Fernández-Marín, H. Hygiene Defense Behaviors Used by a Fungus-Growing Ant Depend on the Fungal Pathogen Stages. Insects 2019, 10, 130. https://doi.org/10.3390/insects10050130
Bonadies E, Wcislo WT, Gálvez D, Hughes WOH, Fernández-Marín H. Hygiene Defense Behaviors Used by a Fungus-Growing Ant Depend on the Fungal Pathogen Stages. Insects. 2019; 10(5):130. https://doi.org/10.3390/insects10050130
Chicago/Turabian StyleBonadies, Ernesto, William T. Wcislo, Dumas Gálvez, William O.H. Hughes, and Hermógenes Fernández-Marín. 2019. "Hygiene Defense Behaviors Used by a Fungus-Growing Ant Depend on the Fungal Pathogen Stages" Insects 10, no. 5: 130. https://doi.org/10.3390/insects10050130
APA StyleBonadies, E., Wcislo, W. T., Gálvez, D., Hughes, W. O. H., & Fernández-Marín, H. (2019). Hygiene Defense Behaviors Used by a Fungus-Growing Ant Depend on the Fungal Pathogen Stages. Insects, 10(5), 130. https://doi.org/10.3390/insects10050130