First Record of the Velvet Ant Mutilla europaea (Hymenoptera: Mutillidae) Parasitizing the Bumblebee Bombus breviceps (Hymenoptera: Apidae)


Abstract
:1. Introduction
2. Materials and Methods
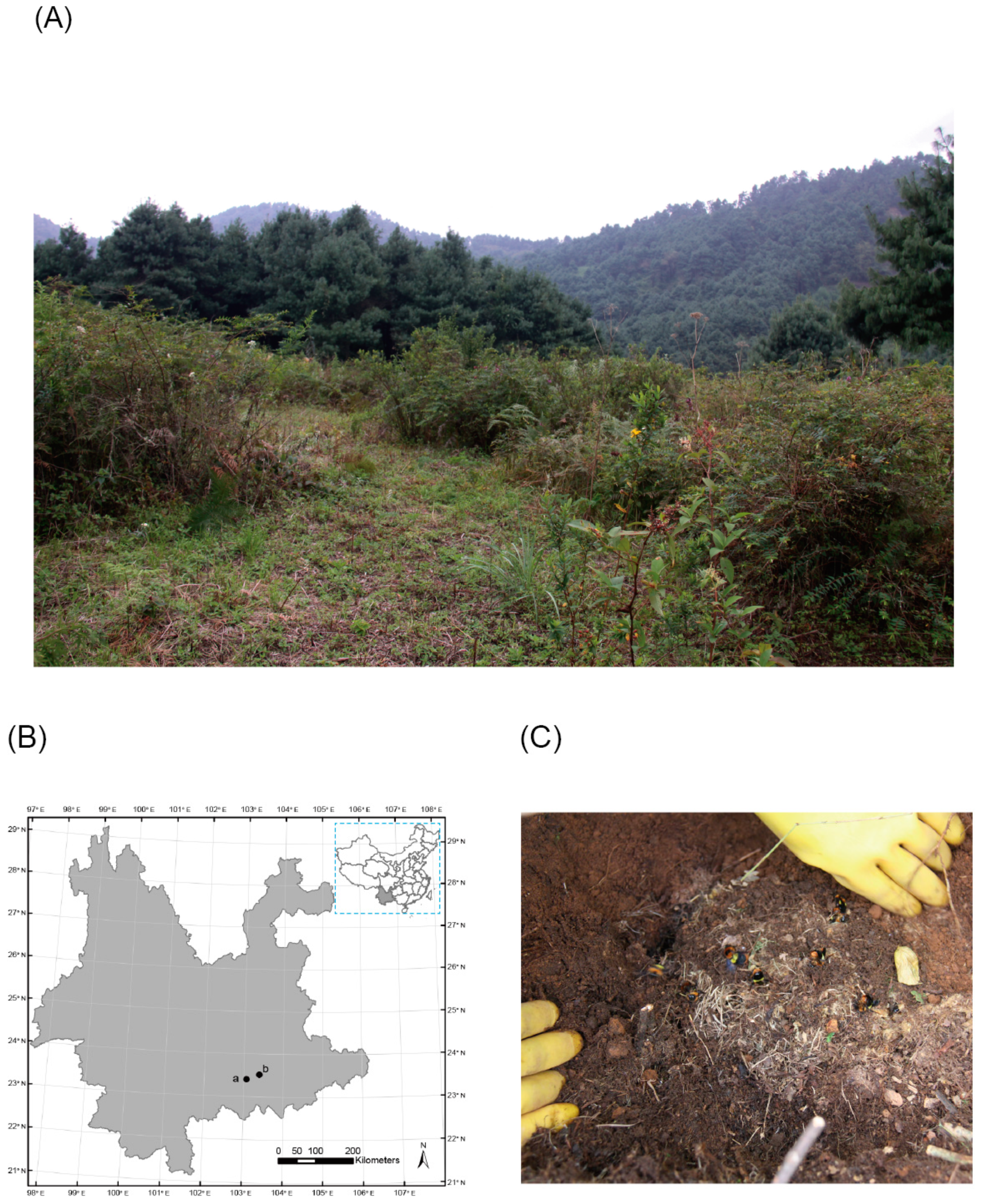
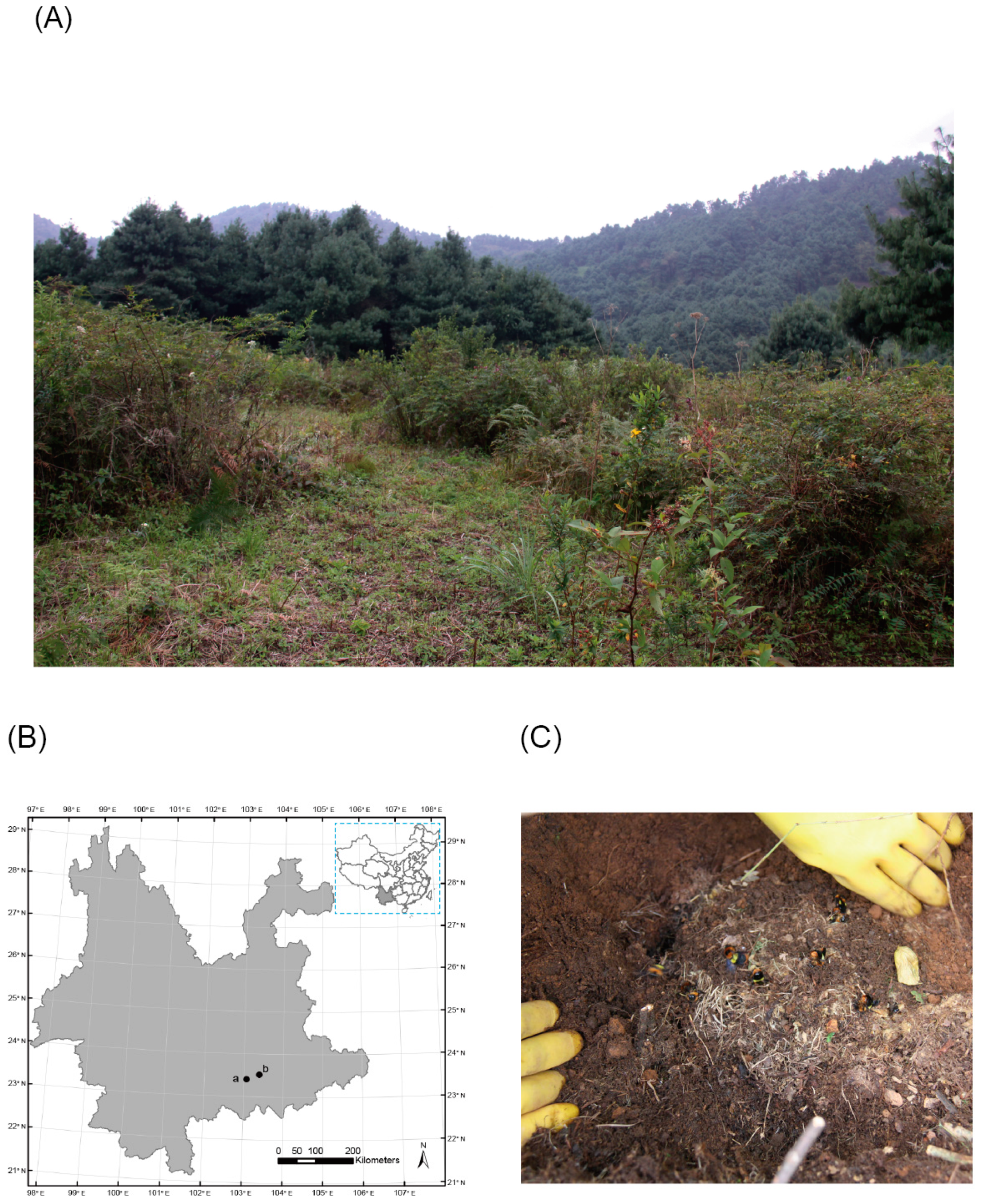
2.1. Samples
2.2. Biology of B. breviceps Parasitism by M. europaea
2.3. Observation of the Dispersal Ability of M. europaea
3. Results
3.1. Field Investigation of the Parasite
3.2. Biology of M. europaea Parasitism in B. breviceps Colonies
3.3. Dispersal Ability
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deyrup, M.; Manley, D. Sex-biased size variation in velvet ants (Hymenoptera: Mutillidae). Fla. Entomol. 1986, 69, 327. [Google Scholar] [CrossRef]
- Brothers, D.J. Alternative life-history styles of mutillid wasps (Insecta Hymenoptera). In Alternative Life-History Styles of Animals. Perspectives in Vertebrate Science; Bruton, M.N., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1989; pp. 279–291. [Google Scholar]
- Brothers, D.J.; Tschuch, G.; Burger, F. Associations of Mutillid wasps (Hymenoptera, Mutillidae) with eusocial insects. Insectes Soc. 2000, 47, 201–211. [Google Scholar] [CrossRef]
- Mickel, C.E. Biological and Taxonomic Investigations on the Mutillid Wasps; United States Government Printing Office: Washington, DC, USA, 1928; 351p.
- Amini, A.; Lelej, A.S.; Sadeghi, H.; Karimi, J. First record of the velvet ants (Hymenoptera: Mutillidae) reared from puparia of the ber fruit fly Carpomya vesuviana Costa (Diptera: Tephritidae) in Iran. Zootaxa 2014, 3861, 585–590. [Google Scholar] [CrossRef]
- Eminson, R.A.F. Observations on Glossina morsitans in Northern Rhodesia. B Entomol. Res. 1915, 5, 381–382. [Google Scholar]
- Péringuey, L. Descriptions of some new or little known South African Mutillidae in the collection of the South African museum. Ann. S. Afr. Mus. 1898, 1, 33–94. [Google Scholar]
- Seyrig, A. Un Mutillide parasite d’un Lépidoptère: Stenomutilla freyi. In Livre Jubilaire de M. Eugène-Louis Bouvier, Member de l’Institut, Professor Honoraire au Muséum; Firmin-Didot: Paris, France, 1936; pp. 313–316. [Google Scholar]
- Mickel, C.E. Mutillidae miscellanea: Taxonomy and distribution. Ann. Entomol. Soc. Am. 1974, 67, 461–471. [Google Scholar] [CrossRef]
- Lelej, A.; Brothers, D. The genus-group names of Mutillidae (Hymenoptera) and their type species, with a new genus, new name, new synonymies, new combinations and lectotypifications. Zootaxa 2008, 1889, 1–79. [Google Scholar]
- Bayliss, P.S.; Brothers, D.J. Biology of Tricholabiodes Radoszkowski in Southern Africa, with a new synonymy and review of recent biological literature (Hymenoptera: Mutillidae). J. Hymenopt. Res. 1996, 5, 249–258. [Google Scholar]
- Fontaine, C.; Dajoz, I.; Meriguet, J.; Loreau, M. Functional diversity of plant-pollinator interaction webs enhances the persistence of plant communities. PLoS Biol. 2006, 4, e1. [Google Scholar] [CrossRef]
- Velthuis, H.; van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Sladen, F.W.L. The Humble-Bee: Its Life-History and How to Domesticate It, with Descriptions of All the British Species of Bombus and Psithyrus; Macmillan: London, UK, 1912; 271p. [Google Scholar]
- Christ, J.L. Naturgeschichte, Klassification und Nomenclatur der Insekten vom Bienen, Wespen und Ameisengeschlecht.als der fünften Klasse fünfte Ordnung des Linnéischen Natursystems von den Insekten: Hymenoptera, mit häutigen Flügeln; Humboldt-Universität zu Berlin: Berlin, Germany, 1791; 535p. [Google Scholar]
- Williams, P.H. An annotated checklist of bumble bees with an analysis of patterns of description (Hymenoptera: Apidae, Bombini). Bull. Br. Mus. Entomol. 1998, 67, 79–152. [Google Scholar]
- Williams, P.H.; Tang, Y.; Yao, J.; Cameron, S. The bumblebees of Sichuan (Hymenoptera: Apidae, Bombini). Syst. Biodivers. 2009, 7, 101–189. [Google Scholar] [CrossRef]
- Huang, J.X.; An, J.D. Species diversity, pollination application and strategy for conservation of the bumblebees of China. Biodivers. Sci. 2018, 26, 486–497. [Google Scholar] [CrossRef]
- Deka, T.N.; Gudade, B.A.; Saju, K.A.; Vijayan, A.K.; Chhetri, P. Foraging behaviour of bumble bee (Bombus breviceps Smith) in large cardamom (Amomum subulatum Roxb.) at Dzongu, North Sikkim. Ecol. Environ. Conserv. 2014, 20, 197–199. [Google Scholar]
- Deka, T.N.; Sudharshan, M.R.; Saju, K.A. New record of bumble bee, Bombus breviceps Smith as a pollinator of large cardamom. Curr. Sci. India 2011, 100, 926–928. [Google Scholar]
- Zhang, H.; Zhou, Z.; Huang, J.; Yuan, X.; Ding, G.; An, J. Queen traits and colony size of four bumblebee species of China. Insectes Soc. 2018, 65, 537–547. [Google Scholar] [CrossRef]
- Osborne, J.L.; Martin, A.P.; Shortall, C.R.; Todd, A.D.; Goulson, D.; Knight, M.E.; Hale, R.J.; Sanderson, R.A. Quantifying and comparing bumblebee nest densities in gardens and countryside habitats. J. Appl. Ecol. 2008, 45, 784–792. [Google Scholar] [CrossRef]
- Geib, J.C.; Strange, J.P.; Galenj, C. Bumble bee nest abundance, foraging distance, and host-plant reproduction: Implications for management and conservation. Ecol. Appl. 2016, 25, 768–778. [Google Scholar] [CrossRef]
- Hoffer, E. Zur biologie der Mutilla europaea L. Zool. Jahrb. 1886, 1, 679–686. [Google Scholar]
- Yang, D.R. The status of species diversity and conservation strategy of bumble bee, a pollination insect in Lancang River Basin of Yunnan. Chin. Biodivers. 1999, 7, 170–174. [Google Scholar]
- Uboni, A.; Lorenzi, M.C. Poor odors, strength, and persistence give their rewards to Mutilla europaea visiting dangerous wasp nests. J. Insect Behav. 2013, 26, 246–252. [Google Scholar] [CrossRef]
- Schmidt, J.O.; Blum, M.S. Adaptions and responses of Dasymutilla occidentalis (Hymenoptera: Mutillidae) to predators. Entomol. Exp. Appl. 2011, 21, 99–111. [Google Scholar] [CrossRef]
- Uboni, A.; Bagnères, A.G.; Christidès, J.P.; Lorenzi, M.C. Cleptoparasites, social parasites and a common host: Chemical insignificance for visiting host nests, chemical mimicry for living in. J. Insect Physiol. 2012, 58, 1259–1264. [Google Scholar] [CrossRef]
- Polidori, C.A.M.P. Temporal asynchrony and spatial co-occurrence with the host: The foraging patterns of Nemka viduata, a parasitoid of digger wasps (Hymenoptera: Mutillidae and Crabronidae). J. Ethol. 2010, 28, 353–361. [Google Scholar] [CrossRef]
- Miyamoto, S. Biology of Bombus ignitus Smith. Jpn. J. Entomol. 1963, 31, 91–98. [Google Scholar]
- Timothy, M.J.; Roxane, M.M.; Matthew, P.F. A nutritional profile of the social wasp Polistes metricus: Differences in nutrient levels between castes and changes within castes during the annual life cycle. J. Insect Physiol. 2010, 56, 42–56. [Google Scholar]
- Evans, D.A.; Miller, B.R. A note on adult overwintering of Dasymutilla nigripes in Michigan (Hymenoptera: Mutillidae). Mich. Entomol. 1969, 2, 74. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Colony ID | Queen | Virgin Queen | Worker | Queen Pupae | Female of M. europaea |
|---|---|---|---|---|---|
| 1 | Yes | 0 | 80 | 15 | 20 |
| 2 | Yes | 0 | 200 | 13 | >10 |
| 3 | Yes | 0 | 150 | 10 | >10 |
| 4 | Yes | 10 | 150 | 20 | >10 |
| 5 | Yes | 0 | 200 | 20 | >10 |
| 6 | Yes | 0 | 30 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, W.; Liang, C.; Ding, G.; Jiang, Y.; Huang, J.; Wu, J. First Record of the Velvet Ant Mutilla europaea (Hymenoptera: Mutillidae) Parasitizing the Bumblebee Bombus breviceps (Hymenoptera: Apidae). Insects 2019, 10, 104. https://doi.org/10.3390/insects10040104
Su W, Liang C, Ding G, Jiang Y, Huang J, Wu J. First Record of the Velvet Ant Mutilla europaea (Hymenoptera: Mutillidae) Parasitizing the Bumblebee Bombus breviceps (Hymenoptera: Apidae). Insects. 2019; 10(4):104. https://doi.org/10.3390/insects10040104
Chicago/Turabian StyleSu, Wenting, Cheng Liang, Guiling Ding, Yusuo Jiang, Jiaxing Huang, and Jie Wu. 2019. "First Record of the Velvet Ant Mutilla europaea (Hymenoptera: Mutillidae) Parasitizing the Bumblebee Bombus breviceps (Hymenoptera: Apidae)" Insects 10, no. 4: 104. https://doi.org/10.3390/insects10040104
APA StyleSu, W., Liang, C., Ding, G., Jiang, Y., Huang, J., & Wu, J. (2019). First Record of the Velvet Ant Mutilla europaea (Hymenoptera: Mutillidae) Parasitizing the Bumblebee Bombus breviceps (Hymenoptera: Apidae). Insects, 10(4), 104. https://doi.org/10.3390/insects10040104